
Partial-, Double-Enzymatic Dephosphorylation and EndoGluC Hydrolysis as an Original Approach to Enhancing Identification of Casein Phosphopeptides (CPPs) by Mass Spectrometry

 , and
, and 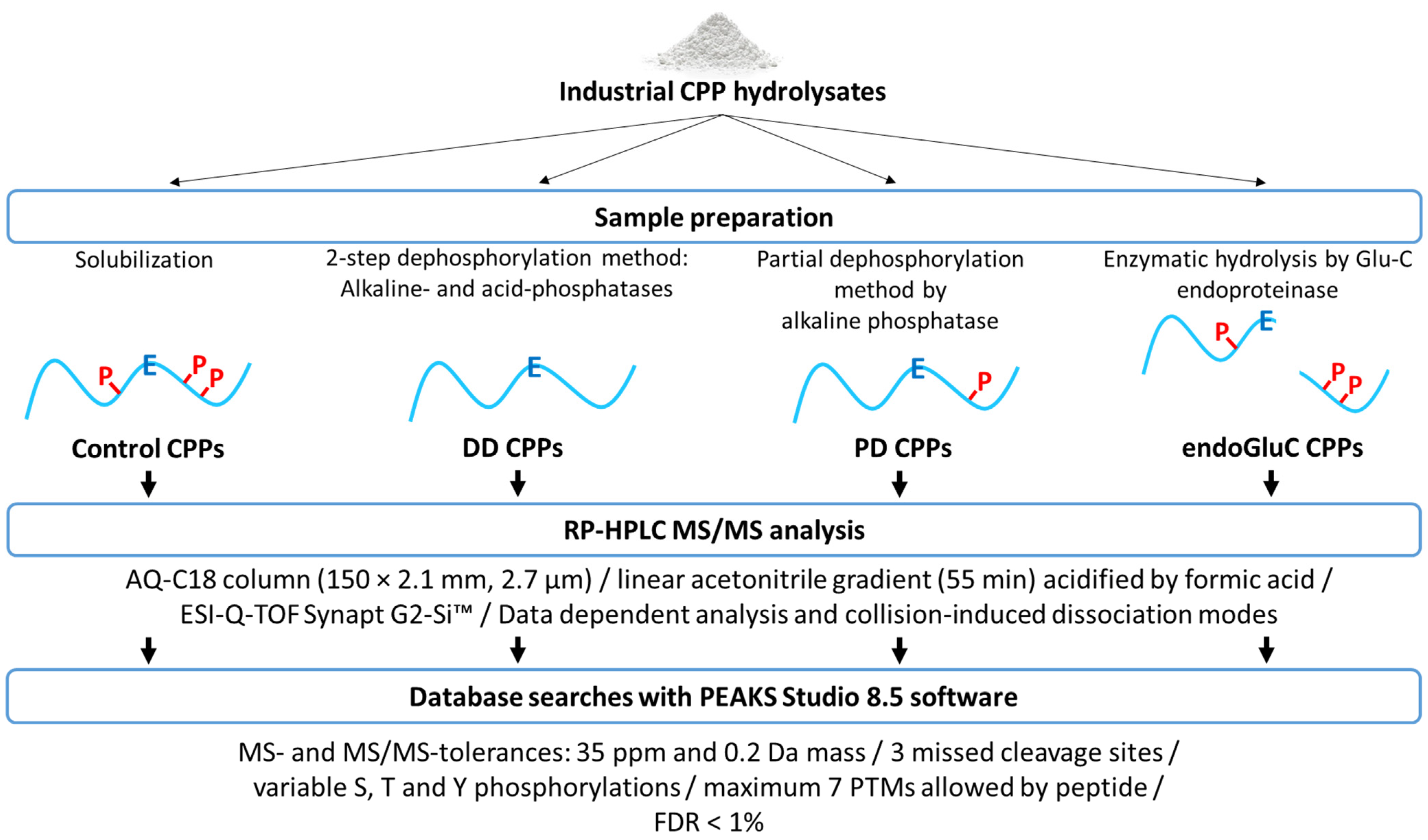{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
2.2. Preparation of Control CPP Hydrolysates
2.3. Preparation of Doubly Dephosphorylated CPP Hydrolysates
2.4. Preparation of Partially Dephosphorylated CPP Hydrolysates
2.5. Preparation of endoGluC Digested CPP Hydrolysates
2.6. CPP Identification by RP-HPLC-MS/MS and Database Search
3. Results and Discussion
3.1. MS Data Comparison of the Four Phases of the Protocol
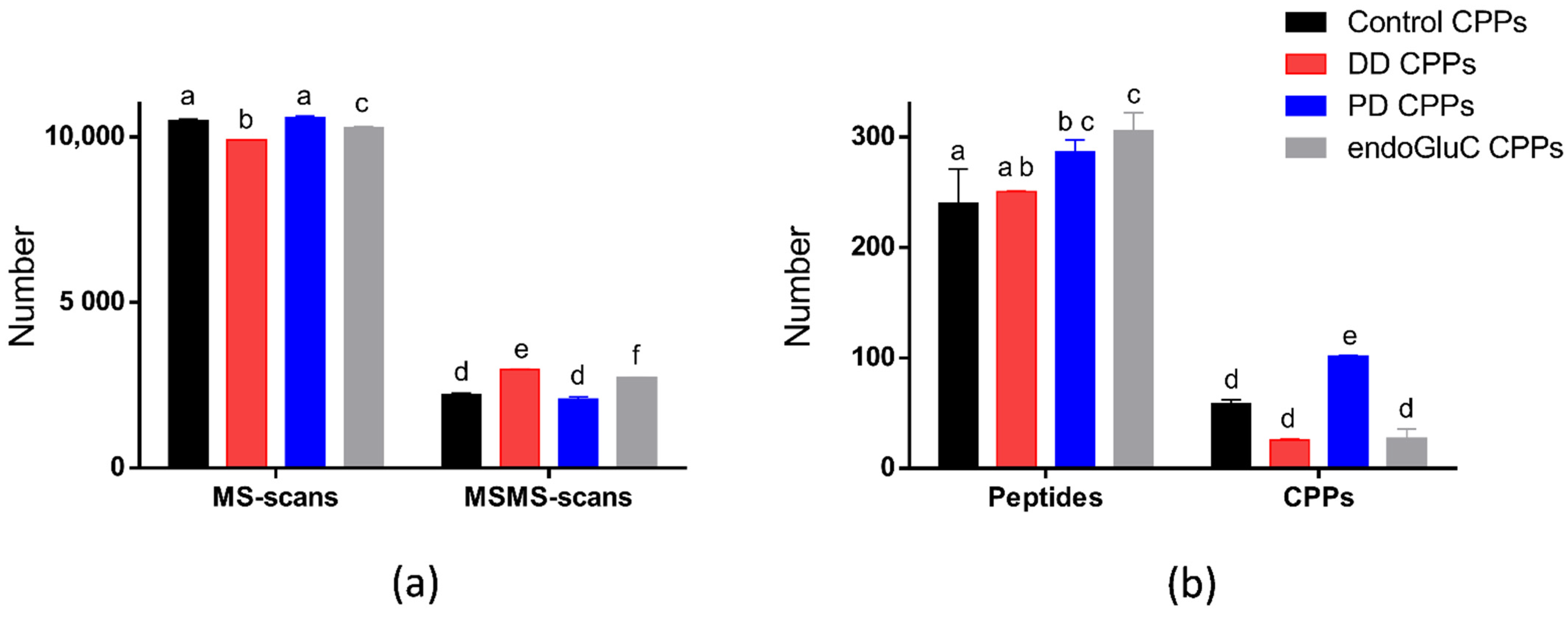
3.1.1. MS Scans, MS/MS Scans and Identified Peptides
3.1.2. CPP Identification
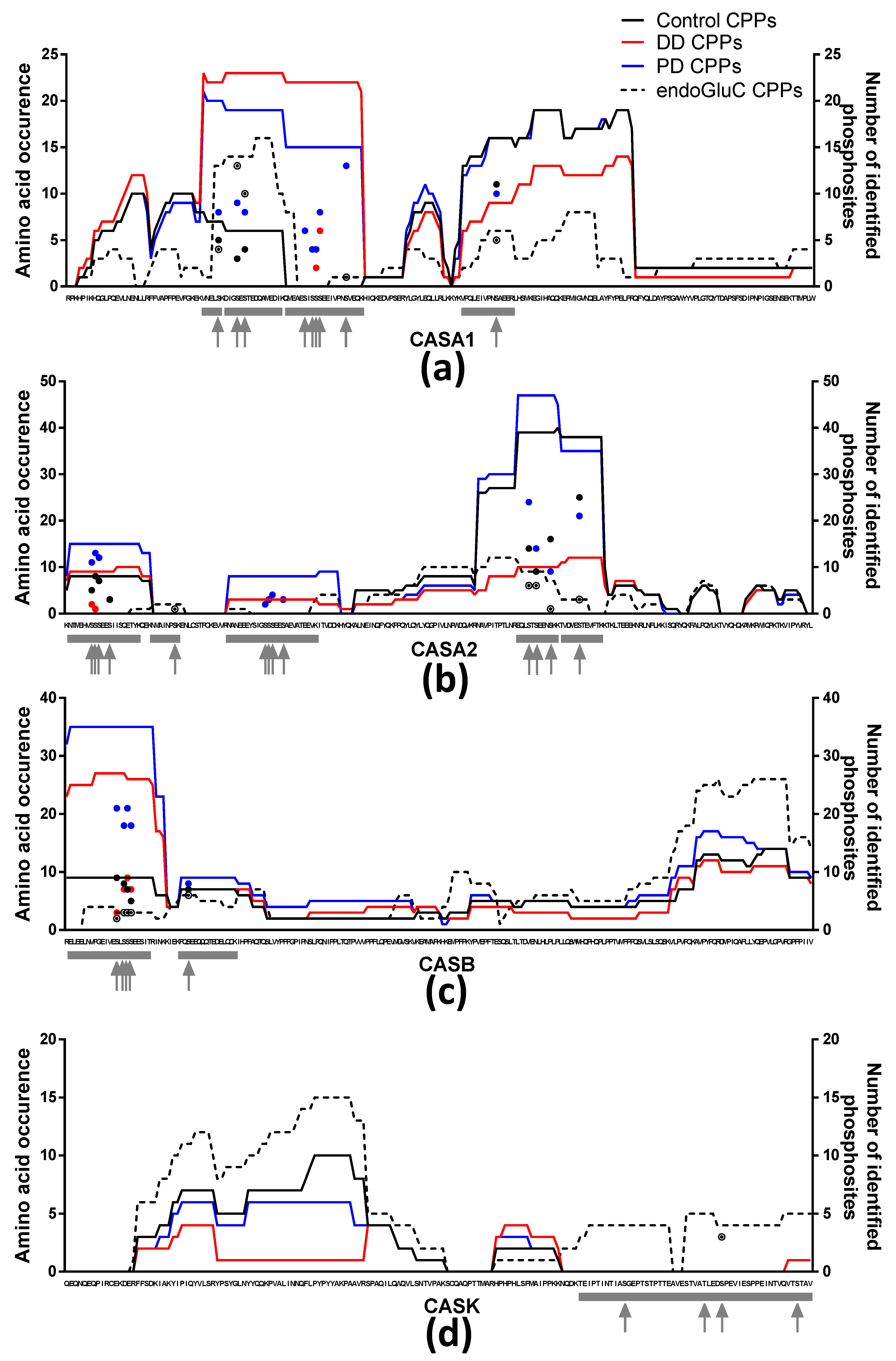
3.2. CPPs and Phosphosites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Petrotos, K.; Tsakali, E.; D’Alessandro, A. Casein and Whey Proteins in Human Health; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 94–146. ISBN 978-1-4443-3683-2. [Google Scholar]
- Ferraretto, A.; Gravaghi, C.; Fiorilli, A.; Tettamanti, G. Casein-derived bioactive phosphopeptides: Role of phosphorylation and primary structure in promoting calcium uptake by HT-29 tumor cells. FEBS Lett. 2003, 551, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Gravaghi, C.; Del Favero, E.; Cantu’, L.; Donetti, E.; Bedoni, M.; Fiorilli, A.; Tettamanti, G.; Ferraretto, A. Casein phosphopeptide promotion of calcium uptake in HT-29 cells—Relationship between biological activity and supramolecular structure. FEBS J. 2007, 274, 4999–5011. [Google Scholar] [CrossRef]
- Cruz-Huerta, E.; García-Nebot, M.J.; Miralles, B.; Recio, I.; Amigo, L. Caseinophosphopeptides released after tryptic hydrolysis versus simulated gastrointestinal digestion of a casein-derived by-product. Food Chem. 2015, 168, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.; Decker, E.A. Antioxidant mechanisms of caseinophosphopeptides and casein hydrolysates and their application in ground beef. J. Agric. Food Chem. 2004, 52, 8208–8213. [Google Scholar] [CrossRef]
- Kitts, D.D.; Nakamura, S. Calcium-enriched casein phosphopeptide stimulates release of IL-6 cytokine in human epithelial intestinal cell line. J. Dairy Res. 2006, 73, 44–48. [Google Scholar] [CrossRef]
- Bottani, M.; Cattaneo, S.; Pica, V.; Stuknyte, M.; Gomarasca, M.; Lombardi, G.; Banfi, G.; de Noni, I.; Ferraretto, A. Gastrointestinal in vitro digests of infant biscuits formulated with bovine milk proteins positively affect in vitro differentiation of human osteoblast-like cells. Foods 2020, 9, 1510. [Google Scholar] [CrossRef] [PubMed]
- García-Nebot, M.J.; Cilla, A.; Alegría, A.; Barberá, R. Caseinophosphopeptides exert partial and site-specific cytoprotection against H2O2-induced oxidative stress in Caco-2 cells. Food Chem. 2011, 129, 1495–1503. [Google Scholar] [CrossRef]
- Aimutis, W.R. Bioactive Properties of Milk Proteins with Particular Focus on Anticariogenesis. J. Nutr. 2004, 134, 989S–995S. [Google Scholar] [CrossRef] [Green Version]
- Kanekanian, A.D.; Williams, R.J.H.; Brownsell, V.L.; Andrews, A.T. Caseinophosphopeptides and dental protection: Concentration and pH studies. Food Chem. 2008, 107, 1015–1021. [Google Scholar] [CrossRef]
- Zhu, Y.S.; FitzGerald, R.J. Direct nanoHPLC-ESI-QTOF MS/MS analysis of tryptic caseinophosphopeptides. Food Chem. 2010, 123, 753–759. [Google Scholar] [CrossRef]
- McDonagh, D.; FitzGerald, R.J. Production of caseinophosphopeptides (CPPs) from sodium caseinate using a range of commercial protease preparations. Int. Dairy J. 1998, 8, 39–45. [Google Scholar] [CrossRef]
- Salimi, K.; Usta, D.D.; Çelikbıçak, Ö.; Pinar, A.; Salih, B.; Tuncel, A. Ti(IV) carrying polydopamine-coated, monodisperse-porous SiO2 microspheres with stable magnetic properties for highly selective enrichment of phosphopeptides. Colloids Surf. B Biointerfaces 2017, 153, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, A.L.; Cavaliere, C.; Ferraris, F.; Gianotti, V.; Laus, M.; Piovesana, S.; Sparnacci, K.; Zenezini Chiozzi, R.; Laganà, A. New Ti-IMAC magnetic polymeric nanoparticles for phosphopeptide enrichment from complex real samples. Talanta 2018, 178, 274–281. [Google Scholar] [CrossRef]
- Zhu, Y.-S.; FitzGerald, R.J. Caseinophosphopeptide enrichment and identification. Int. J. Food Sci. Technol. 2012, 47, 2235–2242. [Google Scholar] [CrossRef]
- Bian, Y.; Song, C.; Cheng, K.; Dong, M.; Wang, F.; Huang, J.; Sun, D.; Wang, L.; Ye, M.; Zou, H. An enzyme assisted RP-RPLC approach for in-depth analysis of human liver phosphoproteome. J. Proteom. 2014, 96, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Beausoleil, S.A.; Jedrychowski, M.; Schwartz, D.; Elias, J.E.; Villén, J.; Li, J.; Cohn, M.A.; Cantley, L.C.; Gygi, S.P. Large-scale characterization of HeLa cell nuclear phosphoproteins. Proc. Natl. Acad. Sci. USA 2004, 101, 12130–12135. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, N.; Masuda, T.; Shinoda, K.; Nakamura, A.; Tomita, M.; Ishihama, Y. Phosphopeptide enrichment by aliphatic hydroxy acid-modified metal oxide chromatography for nano-LC-MS/MS in proteomics applications. Mol. Cell. Proteom. 2007, 6, 1103–1109. [Google Scholar] [CrossRef] [Green Version]
- Arrington, J.V.; Hsu, C.C.; Elder, S.G.; Andy Tao, W. Recent advances in phosphoproteomics and application to neurological diseases. Analyst 2017, 142, 4373–4387. [Google Scholar] [CrossRef]
- Heemskerk, A.A.M.; Busnel, J.M.; Schoenmaker, B.; Derks, R.J.E.; Klychnikov, O.; Hensbergen, P.J.; Deelder, A.M.; Mayboroda, O.A. Ultra-low flow electrospray ionization-mass spectrometry for improved ionization efficiency in phosphoproteomics. Anal. Chem. 2012, 84, 4552–4559. [Google Scholar] [CrossRef]
- Oda, Y.; Nagasu, T.; Chait, B.T. Enrichment analysis of phosphorylated proteins as a tool for probing the phosphoproteome. Nat. Biotechnol. 2001, 19, 379–382. [Google Scholar] [CrossRef]
- Mattila, K.; Siltainsuu, J.; Balaspiri, L.; Ora, M.; Lönnberg, H. Derivatization of phosphopeptides with mercapto- and amino-functionalized conjugate groups by phosphate elimination and subsequent Michael addition. Org. Biomol. Chem. 2005, 3, 3039–3044. [Google Scholar] [CrossRef]
- Matéos, A.; Miclo, L.; Mollé, D.; Dary, A.; Girardet, J.M.; Gaillard, J.L. Equine αS1-casein: Characterization of alternative splicing isoforms and determination of phosphorylation levels. J. Dairy Sci. 2009, 92, 3604–3615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Tsugawa, H.; Cajka, T.; Ma, Y.; Lai, Z.; Mehta, S.S.; Wohlgemuth, G.; Barupal, D.K.; Showalter, M.R.; Arita, M.; et al. Identification of small molecules using accurate mass MS/MS search. Mass Spectrom. Rev. 2018, 37, 513–532. [Google Scholar] [CrossRef]
- Defossez, E.; Bourquin, J.; von Reuss, S.; Rasmann, S.; Glauser, G. Eight key rules for successful data-dependent acquisition in mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2021. [Google Scholar] [CrossRef]
- Matéos, A.; Girardet, J.M.; Mollé, D.; Corbier, C.; Gaillard, J.L.; Miclo, L. Identification of phosphorylation sites of equine β-casein isoforms. Rapid Commun. Mass Spectrom. 2010, 24, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- Kyono, Y.; Sugiyama, N.; Tomita, M.; Ishihama, Y. Chemical dephosphorylation for identification of multiply phosphorylated peptides and phosphorylation site determination. Rapid Commun. Mass Spectrom. 2010, 24, 2277–2282. [Google Scholar] [CrossRef] [PubMed]
- Knight, Z.A.; Schilling, B.; Row, R.H.; Kenski, D.M.; Gibson, B.W.; Shokat, K.M. Phosphospecific proteolysis for mapping sites of protein phosphorylation. Nat. Biotechnol. 2003, 21, 1047–1054. [Google Scholar] [CrossRef]
- Fang, Z.H.; Visker, M.H.P.W.; Miranda, G.; Delacroix-Buchet, A.; Bovenhuis, H.; Martin, P. The relationships among bovine αS-casein phosphorylation isoforms suggest different phosphorylation pathways. J. Dairy Sci. 2016, 99, 8168–8177. [Google Scholar] [CrossRef] [PubMed]
- Kappeler, S.; Farah, Z.; Puhan, Z. Sequence analysis of Camelus dromedarius milk caseins. J. Dairy Res. 1998, 65, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Kawasue, S.; Sakaguchi, Y.; Koga, R.; Hayama, T.; Yoshida, H.; Nohta, H. Multiple phosphorylated protein selective analysis via fluorous derivatization and liquid chromatography-electrospray ionization-mass spectrometry analysis. Anal. Biochem. 2021, 628, 114247. [Google Scholar] [CrossRef] [PubMed]
- Panyayai, T.; Ngamphiw, C.; Tongsima, S.; Mhuantong, W.; Limsripraphan, W.; Choowongkomon, K.; Sawatdichaikul, O. FeptideDB: A web application for new bioactive peptides from food protein. Heliyon 2019, 5, e02076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deracinois, B.; Matéos, A.; Romelard, A.; Boulier, A.; Auger, J.; Baniel, A.; Ravallec, R.; Flahaut, C. Partial-, Double-Enzymatic Dephosphorylation and EndoGluC Hydrolysis as an Original Approach to Enhancing Identification of Casein Phosphopeptides (CPPs) by Mass Spectrometry. Foods 2021, 10, 2134. https://doi.org/10.3390/foods10092134
Deracinois B, Matéos A, Romelard A, Boulier A, Auger J, Baniel A, Ravallec R, Flahaut C. Partial-, Double-Enzymatic Dephosphorylation and EndoGluC Hydrolysis as an Original Approach to Enhancing Identification of Casein Phosphopeptides (CPPs) by Mass Spectrometry. Foods. 2021; 10(9):2134. https://doi.org/10.3390/foods10092134
Chicago/Turabian StyleDeracinois, Barbara, Aurélie Matéos, Audrey Romelard, Audrey Boulier, Julie Auger, Alain Baniel, Rozenn Ravallec, and Christophe Flahaut. 2021. "Partial-, Double-Enzymatic Dephosphorylation and EndoGluC Hydrolysis as an Original Approach to Enhancing Identification of Casein Phosphopeptides (CPPs) by Mass Spectrometry" Foods 10, no. 9: 2134. https://doi.org/10.3390/foods10092134