
Tropical Fruits and Their Co-Products as Bioactive Compounds and Their Health Effects: A Review



Abstract
:1. Introduction
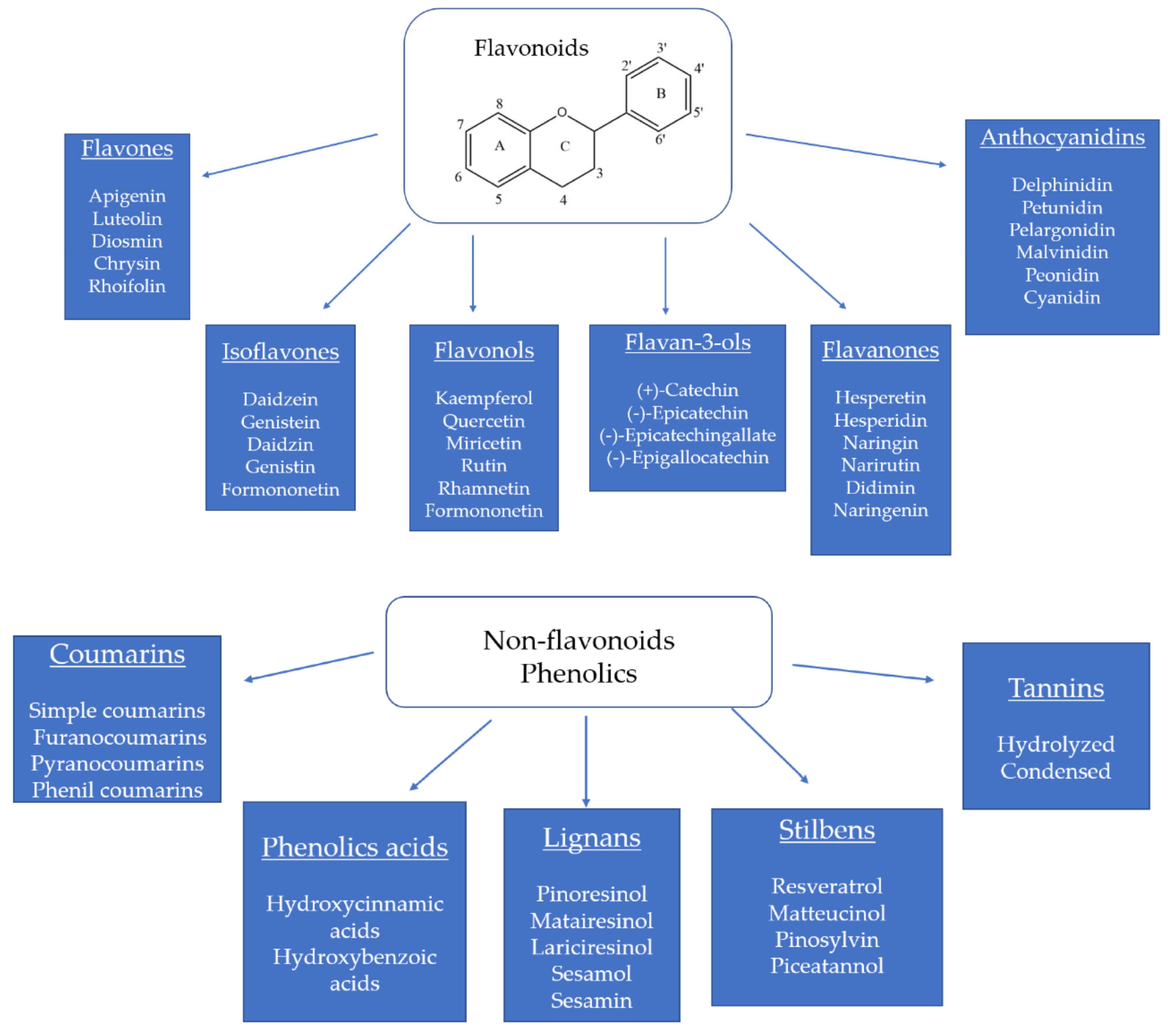
2. Flavonoids and Non-Flavonoid Phenolics in Tropical Fruits
2.1. Açai
2.2. Pomegranate
2.3. Passion Fruit
2.4. Guava
2.5. Pineapple
2.6. Mango
2.7. Avocado
2.8. Tamarind
2.9. Coconut
2.10. Cocoa
2.11. Banana
2.12. Jackfruit
2.13. Other Fruits
3. Biological Effects
3.1. Açai
3.2. Pomegranate
3.3. Passion Fruit
3.4. Guava
3.5. Pineapple
3.6. Mango
3.7. Avocado
3.8. Tamarind
3.9. Coconut
3.10. Banana
3.11. Cocoa
3.12. Jackfruit
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villacís-Chiriboga, J.; Elst, K.; Van Camp, J.; Vera, E.; Ruales, J. Valorization of byproducts from tropical fruits: Extraction methodologies, applications, environmental, and economic assessment: A review (Part 1: General overview of the byproducts, traditional biorefinery practices, and possible applications). Compr. Rev. Food Sci. Food Saf. 2020, 19, 405–447. [Google Scholar] [CrossRef] [PubMed]
- Ding, P. Tropical fruits. In Encyclopedia of Applied Plant Sciences; Thomas, B., Murray, B.G., Murphy, D.J., Eds.; Academic Press: Oxford, UK, 2017; pp. 431–434. [Google Scholar]
- Cheok, C.Y.; Mohd Adzahan, N.; Abdul Rahman, R.; Zainal Abedin, N.H.; Hussain, N.; Sulaiman, R.; Chong, G.H. Current trends of tropical fruit waste utilization. Crit. Rev. Food Sci. Nutr. 2018, 58, 335–361. [Google Scholar] [CrossRef] [PubMed]
- Resende, L.M.; Franca, A.S. Flours based on exotic fruits and their processing residues-features and potential applications to health and disease prevention. In Flour and Breads and Their Fortification in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2019; pp. 387–401. ISBN 9780128146392. [Google Scholar]
- Altendorf, S. Major Tropical Fruits Market Review 2018; FAO: Rome, Italy, 2019; pp. 1–9. [Google Scholar]
- De la Luz Cádiz-Gurrea, M.; del Carmen Villegas-Aguilar, M.; Leyva-Jiménez, F.J.; Pimentel-Moral, S.; Fernández-Ochoa, Á.; Alañón, M.E.; Segura-Carretero, A. Revalorization of bioactive compounds from tropical fruit by-products and industrial applications by means of sustainable approaches. Food Res. Int. 2020, 138, 109786. [Google Scholar] [CrossRef] [PubMed]
- Arraes Maia, G.; Ribeiro da Silva, L.M.; Matias do Prado, G.; Vasconcelos Fonseca, A.V.; Machado de Sousa, P.H.; de Figueiredo, R.W. Development of Mixed Beverages Based on Tropical Fruits. In Non-Alcoholic Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 129–162. [Google Scholar] [CrossRef]
- Alvarez-Rivera, G.; Ballesteros-Vivas, D.; Ibañez, E.; Parada-Alfonso, F.; Cifuentes, A. Foodomics of Bioactive Compounds from Tropical Fruits By-Products. In Comprehensive Foodomics; Cifuentes, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 672–688. [Google Scholar] [CrossRef]
- Schulz, M.; Seraglio, S.K.T.; Brugnerotto, P.; Gonzaga, L.V.; Costa, A.C.O.; Fett, R. Composition and potential health effects of dark-colored underutilized Brazilian fruits—A review. Food Res. Int. 2020, 137, 109744. [Google Scholar] [CrossRef]
- Can-Cauich, C.A.; Sauri-Duch, E.; Betancur-Ancona, D.; Chel-Guerrero, L.; González-Aguilar, G.A.; Cuevas-Glory, L.F.; Pérez-Pacheco, E.; Moo-Huchin, V.M. Tropical fruit peel powders as functional ingredients: Evaluation of their bioactive compounds and antioxidant activity. J. Funct. Foods 2017, 37, 501–506. [Google Scholar] [CrossRef]
- Pereira-Netto, A.B. Tropical fruits as natural, exceptionally rich, sources of bioactive compounds. Int. J. Fruit Sci. 2018, 18, 231–242. [Google Scholar] [CrossRef]
- Azizan, A.; Lee, A.X.; Hamid, N.A.A.; Maulidiani, M.; Mediani, A.; Ghafar, S.Z.A.; Zolkeflee, N.K.Z.; Abas, F. Potentially bioactive metabolites from pineapple waste extracts and their antioxidant and α-glucosidase inhibitory activities by 1H NMR. Foods 2020, 9, 173. [Google Scholar] [CrossRef] [Green Version]
- Pirozzi, A.V.A.; Imbimbo, P.; D’agostino, A.; Tirino, V.; Finamore, R.; Monti, D.M.; Piccoli, R.; Schiraldi, C. Antioxidant and hypolipidemic activity of açai fruit makes it a valuable functional food. Antioxidants 2021, 10, 40. [Google Scholar] [CrossRef]
- El-Hadary, A.E.; Ramadan, M.F. Phenolic profiles, antihyperglycemic, antihyperlipidemic, and antioxidant properties of pomegranate (Punica granatum) peel extract. J. Food Biochem. 2019, 43, 1–9. [Google Scholar] [CrossRef]
- Gómez-García, A.C.; Peñaloza, J.; Cañas, J.; Rojano, B.; Ramos, A.R.; Maldonado, M.E. Impact of Colombian yellow fruits and tropical fruits drinks consumption on the antioxidant status of healthy women. J. Med. Plant. Res. 2019, 10, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Arbizu-Berrocal, S.H.; Kim, H.; Fang, C.; Krenek, K.A.; Talcott, S.T.; Mertens-Talcott, S.U. Polyphenols from mango (Mangifera indica L.) modulate PI3K/AKT/mTOR-associated micro-RNAs and reduce inflammation in non-cancer and induce cell death in breast cancer cells. J. Funct. Foods 2019, 55, 9–16. [Google Scholar] [CrossRef]
- Dabas, D.; Elias, R.J.; Ziegler, G.R.; Lambert, J.D. In vitro antioxidant and cancer inhibitory activity of a colored avocado seed extract. Int. J. Food Sci. 2019, 2019, 6509421. [Google Scholar] [CrossRef] [Green Version]
- Horie, K.; Hossain, M.S.; Morita, S.; Kim, Y.; Yamatsu, A.; Watanabe, Y.; Ohgitani, E.; Mazda, O.; Kim, M. The potency of a novel fermented unripe banana powder as a functional immunostimulatory food ingredient. J. Funct. Foods 2020, 70, 103980. [Google Scholar] [CrossRef]
- De Albuquerque, M.A.C.; Levit, R.; Beres, C.; Bedani, R.; de Moreno de LeBlanc, A.; Saad, S.M.I.; LeBlanc, J.G. Tropical fruit by-products water extracts of tropical fruit by-products as sources of soluble fibres and phenolic compounds with potential antioxidant, anti-inflammatory, and functional properties. J. Funct. Foods 2019, 52, 724–733. [Google Scholar] [CrossRef]
- Singh, P.G.; Madhu, S.B.; Shailasree, S.; TS, G.; Basalingappa, K.M.; BV, S. In Vitro Antioxidant, Anti-Inflammatory and Anti-Microbial Activity of Carica Papaya Seeds. Glob. J. Med. Res. 2020, 20, 19–38. [Google Scholar] [CrossRef]
- Rodríguez-González, S.; Gutiérrez-Ruíz, I.M.; Pérez-Ramírez, I.F.; Mora, O.; Ramos-Gomez, M.; Reynoso-Camacho, R. Mechanisms related to the anti-diabetic properties of mango (Mangifera indica L.) juice by-product. J. Funct. Foods 2017, 37, 190–199. [Google Scholar] [CrossRef]
- Lasano, N.F.; Hamid, A.H.; Karim, R.; Dek, M.S.P.; Shukri, R.; Ramli, N.S. Nutritional Composition, Anti-Diabetic Properties and Identification of Active Compounds Using UHPLC-ESI-Orbitrap-MS/MS in Mangifera odorata L. Peel and seed kernel. Molecules 2019, 24, 320. [Google Scholar] [CrossRef] [Green Version]
- Fidelis, M.; do Carmo, M.A.V.; da Cruz, T.M.; Azevedo, L.; Myoda, T.; Miranda Furtado, M.; Marques, M.B.; Sant’Ana, A.S.; Genovese, M.I.; Oh, W.Y.; et al. Camu-camu seed (Myrciaria dubia)—From side stream to an antioxidant, antihyperglycemic, antiproliferative, antimicrobial, antihemolytic, anti-inflammatory, and antihypertensive ingredient. Food Chem. 2020, 310, 125909. [Google Scholar] [CrossRef]
- Tanghe, A.; Heyman, E.; Vanden Wyngaert, K.; Van Ginckel, A.; Celie, B.; Rietzschel, E.; Calders, P.; Shadid, S. Evaluation of blood pressure lowering effects of cocoa flavanols in diabetes mellitus: A systematic review and meta-analysis. J. Funct. Foods 2021, 79, 104399. [Google Scholar] [CrossRef]
- Xiong, J.; Matta, F.V.; Grace, M.; Lila, M.A.; Ward, N.I.; Felipe-Sotelo, M.; Esposito, D. Phenolic content, anti-inflammatory properties, and dermal wound repair properties of industrially processed and non-processed acai from the Brazilian Amazon. Food Funct. 2020, 11, 4903–4914. [Google Scholar] [CrossRef]
- Norhaslinda, R.; Noratiqah, J.M.; Amin, B.A.; Khalili, R.M.A. Quantitative and optimization of anthocyanin extracted from pomegranate (Punica granatum) extract by High-Performance Liquid Chromatography (HPLC). Pharmacogn. J. 2018, 10, 650–653. [Google Scholar] [CrossRef] [Green Version]
- Viganó, J.; Brumer, I.Z.; de Campos Braga, P.A.; da Silva, J.K.; Maróstica Júnior, M.R.; Reyes Reyes, F.G.; Martínez, J. Pressurized liquids extraction as an alternative process to readily obtain bioactive compounds from passion fruit rinds. Food Bioprod. Process. 2016, 100, 382–390. [Google Scholar] [CrossRef]
- Nunes, J.C.; Lago, M.G.; Castelo-Branco, V.N.; Oliveira, F.R.; Guedes Torres, A.; Perrone, D.; Monteiro, M. Effect of drying method on volatile compounds, phenolic profile and antioxidant capacity of guava powders. Food Chem. 2016, 197, 881–890. [Google Scholar] [CrossRef]
- Sun, G.M.; Zhang, X.M.; Soler, A.; Marie-Alphonsine, P.A. Chapter 25—Nutritional Composition of Pineapple (Ananas comosus (L.) Merr.). In Nutritional Composition of Fruit Cultivars; Simmonds, M.S.J., Preedy, V.R., Eds.; Academic Press: Oxford, UK, 2016; pp. 609–637. [Google Scholar] [CrossRef]
- Marcillo-Parra, V.; Anaguano, M.; Molina, M.; Tupuna-Yerovi, D.S.; Ruales, J. Characterization and quantification of bioactive compounds and antioxidant activity in three different varieties of mango (Mangifera indica L.) peel from the Ecuadorian region using HPLC-UV/VIS and UPLC-PDA. NFS J. 2021, 23, 1–7. [Google Scholar] [CrossRef]
- Trujillo-Mayol, I.; Badillo-Muñoz, G.; Céspedes-Acuña, C.; Alarcón-Enos, J. The relationship between fruit size and phenolic and enzymatic composition of avocado byproducts (Persea americana mill.): The importance for biorefinery applications. Horticulturae 2020, 6, 91. [Google Scholar] [CrossRef]
- Bressiani, P.A.; De Lima, G.R.F.; Düsman, E.; Tonin, L.T.D. Cytotoxic and antioxidant activities of Tamarindus indica pulp extract from Brazil. J. Food Meas. Charact. 2021, 15, 2743–2749. [Google Scholar] [CrossRef]
- Arivalagan, M.; Roy, T.K.; Yasmeen, A.M.; Pavithra, K.C.; Jwala, P.N.; Shivasankara, K.S.; Manikantan, M.R.; Hebbar, K.B.; Kanade, S.R. Extraction of phenolic compounds with antioxidant potential from coconut (Cocos nucifera L.) testa and identification of phenolic acids and flavonoids using UPLC coupled with TQD-MS/MS. LWT Food Sci. Technol. 2018, 92, 116–126. [Google Scholar] [CrossRef]
- Botella-Martínez, C.; Lucas-Gonzalez, R.; Ballester-Costa, C.; Pérez-Álvarez, J.Á.; Fernández-López, J.; Delgado-Ospina, J.; Chaves-López, C.; Viuda-Martos, M. Ghanaian cocoa (Theobroma cacao L.) bean shells coproducts: Effect of particle size on chemical composition, bioactive compound content and antioxidant activity. Agronomy 2021, 11, 401. [Google Scholar] [CrossRef]
- Anyasi, T.A.; Jideani, A.I.O.; Mchau, G.R.A. Phenolics and essential mineral profile of organic acid pretreated unripe banana flour. Food Res. Int. 2018, 104, 100–109. [Google Scholar] [CrossRef]
- Wang, X.L.; Di, X.X.; Shen, T.; Wang, S.Q.; Wang, X.N. New phenolic compounds from the leaves of Artocarpus heterophyllus. Chin. Chem. Lett. 2016, 28, 37–40. [Google Scholar] [CrossRef]
- Latos-Brozio, M.; Masek, A. Structure-activity relationships analysis of monomeric and polymeric polyphenols (quercetin, rutin and catechin) obtained by various polymerization methods. Chem. Biodivers. 2019, 16, e1900426. [Google Scholar] [CrossRef] [PubMed]
- Garzón, G.A.; Narváez-Cuenca, C.E.; Vincken, J.P.; Gruppen, H. Polyphenolic composition and antioxidant activity of açai (Euterpe oleracea Mart.) from Colombia. Food Chem. 2017, 217, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Crespo-López, M.E.; Soares, E.S.; de Matos Macchi, B.; Santos-Sacramento, L.; Takeda, P.Y.; Lopes-Araújo, A.; de Oliveira Paraense, R.S.; Souza-Monteiro, J.R.; Augusto-Oliveira, M.; Luz, D.A.; et al. Towards therapeutic alternatives for mercury neurotoxicity in the amazon: Unraveling the pre-clinical effects of the superfruit açaí (Euterpe oleracea, mart.) as juice for human consumption. Nutrients 2019, 11, 2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, P.S.; Massarioli, A.P.; Lazarini, J.G.; Soares, J.C.; Franchin, M.; Rosalen, P.L.; de Alencar, S.M. Simulated gastrointestinal digestion of Brazilian açaí seeds affects the content of flavan-3-ol derivatives, and their antioxidant and anti-inflammatory activities. Heliyon 2020, 6, e05214. [Google Scholar] [CrossRef]
- Haghighian, M.K.; Rafraf, M.; Hemmati, S.; Haghravan, S.; Asghari-Jafarabadi, M. Effects of pomegranate (Punica granatum L.) peel extract supplementation on serum lipid profile and oxidative stress in obese women with knee osteoarthritis: A double blind, randomized, placebo controlled study. Adv. Integr. Med. 2020, 8, 107–113. [Google Scholar] [CrossRef]
- Rongai, D.; Pulcini, P.; Di Lernia, G.; Nota, P.; Preka, P.; Milano, F. Punicalagin content and antifungal activity of different pomegranate (Punica ganatum L.) genotypes. Horticulturae 2019, 5, 52. [Google Scholar] [CrossRef] [Green Version]
- Kostka, T.; Ostberg-Potthoff, J.J.; Briviba, K.; Matsugo, S.; Winterhalter, P.; Esatbeyoglu, T. Pomegranate (Punica granatum L.) extract and its anthocyanin and copigment fractions—free radical scavenging activity and influence on cellular oxidative stress. Foods 2020, 9, 1617. [Google Scholar] [CrossRef]
- Díaz-Mula, H.M.; Tomás-Barberán, F.A.; García-Villalba, R. Pomegranate fruit and juice (cv. Mollar), rich in ellagitannins and anthocyanins, also provide a significant content of a wide range of proanthocyanidins. J. Agric. Food Chem. 2019, 67, 9160–9167. [Google Scholar] [CrossRef]
- De Souza, C.G.; Rodrigues, T.H.S.; de Silva, L.M.A.; Ribeiro, P.R.V.; de Brito, E.S. Sequential extraction of flavonoids and pectin from yellow passion fruit rind using pressurized solvent or ultrasound: Extraction of flavonoids and pectin from passion fruit. J. Sci. Food Agric. 2018, 98, 1362–1368. [Google Scholar] [CrossRef]
- Carmona-Hernandez, J.C.; Taborda-Ocampo, G.; Valdez, J.C.; Bolling, B.W.; González-Correa, C.H. Polyphenol extracts from three colombian passifloras (passion fruits) prevent inflammation-induced barrier dysfunction of Caco-2 cells. Molecules 2019, 24, 4614. [Google Scholar] [CrossRef] [Green Version]
- Dos Reis, L.C.R.; Facco, E.M.P.; Salvador, M.; Hickmann Flôres, S.; de Oliveira Rios, A. Antioxidant potential and physicochemical characterization of yellow, purple and orange passion fruit. J. Food Sci. Technol. 2018, 55, 2679–2691. [Google Scholar] [CrossRef]
- Lopes dos Santos, W.N.; da Silva Sauthier, M.C.; Pinto dos Santos, A.M.; de Andrade Santana, D.; Almeida Azevedo, R.S.; da Cruz Caldas, J. Simultaneous determination of 13 phenolic bioactive compounds in guava (Psidium guajava L.) by HPLC-PAD with evaluation using PCA and Neural Network Analysis (NNA). Microchem. J. 2017, 133, 583–592. [Google Scholar] [CrossRef]
- Da Silva Lima, R.; Salvador Ferreira, S.R.; Vitali, L.; Block, J.M. May the superfruit red guava and its processing waste be a potential ingredient in functional foods? Food Res. Int. 2019, 115, 451–459. [Google Scholar] [CrossRef]
- Tan, S.; Wang, Z.; Xiang, Y.; Deng, T.; Zhao, X.; Shi, S.; Zheng, Q.; Gao, X.; Li, W. The effects of drying methods on chemical profiles and antioxidant activities of two cultivars of Psidium guajava fruits. LWT Food Sci. Technol. 2020, 118, 108723. [Google Scholar] [CrossRef]
- Rosas Dominguez, C.; Domínguez Avila, J.A.; Pareek, S.; Villegas Ochoa, M.A.; Ayala Zavala, J.F.; Yahia, E.; González-Aguilar, G.A. Content of bioactive compounds and their contribution to antioxidant capacity during ripening of pineapple (Ananas comosus L.) cv Esmeralda. J. Appl. Bot. Food Qual. 2018, 91, 61–68. [Google Scholar] [CrossRef]
- Arampath, P.C.; Dekker, M. Bulk storage of mango (Mangifera indica L.) and pineapple (Ananas comosus L.) pulp: Effect of pulping and storage temperature on phytochemicals and antioxidant activity. J. Sci. Food Agric. 2019, 99, 5157–5167. [Google Scholar] [CrossRef]
- Morales, M.; Zapata, S.; Jaimes, T.R.; Rosales, S.; Alzate, A.F.; Maldonado, M.E.; Zamorano, P.; Rojano, B.A. Mangiferin content, carotenoids, tannins and oxygen radical absorbance capacity (ORAC) values of six mango (Mangifera indica) cultivars from the Colombian Caribbean. J. Med. Plants Res. 2017, 11, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.C.X.; Fu, S.F.; Bi, X.F.; Chen, F.; Liao, X.J.; Hu, X.S.; Wu, H.J. Physico-chemical and antioxidant properties of four mango (Mangifera indica L.) cultivars in China. Food Chem. 2013, 138, 396–405. [Google Scholar] [CrossRef]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R. A Review on Ethnopharmacological Applications, pharmacological activities, and bioactive compounds of Mangifera indica (Mango). Evid. Based Complement. Altern. Med. 2017, 2017, 6949835. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.M.P.; Le, T.T.; Vissenaekens, H.; Gonzales, G.B.; Van Camp, J.; Smagghe, G.; Raes, K. In vitro antioxidant activity and phenolic profiles of tropical fruit by-products. Int. J. Food Sci. Technol. 2019, 54, 1169–1178. [Google Scholar] [CrossRef]
- Sáyago-Ayerdi, S.G.; Moreno-Hernández, C.L.; Montalvo-González, E.; García-Magaña, M.L.; Mata-Montes de Oca, M.; Torres, J.L.; Pérez-Jiménez, J. Mexican “Ataulfo” mango (Mangifera indica L) as a source of hydrolyzable tannins. Analysis by MALDI-TOF/TOF MS. Food Res. Int. 2013, 51, 188–194. [Google Scholar] [CrossRef]
- Castro-Vargas, H.I.; Vivas, D.B.; Barbosa, J.O.; Medina, S.J.M.; Gutiérrez, F.A.; Parada-Alfonso, F. Bioactive phenolic compounds from the agroindustrial waste of Colombian mango cultivars ‘sugar mango’ and ‘tommy atkins’—An alternative for their use and valorization. Antioxidants 2019, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Mwaurah, P.W.; Kumar, S.; Kumar, N.; Panghal, A.; Attkan, A.K.; Singh, V.K.; Garg, M.K. Physicochemical characteristics, bioactive compounds and industrial applications of mango kernel and its products: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2421–2446. [Google Scholar] [CrossRef]
- Salazar-López, N.J.; Domínguez-Avila, J.A.; Yahia, E.M.; Belmonte-Herrera, B.H.; Wall-Medrano, A.; Montalvo-González, E.; González-Aguilar, G.A. Avocado fruit and by-products as potential sources of bioactive compounds. Food Res. Int. 2020, 138, 109774. [Google Scholar] [CrossRef]
- Araujo, R.G.; Rodríguez-Jasso, R.M.; Ruíz, H.A.; Govea-Salas, M.; Pintado, M.; Aguilar, C.N. Recovery of bioactive components from avocado peels using microwave-assisted extraction. Food Bioprod. Process. 2021, 127, 152–161. [Google Scholar] [CrossRef]
- Ramos-Aguilar, A.L.; Ornelas-Paz, J.; Tapia-Vargas, L.M.; Gardea-Béjar, A.A.; Yahia, E.M.; de Jesus Ornelas-Paz, J.; Ruiz-Cruz, S.; Rios-Velasco, C.; Escalante-Minakata, P. Effect of cultivar on the content of selected phytochemicals in avocado peels. Food Res. Int. 2021, 140, 110024. [Google Scholar] [CrossRef]
- Saavedra, J.; Córdova, A.; Navarro, R.; Díaz-Calderón, P.; Fuentealba, C.; Astudillo-Castro, C.; Toledo, L.; Enrione, J.; Galvez, L. Industrial avocado waste: Functional compounds preservation by convective drying process. J. Food Eng. 2017, 198, 81–90. [Google Scholar] [CrossRef]
- López-Cobo, A.; Gómez-Caravaca, A.M.; Pasini, F.; Caboni, M.F.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC-DAD-ESI-QTOF-MS and HPLC-FLD-MS as valuable tools for the determination of phenolic and other polar compounds in the edible part and by-products of avocado. LWT Food Sci. Technol. 2016, 73, 505–513. [Google Scholar] [CrossRef]
- Tril, U.; Fernández-López, J.; Pérez Álvarez, J.A.; Viuda-Martos, M. Chemical, physicochemical, technological, antibacterial and antioxidant properties of rich-fibre powder extract obtained from tamarind (Tamarindus indica L.). Ind. Crop. Prod. 2014, 55, 155–162. [Google Scholar] [CrossRef]
- Nabeel, K.; Fathima, A.; Khanum, F.; Manjula, S.N.; Mruthunjaya, K.; Vengal Rao, P.; Mehdi, S. Anti-obesity effect of tamarindus indicus seed extract against a high-fat diet-induced obese model in rats. Int. J. Res. Pharm. Sci. 2020, 11, 2083–2089. [Google Scholar] [CrossRef] [Green Version]
- Cvetanović, A.; Uysal, S.; Pavlić, B.; Sinan, K.I.; Llorent-Martínez, E.J.; Zengin, G. Tamarindus indica L. Seed: Optimization of Maceration Extraction Recovery of Tannins. Food Anal. Methods 2020, 13, 579–590. [Google Scholar] [CrossRef]
- Govindappa, M.; Birawat, R.K.A.; Raghavendra, V.B.; Munawer, U.; Ningaraju, S.; Al-Rashed, S.; Chowdappa, S.; Pugazhendhi, A. In vitro therapeutic evaluation of nanoliposome loaded with Xyloglucans polysaccharides from Tamarindus flower extract. Int. J. Biol. Macromol. 2021, 178, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Jericó Santos, T.R.; Santos Vasconcelos, A.G.; Lins de Aquino Santana, L.C.; Gualberto, N.C.; Buarque Feitosa, P.R.; Pires de Siqueira, A.C. Solid-state fermentation as a tool to enhance the polyphenolic compound contents of acidic Tamarindus indica by-products. Biocatal. Agric. Biotechnol. 2020, 30, 101851. [Google Scholar] [CrossRef]
- El-Haddad, A.E.S.; Saadeldeen, A.M.; El-Emam, S.Z. Anti-angiogenic activity of major phenolics in tamarind assessed with molecular docking study on vegf kinase proteins. Pakistan J. Biol. Sci. 2019, 22, 502–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahayothee, B.; Koomyart, I.; Khuwijitjaru, P.; Siriwongwilaichat, P.; Nagle, M.; Müller, J. Phenolic compounds, antioxidant activity, and medium chain fatty acids profiles of coconut water and meat at different maturity stages. Int. J. Food Prop. 2016, 19, 2041–2051. [Google Scholar] [CrossRef]
- Li, N.; Jiang, H.; Yang, J.; Wang, C.; Wu, L.; Hao, Y.; Liu, Y. Characterization of phenolic compounds and anti-acetylcholinase activity of coconut shells. Food Biosci. 2021, 42, 101204. [Google Scholar] [CrossRef]
- Mulyadi, A.F.; Schreiner, M.; Dewi, I.A. Phenolic and volatile compounds, antioxidant activity, and sensory properties of virgin coconut oil: Occurrence and their relationship with quality. AIP Conf. Proc. 2018, 2021, 070020. [Google Scholar] [CrossRef]
- Peláez, P.; Bardón, I.; Camasca, P. Methylxanthine and catechin content of fresh and fermented cocoa beans, dried cocoa beans, and cocoa liquor. Sci. Agropecu. 2016, 7, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Urbańska, B.; Kowalska, H.; Szulc, K.; Ziarno, M.; Pochitskaya, I.; Kowalska, J. Comparison of the effects of douching parameters on the contents of three dominant flavan3-ols, rheological properties and sensory quality in chocolate milk mass based on liquor from unroasted cocoa beans. Molecules 2021, 26, 2502. [Google Scholar] [CrossRef]
- Hernández-Hernández, C.; Morales-Sillero, A.; Fernández-Bolaños, J.; Bermúdez-Oria, A.; Morales, A.A.; Rodríguez-Gutiérrez, G. Cocoa bean husk: Industrial source of antioxidant phenolic extract. J. Sci. Food Agric. 2019, 99, 325–333. [Google Scholar] [CrossRef]
- Oliveira, B.G.; Pimentel, E.F.; Pereira, A.C.H.; Tosato, F.; Pinto, F.E.; Ventura, J.A.; Endringer, D.C.; Romão, W. Phenolic and glycidic profiling of bananas Musa spp. associated with maturation stage and cancer chemoprevention activities. Microchem. J. 2020, 153, 104391. [Google Scholar] [CrossRef]
- Lopes, S.; Borges, C.V.; Sousa Cardoso, S.M.; Almeida Pereira da Rocha, M.F.; Maraschin, M. Banana (Musa spp.) as a source of bioactive compounds for health promotion. Handb. Banan. Prod. Postharvest Sci. Process. Technol. Nutr. 2020, 227–244. [Google Scholar] [CrossRef]
- Li, Z.; Lan, Y.; Miao, J.; Chen, X.; Chen, B.; Liu, G.; Wu, X.; Zhu, X.; Cao, Y. Phytochemicals, antioxidant capacity and cytoprotective effects of jackfruit (Artocarpus heterophyllus Lam.) axis extracts on HepG2 cells. Food Biosci. 2021, 41, 100933. [Google Scholar] [CrossRef]
- Fang, S.C.; Hsu, C.L.; Yen, G.C. Anti-inflammatory effects of phenolic compounds isolated from the fruits of Artocarpus heterophyllus. J. Agric. Food Chem. 2008, 56, 4463–4468. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Oszmiański, J.; Golis, T. Phytochemical compounds and biological effects of Actinidia fruits. J. Funct. Foods 2017, 30, 194–202. [Google Scholar] [CrossRef]
- Guedes, T.J.F.L.; Rajan, M.; Barbosa, P.F.; Silva, E.S.; Machado, T.O.X.; Narain, N. Phytochemical composition and antioxidant potential of different varieties viz. Flor Branca, Costa Rica and Junco of green unripe acerola (Malphigia emarginata D.C.) fruits. Food Sci. Technol. 2020, 12, 1–9. [Google Scholar] [CrossRef]
- Batista, A.C.V.; de Alencar Ribeiro, M.; Alves de Oliveira, K.; de Freitas, P.A.; dos Santos, N.S.; Magalhães, L.A.; Magalhães, S.C.; da Cruz Fonseca, S.G.; de Souza Aquino, J.; de Souza, E.L.; et al. Effects of consumption of acerola, cashew and guava by-products on adiposity and redox homeostasis of adipose tissue in obese rats. Clin. Nutr. ESPEN 2021, 43, 283–289. [Google Scholar] [CrossRef]
- Pimenta Inada, K.O.; Nunes, S.; Martínez-Blázquez, J.A.; Tomás-Barberán, F.A.; Perrone, D.; Monteiro, M. Effect of high hydrostatic pressure and drying methods on phenolic compounds profile of jabuticaba (Myrciaria jaboticaba) peel and seed. Food Chem. 2020, 309, 125794. [Google Scholar] [CrossRef]
- Fidelis, M.; Santos, J.S.; Escher, G.B.; Rocha, R.S.; Cruz, A.G.; Cruz, T.M.; Marques, M.B.; Nunes, J.B.; do Carmo, M.A.V.; de Almeida, L.A.; et al. Polyphenols of jabuticaba [Myrciaria jaboticaba (Vell.) O.Berg] seeds incorporated in a yogurt model exert antioxidant activity and modulate gut microbiota of 1,2-dimethylhydrazine-induced colon cancer in rats. Food Chem. 2021, 334, 127565. [Google Scholar] [CrossRef]
- Mehdizadeh, S.; Lasekan, O.; Muhammad, K.; Baharin, B. Variability in the fermentation index, polyphenols and amino acids of seeds of rambutan (Nephelium lappaceum L.) during fermentation. J. Food Compos. Anal. 2015, 37, 128–135. [Google Scholar] [CrossRef]
- Man, S.; Ma, J.; Wang, C.; Li, Y.; Gao, W.; Lu, F. Chemical composition and hypoglycaemic effect of polyphenol extracts from Litchi chinensis seeds. J. Funct. Foods 2016, 22, 313–324. [Google Scholar] [CrossRef]
- Paśko, P.; Galanty, A.; Zagrodzki, P.; Luksirikul, P.; Barasch, D.; Nemirovski, A.; Gorinstein, S. Dragon fruits as a reservoir of natural polyphenolics with chemopreventive properties. Molecules 2021, 26, 2158. [Google Scholar] [CrossRef] [PubMed]
- Lucas-González, R.; Fernández-López, J.; Pérez-Álvarez, J.Á.; Viuda-Martos, M. Effect of particle size on phytochemical composition and antioxidant properties of two persimmon flours from Diospyros kaki Thunb. vars. ‘Rojo Brillante’ and ‘Triumph’ co-products. J. Sci. Food Agric. 2018, 98, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Y.; He, X.M.; Sun, J.; Li, C.B.; Li, L.; Sheng, J.F.; Xin, M.; Li, Z.C.; Zheng, F.J.; Liu, G.M.; et al. Polyphenols and alkaloids in byproducts of longan fruits (Dimocarpus longan Lour.) and their bioactivities. Molecules 2019, 24, 1186. [Google Scholar] [CrossRef] [Green Version]
- Fidelis, M.; Santos, J.S.; Escher, G.B.; Vieira do Carmo, M.; Azevedo, L.; Cristina da Silva, M.; Putnik, P.; Granato, D. In vitro antioxidant and antihypertensive compounds from camu-camu (Myrciaria dubia McVaugh, Myrtaceae) seed coat: A multivariate structure-activity study. Food Chem. Toxicol. 2018, 120, 479–490. [Google Scholar] [CrossRef]
- De Azevêdo, J.C.S.; Fujita, A.; de Oliveira, E.L.; Genovese, M.I.; Correia, R.T.P. Dried camu-camu (Myrciaria dubia H.B.K. McVaugh) industrial residue: A bioactive-rich Amazonian powder with functional attributes. Food Res. Int. 2014, 62, 934–940. [Google Scholar] [CrossRef]
- Bittová, M.; Hladůvková, D.; Roblová, V.; Kráčmar, S.; Kubáň, P.; Kubáň, V. Analysis of organic acids, deacetyl asperulosidic acid and polyphenolic compounds as a potential tool for characterization of noni (Morinda citrifolia) products. Nat. Prod. Commun. 2015, 10, 1817–1820. [Google Scholar] [CrossRef] [Green Version]
- Jamaludin, R.; Kim, D.S.; Md Salleh, L.; Lim, S. Bin Optimization of high hydrostatic pressure extraction of bioactive compounds from noni fruits. J. Food Meas. Charact. 2020, 14, 2810–2818. [Google Scholar] [CrossRef]
- Martín Ortega, A.M.; Segura Campos, M.R. Bioactive compounds as Therapeutic alternatives. In Bioactive Compounds; Campos, M.R.S., Ed.; Woodhead Publishing: Duxford, UK, 2019; pp. 247–264. [Google Scholar]
- Pan, L.; Duan, Y.; Ma, F.; Lou, L. Punicalagin inhibits the viability, migration, invasion, and EMT by regulating GOLPH3 in breast cancer cells. J. Recept. Signal. Transduct. 2020, 40, 173–180. [Google Scholar] [CrossRef]
- Guo, X.D.; Zhang, D.Y.; Gao, X.J.; Parry, J.; Liu, K.; Liu, B.L.; Wang, M. Quercetin and quercetin-3-O-glucuronide are equally effective in ameliorating endothelial insulin resistance through inhibition of reactive oxygen species-associated inflammation. Mol. Nutr. Food Res. 2013, 57, 1037–1045. [Google Scholar] [CrossRef]
- Boo, Y.C. Emerging Strategies to Protect the Skin from Ultraviolet Rays Using Plant-Derived Materials. Antioxidants 2020, 9, 637. [Google Scholar] [CrossRef]
- Wojnar, W.; Zych, M.; Kaczmarczyk-Sedlak, I. Antioxidative effect of flavonoid naringenin in the lenses of type 1 diabetic rats. Biomed. Pharmacother. 2018, 108, 974–984. [Google Scholar] [CrossRef]
- Yang, S.; Zhou, J.; Li, D.; Shang, C.; Peng, L.; Pan, S. The structure-antifungal activity relationship of 5,7-dihydroxyflavonoids against Penicillium itallicum. Food Chem. 2017, 224, 26–31. [Google Scholar] [CrossRef]
- Bhadoriya, S.S.; Ganeshpurkar, A.; Bhadoriya, R.P.S.; Sahu, S.K.; Patel, J.R. Antidiabetic potential of polyphenolic-rich fraction of Tamarindus indica seed coat in alloxan-induced diabetic rats. J. Basic Clin. Physiol. Pharmacol. 2018, 29, 37–45. [Google Scholar] [CrossRef]
- Barros, L.; Calhelha, R.C.; Queiroz, M.J.R.P.; Santos-Buelga, C.; Santos, E.A.; Regis, W.C.B.; Ferreira, I.C.F.R. The powerful in vitro bioactivity of Euterpe oleracea Mart. seeds and related phenolic compounds. Ind. Crops Prod. 2015, 76, 318–322. [Google Scholar] [CrossRef] [Green Version]
- Tremocoldi, M.A.; Rosalen, P.L.; Franchin, M.; Massarioli, A.P.; Denny, C.; Daiuto, É.R.; Paschoal, J.A.R.; Melo, P.S.; De Alencar, S.M. Exploration of avocado by-products as natural sources of bioactive compounds. PLoS ONE 2018, 13, e0192577. [Google Scholar] [CrossRef] [Green Version]
- Powar, A.; Gupta, A.; Kamble, S.; Shinde, B. Immunosuppressive and antidiabetic activity of polyphenols from Musa paradisiaca. Eur. J. Biotech. Biosci. 2017, 5, 62–65. [Google Scholar]
- Rossin, D.; Barbosa-Pereira, L.; Iaia, N.; Testa, G.; Sottero, B.; Poli, G.; Zeppa, G.; Biasi, F. A dietary mixture of oxysterols induces in vitro intestinal inflammation through TLR2/4 activation: The protective effect of cocoa bean shells. Antioxidants 2019, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Youryon, P.; Supapvanich, S. Physicochemical quality and antioxidant changes in ‘Leb Mue Nang’ banana fruit during ripening. Agric. Nat. Resour. 2017, 51, 47–52. [Google Scholar] [CrossRef]
- Fragoso, M.F.; Romualdo, G.R.; Vanderveer, L.A.; Franco-Barraza, J.; Cukierman, E.; Clapper, M.L.; Carvalho, R.F.; Barbisan, L.F. Lyophilized açaí pulp (Euterpe oleracea Mart) attenuates colitis-associated colon carcinogenesis while its main anthocyanin has the potential to affect the motility of colon cancer cells. Food Chem. Toxicol. 2018, 121, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Enayati, A.; Yassa, N.; Mazaheri, Z.; Rajaei, M.; Pourabouk, M.; Ghorghanlu, S.; Basiri, S.; Khori, V. Cardioprotective and anti-apoptotic effects of Potentilla reptans L. root via Nrf2 pathway in an isolated rat heart ischemia/reperfusion model. Life Sci. 2018, 215, 216–226. [Google Scholar] [CrossRef]
- Sheela, D.L.; Nazeem, P.A.; Narayanankutty, A.; Shylaja, R.M.; Davis, S.P.; James, P.; Valsalan, R.; Devassy Babu, T.; Raghavamenon, A.C. Coconut phytocompounds inhibits polyol pathway enzymes: Implication in prevention of microvascular diabetic complications. Prostaglandins Leukot. Essent. Fat. Acids 2017, 127, 20–24. [Google Scholar] [CrossRef]
- Sgarbossa, A.; Giacomazza, D.; di Carlo, M. Ferulic acid: A hope for alzheimer’s disease therapy from plants. Nutrients 2015, 7, 5764–5782. [Google Scholar] [CrossRef]
- Verma, S.; Singh, A.; Mishra, A. Gallic acid: Molecular rival of cancer. Environ. Toxicol. Pharmacol. 2013, 35, 473–485. [Google Scholar] [CrossRef]
- Dludla, P.V.; Nkambule, B.B.; Jack, B.; Mkandla, Z.; Mutize, T.; Silvestri, S.; Orlando, P.; Tiano, L.; Louw, J.; Mazibuko-Mbeje, S.E. Inflammation and oxidative stress in an obese state and the protective effects of gallic acid. Nutrients 2019, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. RSC Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- BenSaad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-inflammatory potential of ellagic acid, gallic acid and punicalagin A&B isolated from Punica granatum. BMC Complement. Altern. Med. 2017, 17, 47. [Google Scholar] [CrossRef] [Green Version]
- Gold-Smith, F.; Fernandez, A.; Bishop, K. Mangiferin and cancer: Mechanisms of action. Nutrients 2016, 8, 396. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.T.; Wang, Z.Z.; Yuan, Y.H.; Sun, H.M.; Chen, N.H.; Zhang, Y. Mangiferin: A multipotent natural product preventing neurodegeneration in Alzheimer’s and Parkinson’s disease models. Pharmacol. Res. 2019, 146, 104336. [Google Scholar] [CrossRef]
- Maurmann, N.; De Farias, C.B.; Schwartsmann, G.; Roesler, R.; Delgado-Hernández, R.; Pardo-Andreu, G.L. Mangifera indica L. extract (vimang) improves the aversive memory in spinocerebellar ataxia type 2 transgenic mice. J. Pharm. Pharmacogn. Res. 2014, 2, 63–72. [Google Scholar]
- Canuto, K.M. Propriedades Químicas e Farmacológicas de Mangiferina: Um Composto Bioativo de Manga (Mangifera indica L.); Embrapa: Brasilia, Brasil, 2009; pp. 10–11. [Google Scholar]
- Garrido, G.; González, D.; Lemus, Y.; García, D.; Lodeiro, L.; Quintero, G.; Delporte, C.; Núñez-Sellés, A.J.; Delgado, R. In vivo and in vitro anti-inflammatory activity of Mangifera indica L. extract (VIMANG®). Pharmacol. Res. 2004, 50, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Hsun-Wei Huang, T.; Peng, G.; Qian Li, G.; Yamahara, J.; Roufogalis, B.D.; Li, Y. Salacia oblonga root improves postprandial hyperlipidemia and hepatic steatosis in Zucker diabetic fatty rats: Activation of PPAR-α. Toxicol. Appl. Pharmacol. 2006, 210, 225–235. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Magalhães, T.S.S.; de Oliveira Macedo, P.C.; Converti, A.; Neves de Lima, Á.A. The use of Euterpe oleracea Mart. as a new perspective for disease treatment and prevention. Biomolecules 2020, 10, 813. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira Vasconcelos, M.; Freitas Mota, E.; Gomes-Rochette, N.F.; Sousa Nunes-Pinheiro, D.C.; Nabavi, S.M.; Fernandes de Melo, D. Açai or Brazilian Berry (Euterpe oleracea). In Nonvitamin and Nonmineral Nutritional Supplements; Nabavi, S.M., Sanches Silva, A., Eds.; Academic Press: Oxford, UK, 2019; pp. 131–133. [Google Scholar] [CrossRef]
- Martinez, R.M.; De Almeida Bauer Guimarães, D.; Berniz, C.R.; De Abreu, J.P.; Da Rocha, A.P.M.; De Moura, R.S.; Resende, A.C.; Teodoro, A.J. Açai (Euterpe oleracea Mart.) seed extract induces cell cycle arrest and apoptosis in human lung carcinoma cells. Foods 2018, 7, 178. [Google Scholar] [CrossRef] [Green Version]
- De Liz, S.; Cardoso, A.L.; Copetti, C.L.K.; de Fragas Hinnig, P.; Vieira, F.G.K.; da Silva, E.L.; Schulz, M.; Fett, R.; Micke, G.A.; Di Pietro, P.F. Açaí (Euterpe oleracea Mart.) and juçara (Euterpe edulis Mart.) juices improved HDL-c levels and antioxidant defense of healthy adults in a 4-week randomized cross-over study. Clin. Nutr. 2020, 39, 3629–3636. [Google Scholar] [CrossRef]
- Aranha, L.N.; Silva, M.G.; Uehara, S.K.; Luiz, R.R.; Nogueira Neto, J.F.; Rosa, G.; Moraes de Oliveira, G.M. Effects of a hypoenergetic diet associated with açaí (Euterpe oleracea Mart.) pulp consumption on antioxidant status, oxidative stress and inflammatory biomarkers in overweight, dyslipidemic individuals. Clin. Nutr. 2020, 39, 1464–1469. [Google Scholar] [CrossRef]
- Pala, D.; Barbosa, P.O.; Silva, C.T.; de Souza, M.O.; Freitas, F.R.; Volp, A.C.P.; Maranhão, R.C.; de Freitas, R.N. Açai (Euterpe oleracea Mart.) dietary intake affects plasma lipids, apolipoproteins, cholesteryl ester transfer to high-density lipoprotein and redox metabolism: A prospective study in women. Clin. Nutr. 2018, 37, 618–623. [Google Scholar] [CrossRef]
- Choi, Y.J.; Choi, Y.J.; Kim, N.; Nam, R.H.; Lee, S.; Lee, H.S.; Lee, H.N.; Surh, Y.J.; Lee, D.H. Açaí berries inhibit colon tumorigenesis in azoxymethane/dextran sulfate sodium-treated mice. Gut Liver 2017, 11, 243–252. [Google Scholar] [CrossRef]
- Martins, G.R.; do Amaral, F.R.L.; Brum, F.L.; Mohana-Borges, R.; de Moura, S.S.T.; Ferreira, F.A.; Sangenito, L.S.; Santos, A.L.S.; Figueiredo, N.G.; da Silva, A.S.A. Chemical characterization, antioxidant and antimicrobial activities of açaí seed (Euterpe oleracea Mart.) extracts containing A- and B-type procyanidins. LWT Food Sci. Tecnol. 2020, 132, 109830. [Google Scholar] [CrossRef]
- Topalović, A.; Knežević, M.; Gačnik, S.; Mikulic-Petkovsek, M. Detailed chemical composition of juice from autochthonous pomegranate genotypes (Punica granatum L.) grown in different locations in Montenegro. Food Chem. 2020, 330, 127261. [Google Scholar] [CrossRef]
- Grabež, M.; Škrbić, R.; Stojiljković, M.P.; Rudić-Grujić, V.; Paunović, M.; Arsić, A.; Petrović, S.; Vučić, V.; Mirjanić-Azarić, B.; Šavikin, K.; et al. Beneficial effects of pomegranate peel extract on plasma lipid profile, fatty acids levels and blood pressure in patients with diabetes mellitus type-2: A randomized, double-blind, placebo-controlled study. J. Funct. Foods 2020, 64, 103692. [Google Scholar] [CrossRef]
- Farooqi, A.A. Regulation of deregulated cell signaling pathways by pomegranate in different cancers: Re-interpretation of knowledge gaps. Semin. Cancer Biol. 2021, 73, 294–301. [Google Scholar] [CrossRef]
- Sohrab, G.; Roshan, H.; Ebrahimof, S.; Nikpayam, O.; Sotoudeh, G.; Siasi, F. Effects of pomegranate juice consumption on blood pressure and lipid profile in patients with type 2 diabetes: A single-blind randomized clinical trial. Clin. Nutr. ESPEN 2019, 29, 30–35. [Google Scholar] [CrossRef]
- Nasser, M.; Damaj, Z.; Hijazi, A.; Merah, O.; Al-Khatib, B.; Hijazi, N.; Trabolsi, C.; Damaj, R.; Nasser, M. Pomegranate juice extract decreases cisplatin toxicity on peripheral blood mononuclear cells. Medicines 2020, 7, 66. [Google Scholar] [CrossRef]
- Karwasra, R.; Singh, S.; Sharma, D.; Sharma, S.; Sharma, N.; Khanna, K. Pomegranate supplementation attenuates inflammation, joint dysfunction via inhibition of NF-κB signaling pathway in experimental models of rheumatoid arthritis. J. Food Biochem. 2019, 43, e12959. [Google Scholar] [CrossRef]
- Stojanović, I.; Šavikin, K.; Đedović, N.; Živković, J.; Saksida, T.; Momčilović, M.; Koprivica, I.; Vujičić, M.; Stanisavljević, S.; Miljković, D.; et al. Pomegranate peel extract ameliorates autoimmunity in animal models of multiple sclerosis and type 1 diabetes. J. Funct. Foods. 2017, 35, 522–530. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Damasceno, D.C.; Machado Spada, A.P.; Lima, I.E.R.N.; Cressoni Araújo, A.; Guiguer, E.L.; Martuchi, K.A.; Oshiiwa, M.; Mendes, C.G. Effects of Passiflora edulis on the Metabolic Profile of Diabetic Wistar Rat Offspring. J. Med. Food 2011, 14, 1490–1495. [Google Scholar] [CrossRef] [Green Version]
- Ballesteros-Vivas, D.; Alvarez-Rivera, G.; León, C.; Morantes, S.J.; Ibánez, E.; Parada-Alfonso, F.; Cifuentes, A.; Valdés, A. Foodomics evaluation of the anti-proliferative potential of Passiflora mollissima seeds. Food Res. Int. 2020, 130, 108938. [Google Scholar] [CrossRef]
- Fernandes Marques, S.S.; Feio Libonati, R.M.; Sabaa-Srur, A.U.O.; Luo, R.; Shejwalkar, P.; Hara, K.; Dobbs, T.; Smith, R.E. Evaluation of the effects of passion fruit peel flour (Passiflora edulis fo. flavicarpa) on metabolic changes in HIV patients with lipodystrophy syndrome secondary to antiretroviral therapy. Rev. Bras. Farmacogn. 2016, 26, 420–426. [Google Scholar] [CrossRef]
- Kasala, E.R.; Bodduluru, L.N.; Barua, C.C.; Madhana, R.M.; Dahiya, V.; Budhani, M.K.; Mallugari, R.R.; Maramreddy, S.R.; Gogoi, R. Chemopreventive effect of chrysin, a dietary flavone against benzo(a)pyrene induced lung carcinogenesis in Swiss albino mice. Pharmacol. Rep. 2016, 68, 310–318. [Google Scholar] [CrossRef]
- Maryanto, S.; Marsono, Y. The effect of guava on the improvement of lipid profile in hypercholesterolemic rats. IOP Conf. Ser. Earth Environ. Sci. 2019, 276, 012054. [Google Scholar] [CrossRef]
- Shabbir, H.; Kausar, T.; Noreen, S.; ur Rehman, H.; Hussain, A.; Huang, Q.; Gani, A.; Su, S.; Nawaz, A. In vivo screening and antidiabetic potential of polyphenol extracts from guava pulp, seeds and leaves. Animals 2020, 10, 1714. [Google Scholar] [CrossRef]
- Feng, X.; Wang, Z.; Meng, D.; Li, X. Cytotoxic and antioxidant constituents from the leaves of Psidium guajava. Bioorg. Med. Chem. Lett. 2015, 25, 2193–2198. [Google Scholar] [CrossRef]
- Correa, M.G.; Couto, J.S.; Trindade, B.B.; Abreu, J.P.; Nakajima, V.M.; Oliveira, F.L.; Farah, A.; Teodoro, A.J. Antiproliferative effect of guava fruit extracts in MDA-MB-435 and MCF-7 human breast cancer cell lines. An. Acad. Bras. Cienc. 2020, 92, e20191500. [Google Scholar] [CrossRef]
- Li, P.-Y.; Hsu, C.-C.; Yin, M.-C.; Kuo, Y.-H.; Tang, F.-Y.; Chao, C.-Y. Protective Effects of Red Guava on Inflammation and Oxidative Stress in Streptozotocin-Induced Diabetic Mice. Molecules 2015, 20, 22341–22350. [Google Scholar] [CrossRef] [Green Version]
- Saxena, P.; Panjwani, D. Cardioprotective potential of hydro-alcoholic fruit extract of Ananas comosus against isoproterenol induced myocardial infraction in Wistar Albino rats. J. Acute. Dis. 2014, 3, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Seenak, P.; Kumphune, S.; Malakul, W.; Chotima, R.; Nernpermpisooth, N. Pineapple consumption reduced cardiac oxidative stress and inflammation in high cholesterol diet-fed rats. Nutr. Metab. 2021, 18, 36. [Google Scholar] [CrossRef]
- Kumar, M.; Saurabh, V.; Tomar, M.; Hasan, M.; Changan, S.; Sasi, M.; Maheshwari, C.; Prajapati, U.; Singh, S.; Prajapat, R.K.; et al. Mango (Mangifera indica L.) leaves: Nutritional composition, phytochemical profile, and health-promoting bioactivities. Antioxidants 2021, 10, 299. [Google Scholar] [CrossRef]
- Alañón, M.E.; Pimentel-Moral, S.; Arráez-Román, D.; Segura-Carretero, A. HPLC-DAD-Q-ToF-MS profiling of phenolic compounds from mango (Mangifera indica L.) seed kernel of different cultivars and maturation stages as a preliminary approach to determine functional and nutraceutical value. Food Chem. 2021, 337, 127764. [Google Scholar] [CrossRef]
- Imran, M.; Arshad, M.S.; Butt, M.S.; Kwon, J.H.; Arshad, M.U.; Sultan, M.T. Mangiferin: A natural miracle bioactive compound against lifestyle related disorders. Lipids Health Dis. 2017, 16, 84. [Google Scholar] [CrossRef]
- Morozkina, S.N.; Vu, T.H.N.; Generalova, Y.E.; Snetkov, P.P.; Uspenskaya, M.V. Mangiferin as new potential anti-cancer agent and mangiferin-integrated polymer systems: A novel research direction. Biomolecules 2021, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Wall-Medrano, A.; Olivas-Aguirre, F.J.; Ayala-Zavala, J.F.; Domínguez-Avila, J.A.; Gonzalez-Aguilar, G.A.; Herrera-Cazares, L.A.; Gaytan-Martinez, M. Health benefits of mango by-products. Food Wastes By-Prod. 2020, 159–191. [Google Scholar] [CrossRef]
- Figueroa, J.G.; Borrás-Linares, I.; Lozano-Sánchez, J.; Segura-Carretero, A. Comprehensive characterization of phenolic and other polar compounds in the seed and seed coat of avocado by HPLC-DAD-ESI-QTOF-MS. Food Res. Int. 2018, 105, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.; Billing, D.; Connolly, P.; Smith, W.; Cooney, J.; Burdon, J. Maturity, storage and ripening effects on anti-fungal compounds in the skin of ‘Hass’ avocado fruit. Postharvest Biol. Technol. 2018, 146, 43–50. [Google Scholar] [CrossRef]
- Gómez-Maqueo, A.; Escobedo-Avellaneda, Z.; Welti-Chanes, J. Phenolic compounds in mesoamerican fruits—Characterization, health potential and processing with innovative technologies. Int. J. Mol. Sci. 2020, 21, 8357. [Google Scholar] [CrossRef]
- Ervianingsih; Hurria; Astari, C.; Mursyid, M.; Kadir, A.R. Avocado seed waste utilization into biscuits as diabetes mellitus patients’ alternative foods. IOP Conf. Ser. Earth Environ. Sci. 2020, 575, 3–8. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.M.; Ullah, M.O. Hydroxycinnamic acid derivatives: A potential class of natural compounds for the management of lipid metabolism and obesity. Nutr. Metab. 2016, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Eldaim, M.A.; Tousson, E.; El Sayed, I.E.T.; Abd El-Aleim, A.E.A.H.; Elsharkawy, H.N. Grape seeds proanthocyanidin extract ameliorates Ehrlich solid tumor induced renal tissue and DNA damage in mice. Biomed. Pharmacother. 2019, 115, 108908. [Google Scholar] [CrossRef]
- Maisuria, V.B.; Okshevsky, M.; Déziel, E.; Tufenkji, N. Proanthocyanidin Interferes with Intrinsic Antibiotic Resistance Mechanisms of Gram-Negative Bacteria. Adv. Sci. 2019, 6, 1802333. [Google Scholar] [CrossRef]
- Kamaraj, M.; Dhana Rangesh Kumar, V.; Nithya, T.G.; Danya, U. Assessment of Antioxidant, Antibacterial Activity and Phytoactive Compounds of Aqueous Extracts of Avocado Fruit Peel from Ethiopia. Int. J. Pept. Res. Ther. 2020, 26, 1549–1557. [Google Scholar] [CrossRef]
- Quiles, J.L.; Rivas-García, L.; Varela-López, A.; Llopis, J.; Battino, M.; Sánchez-González, C. Do nutrients and other bioactive molecules from foods have anything to say in the treatment against COVID-19? Environ. Res. 2020, 191, 110053. [Google Scholar] [CrossRef]
- Russo, M.; Moccia, S.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Roles of flavonoids against coronavirus infection. Chem. Biol. Interact. 2020, 328, 109211. [Google Scholar] [CrossRef]
- Wu, Y.H.; Tseng, C.K.; Wu, H.C.; Wei, C.K.; Lin, C.K.; Chen, I.S.; Chang, H.S.; Lee, J.C. Avocado (Persea americana) fruit extract (2R,4R)-1,2,4-trihydroxyheptadec-16-yne inhibits dengue virus replication via upregulation of NF-κB–dependent induction of antiviral interferon responses. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Martins, C.M.; Ferro, D.M.; de Brito, E.S.; Ferreira, S.R.S. Industrial relevance of Tamarindus indica L. by-products as source of valuable active metabolites. Innov. Food Sci. Emerg. Technol. 2020, 66, 102518. [Google Scholar] [CrossRef]
- Menezes, A.P.P.; Trevisan, S.C.C.; Barbalho, S.M.; Guiguer, E.L. Tamarindus indica L. A plant with multiple medicinal purposes. J. Pharmacogn. Phytochem. 2016, 5, 50–54. [Google Scholar]
- Martinello, F.; Kannen, V.; Franco, J.J.; Gasparotto, B.; Sakita, J.Y.; Sugohara, A.; Garcia, S.B.; Uyemura, S.A. Chemopreventive effects of a Tamarindus indica fruit extract against colon carcinogenesis depends on the dietary cholesterol levels in hamsters. Food Chem. Toxicol. 2017, 107, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Ghosh, U. Optimization of the extraction of natural phenolic antioxidants from the seeds of Tamarindus indica L.—An undervalued by product of food processing—Using response surface methodology. Acta Biol. Szeged. 2018, 62, 67–74. [Google Scholar] [CrossRef]
- Nikooei, P.; Hosseinzadeh-Attar, M.J.; Asghari, S.; Norouzy, A.; Yaseri, M.; Vasheghani-Farahani, A. Effects of virgin coconut oil consumption on metabolic syndrome components and asymmetric dimethylarginine: A randomized controlled clinical trial. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 939–949. [Google Scholar] [CrossRef]
- Maduwanthi, S.D.T.; Marapana, R.A.U.J. Total phenolics, flavonoids and antioxidant activity following simulated gastro-intestinal digestion and dialysis of banana (Musa acuminata, AAB) as affected by induced ripening agents. Food Chem. 2021, 339, 127909. [Google Scholar] [CrossRef]
- Sidhu, J.S.; Zafar, T.A. Bioactive compounds in banana fruits and their health benefits. Food Qual. Saf. 2018, 2, 183–188. [Google Scholar] [CrossRef]
- Sansone, R.; Rodriguez-Mateos, A.; Heuel, J.; Falk, D.; Schuler, D.; Wagstaff, R.; Kuhnle, G.G.C.; Spencer, J.P.E.; Schroeter, H.; Merx, M.W.; et al. Cocoa flavanol intake improves endothelial function and Framingham Risk Score in healthy men and women: A randomised, controlled, double-masked trial: The Flaviola Health Study. Br. J. Nutr. 2015, 114, 1246–1255. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, I.; Li, A.; Manson, J.A.E.; Sesso, H.D.; Wang, L.; Liu, S. Cocoa flavanol intake and biomarkers for cardiometabolic health: A systematic review and meta-analysis of randomized controlled trials. J. Nutr. 2016, 146, 2325–2333. [Google Scholar] [CrossRef] [Green Version]
- Davinelli, S.; Corbi, G.; Zarrelli, A.; Arisi, M.; Calzavara-Pinton, P.; Grassi, D.; De Vivo, I.; Scapagnini, G. Short-term supplementation with flavanol-rich cocoa improves lipid profile, antioxidant status and positively influences the AA/EPA ratio in healthy subjects. J. Nutr. Biochem. 2018, 61, 33–39. [Google Scholar] [CrossRef]
- Álvarez-Cilleros, D.; López-Oliva, M.E.; Ramos, S.; Martín, M.Á. Preventive effect of cocoa flavanols against glucotoxicity-induced vascular inflammation in the arteria of diabetic rats and on the inflammatory process in TNF-α-stimulated endothelial cells. Food Chem. Toxicol. 2020, 146, 111824. [Google Scholar] [CrossRef]
- Vázquez-González, Y.; Ragazzo-Sánchez, J.A.; Calderón-Santoyo, M. Characterization and antifungal activity of jackfruit (Artocarpus heterophyllus Lam.) leaf extract obtained using conventional and emerging technologies. Food Chem. 2020, 330, 127211. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fruit | Source | Flavonoids | Non-Flavonoids Phenolics | Ref. |
|---|---|---|---|---|
| Kiwi (Actinidia arguta (Siebold & Zucc.) Planch. ex Miq) | Pulp | Rutin (0.33 mg/g dw) and kaempherol-3-O-galactoside (0.25 mg/g dw) | Crypto-chlorogenic acid (0.10 mg/g dw) and neochlorogenic acid (0.08 mg/g dw) | [81] |
| Acerola (Malpighia emarginata D.C.) | Pulp | Keampferol-3-O-glucoside (0.33 mg/g dw) and isorhamnetin (0.11 mg/g dw) | p-coumaric acid (0.29 mg/g dw) | [82] |
| Acerola (Malpighia emarginata D.C.) | Pulp and peel | Myricetin (0.94 mg/g dw) and rutin (0.61 mg/g dw) | 2,5-dihydroxybenzoic acid (7.37 mg/g dw) and 3,4-dihydroxybenzoic acid (0.98 mg/g dw) | [83] |
| Jabuticaba (Myrciaria jaboticaba (Vell.) O.Berg) | Peel | Cyanidin-3-O-glucoside (7.25 mg/g dw) and quercetin-3-O-rutinoside (1.17 mg/g dw) | Ellagic acid (0.23 mg/g dw) and gallic acid (0.13 mg/g dw) | [84] |
| Jabuticaba (Myrciaria jaboticaba (Vell.) O.Berg) | Seeds | Castalagin (0.13 mg/g dw) and vescalagin (0.69 mg/g dw) | Ellagic acid (3.80 mg/g dw) and gallic acid (2.30 mg/g dw) | [85] |
| Rambutan (Nephelium lappaceum L.) | Seeds | Geraniin (0.42 mg/g dw) and corilagin (0.09 mg/g dw) | Ellagic acid (0.46 mg/g dw) and gallic (0.10 mg/g dw) | [86] |
| Lychee (Litchi chinensis Sonn) | Seeds | Rutin (0.098 mg/100 g dw) and scopoletin (0.076 mg/100 g dw) | N.D. | [87] |
| Dragon fruit (Hylocereus spp.) | Pulp | Myricetin (0.18 mg/g dw) and quercetin (0.06 mg/g dw) | Gallic acid (0.23 mg/g dw) | [88] |
| Kaki (Dyospiros kaki L.) | Pulp | Epicatechin (0.03 mg/g dw) and catechin (0.02 mg/g dw) | Gallic acid (2.96 mg/g dw) | [89] |
| Longan (Dimocarpus longan Lour) | Peel | Corilagin (2.15 mg/g dw) | Ellagic acid (0.08 mg/g dw) and gallic acid (0.18 mg/g dw) | [90] |
| Longan (Dimocarpus longan Lour) | Seed coat | Corilagin (5.53 mg/g dw) | Ellagic acid (0.83 mg/g dw) and gallic (0.49 mg/g dw) | [91] |
| Camu-camu (Myrciaria dubia McVaugh) | Seed coat | Rutin (0.13 mg/g dw) | Ferulic acid (0.59 mg/g dw) and p-coumaric acid (0.49 mg/g dw) | [92] |
| Camu-camu (Myrciaria dubia McVaugh) | Pulp | Catechin (0.12 mg/g dw) and myricetin (0.02 mg/g dw) | Ellagic acid (0.19 mg/g dw) and syringic acid (0.07 mg/g dw) | [93] |
| Noni (Morinda citrifolia L.) | Pulp | Catechin (1.11 mg/g dw), rutin (0.40 mg/g dw) and quercetin (0.27 mg/g dw) | Gallic acid (0.24 mg/g dw) | [94] |
| Noni (Morinda citrifolia L.) | Pulp | Scopoletin (0.53 mg/g dw) and rutin (0.54 mg/g dw) | N.D. | [95] |
| Polyphenolic Compound | Biological Activity | Mechanism | Ref. |
|---|---|---|---|
| Tannin | |||
| Punicalagin | Anticancer | Inhibition of the invasion, migration, and viability of breast cancer cells. Decreases the expression of N-Cadherin, Golgi phosphoprotein 3, matrix metalloproteinase-2, matrix metalloproteinase-9 | [97] |
| Flavonols | |||
| Quercetin-3-O-glucuronide | Anti-inflammatory | Decreases in ROS-associated inflammation by inhibition of interleukin-6 and tumor necrosis factor-α production with suppression of IKKβ/NF-κB phosphorylation. | [98] |
| Antidiabetic | Facilitates the PI3K signaling by positive regulation of serine/tyrosine phosphorylation of insulin receptor substrate-1 (IRS-1). | ||
| Quercetin | Photoprotective | Absorbs UV radiation, inhibits UV-induced inflammation in primary keratinocytes, and inhibits skin damage produced by UVB rays. | [99] |
| Naringenin | Antioxidative | Protects ocular neurons from degeneration, reverses retinal pigment epithelium degeneration and laser-induced choroidal neovascularization | [100] |
| Flavone | |||
| Artocarpesin | Anti-inflammatory | Suppresses the LPS-induced production of nitric oxide (NO) and prostaglandin E2 | [80] |
| Baicalein | Antifungal | Inhibition of outflow pumps, since they decreased their efficiency, which eventually led to cell death or apoptosis. | [100] |
| Flavan-3-ols | |||
| Benzo-tropolona | Anticancer | Reduces the viability of human breast (MCF7), lung (H1299), colon (HT29), and prostate (LNCaP) cancer cells in vitro. After 12 h treatment on a model in vivo the G0/G1 phase cell cycle arrests in LNCaP cells. | [17] |
| Antioxidant | Reduces lipid hydroperoxide formation in an oil-in-water emulsion (33% reduction at 500 μg/mL). | [17] | |
| Antiangiogenic | Inhibition of kinase proteins. | [70] | |
| Catechin | Antidiabetic | Protective effect on pancreatic β cells Increased glucose absorption. Reduces the blood glucose level. | [101] |
| Catechins and procyanidins | Antitumor | Inhibitory effect against cervical carcinoma cell | [102] |
| Catechin and epicatechin | Antioxidant | Stabilizes peroxyl radicals, superoxide radical and hypochlorous reactive species. | [103] |
| Epicatechin | Antiproliferative capacity | Immunosuppressive effect on T cell proliferation | [104] |
| Anti-inflammatory | Prevention of oxysterol mixture-induced InterLeukin-8 release (pro-inflammatory cytokine) | [105] | |
| Gallo-catechin | Antioxidant | DPPH radical scavenging activity | [106] |
| Anthocyanins | |||
| Cyanidin glycosides | Anticancer | Decreases the total number of aberrant crypt foci, aberrant crypt foci multiplicity, tumor cell proliferation and incidence of tumors with high grade dysplasia | [107] |
| Pro-anthocyanins | |||
| Resveratrol | Cardioprotective | Effects via nitric oxide release, Nrf2 pathway, and antioxidant activity. | [108] |
| Procyanidin B2 | Anti-obesity | Reduction of lipid accumulation in a dose dependent-manner. Hypolipidemic activity. | [66] |
| Antioxidant | Scavenges radicals and inhibits xanthine oxidase. | [66] | |
| Hydroxycinnamic acids | |||
| Caffeic acid | Antidiabetic | Prevents diabetic complications specifically inhibiting aldose reductase dependent polyol pathway. | [109] |
| Ferulic acid | Neurological | Interactions with fibrils Aβ predominantly by hydrogen bonding with His14 and Glu22, interfering with the formation of β-sheets, thereby inhibiting the formation of aggregates. | [110] |
| Hydroxybenzoic acids | |||
| Gallic acid | Chemo-preventive | Inhibition of activation of nuclear factor-Band Akt signaling pathways along with the activity of COX, ribonucleotide reductase and GSH. Activates ATM kinase signaling pathways to prevent the processes of carcinogenesis | [111] |
| Anti-obesity | Inhibition of myeloperoxidase release and activity, scavenging of superoxide anions, as well as a possible interference with the assembly of active NADPH-oxidase, may account for the inhibition of the inflammatory process. | [112] | |
| Anti-inflammatory | Inhibition of myeloperoxidase activity, hence directly inhibiting the production of hypochlorous acid, and scavenging the hazardous reactive species produced by the enzyme. | [113] | |
| Antiallergic | Blocks the release of histamine, which would otherwise result in immediate hypersensitivity. | [114] | |
| Ellagic acid | Anti-inflammatory | Improves the nitric oxide levels in peritoneal macrophages and attenuate the levels of interleukin-17 and interferon gamma in mesenteric lymph node cells obtained from mice with type 1 diabetes | [115] |
| Xanthone | |||
| Mangiferin | Chemo-preventive | Decreases in the cell cycle by reducing proliferation, metastasis and promoting apoptosis of malignant cells. | [116] |
| Neuroprotective | Up-regulates dopamine concentrations and diminishes neurotoxin’s effects which involve oxidative stress, mitochondrial dysfunction, and apoptosis. | [117] | |
| Improved object recognition memory in healthy rats through a mechanism that might involve an increase in neurotrophin and cytokine levels. | [118] | ||
| Cardioprotective | Increases the concentration of phospholipids in cardiac tissue and restore the activity of antioxidant enzymes: superoxide dismutase, catalase, glutathione peroxidase, glutathione transferase and glutathione reductase. | [119] | |
| Anti-inflammatory | Inhibits the enzyme phospholipase A2, in human synovial secretion, and suppresses the release of inflammatory mediators prostaglandin E2 and leukotriene B4 by macrophages. | [120] | |
| Antidiabetic | Reduction of hyperglycemia and atherogenicity in diabetic rats through a significant decrease in blood glucose levels as well as a decrease in serum concentrations of triglycerides, total cholesterol and low-density lipoprotein cholesterol. | [121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayago-Ayerdi, S.; García-Martínez, D.L.; Ramírez-Castillo, A.C.; Ramírez-Concepción, H.R.; Viuda-Martos, M. Tropical Fruits and Their Co-Products as Bioactive Compounds and Their Health Effects: A Review. Foods 2021, 10, 1952. https://doi.org/10.3390/foods10081952
Sayago-Ayerdi S, García-Martínez DL, Ramírez-Castillo AC, Ramírez-Concepción HR, Viuda-Martos M. Tropical Fruits and Their Co-Products as Bioactive Compounds and Their Health Effects: A Review. Foods. 2021; 10(8):1952. https://doi.org/10.3390/foods10081952
Chicago/Turabian StyleSayago-Ayerdi, Sonia, Diana Laura García-Martínez, Ailin Cecilia Ramírez-Castillo, Heidi Rubí Ramírez-Concepción, and Manuel Viuda-Martos. 2021. "Tropical Fruits and Their Co-Products as Bioactive Compounds and Their Health Effects: A Review" Foods 10, no. 8: 1952. https://doi.org/10.3390/foods10081952