
Differences in Antioxidants, Polyphenols, Protein Digestibility and Nutritional Profile between Ganoderma lingzhi from Industrial Crops in Asia and Ganoderma lucidum from Cultivation and Iberian Origin

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Samples and Mycelium
2.2. Cultivation
2.2.1. Spawn Production
2.2.2. Preparation of Cultivation Substrates
2.2.3. Inoculation, Growing and Recollection
2.3. Preparation of Samples for Analysis
2.4. Nutritional Analysis
2.5. Analysis of Content in Total Phenols and Antioxidant Capacity
2.6. Protein Digestibility and Electrophoretic Profile
2.7. Analysis of Data
3. Results
3.1. Cultivation
3.2. Nutritional Analysis
3.3. Fatty Acids Analysis
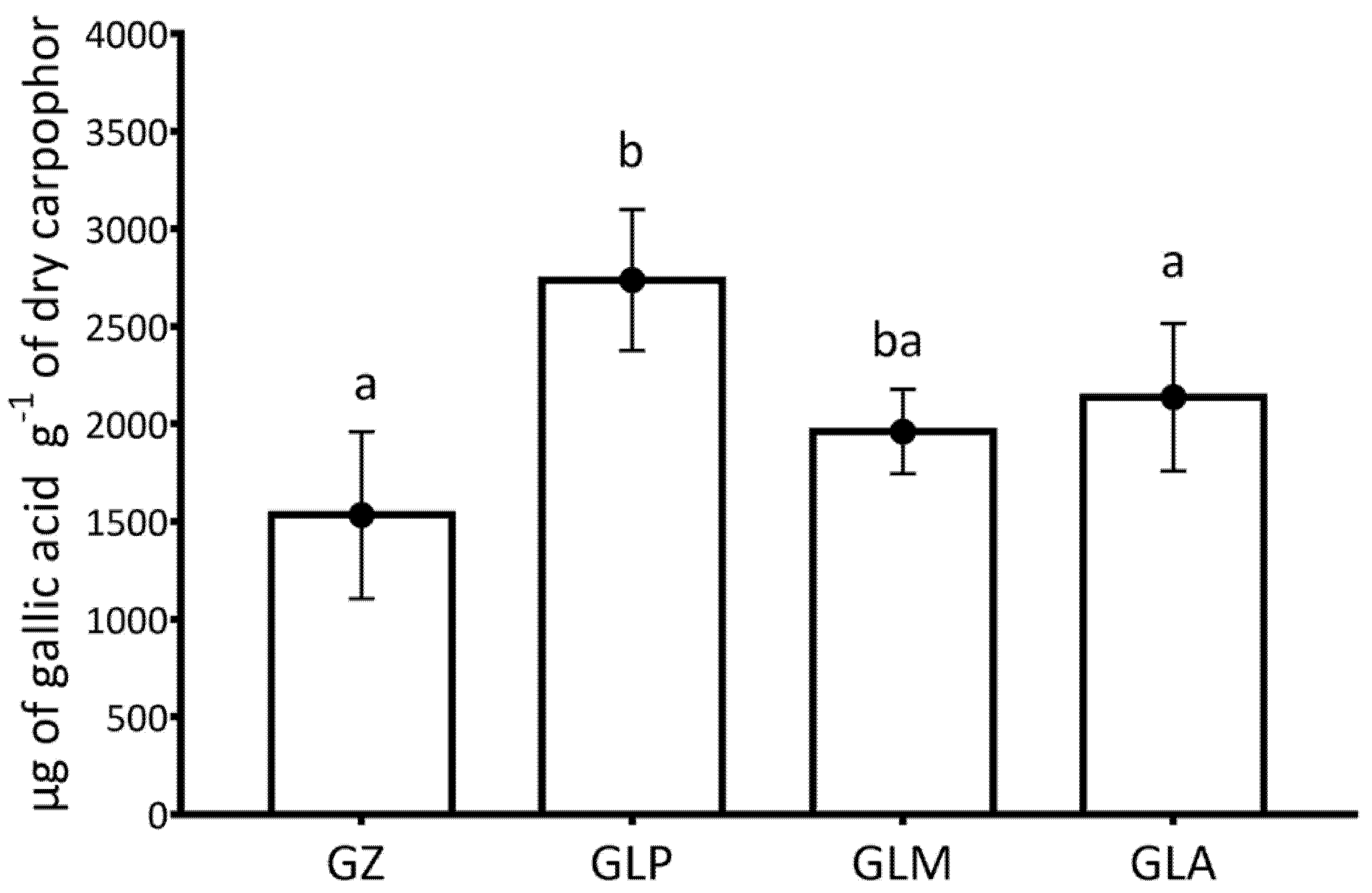
3.4. Analysis of Content in Total Phenols and Antioxidant Capacity
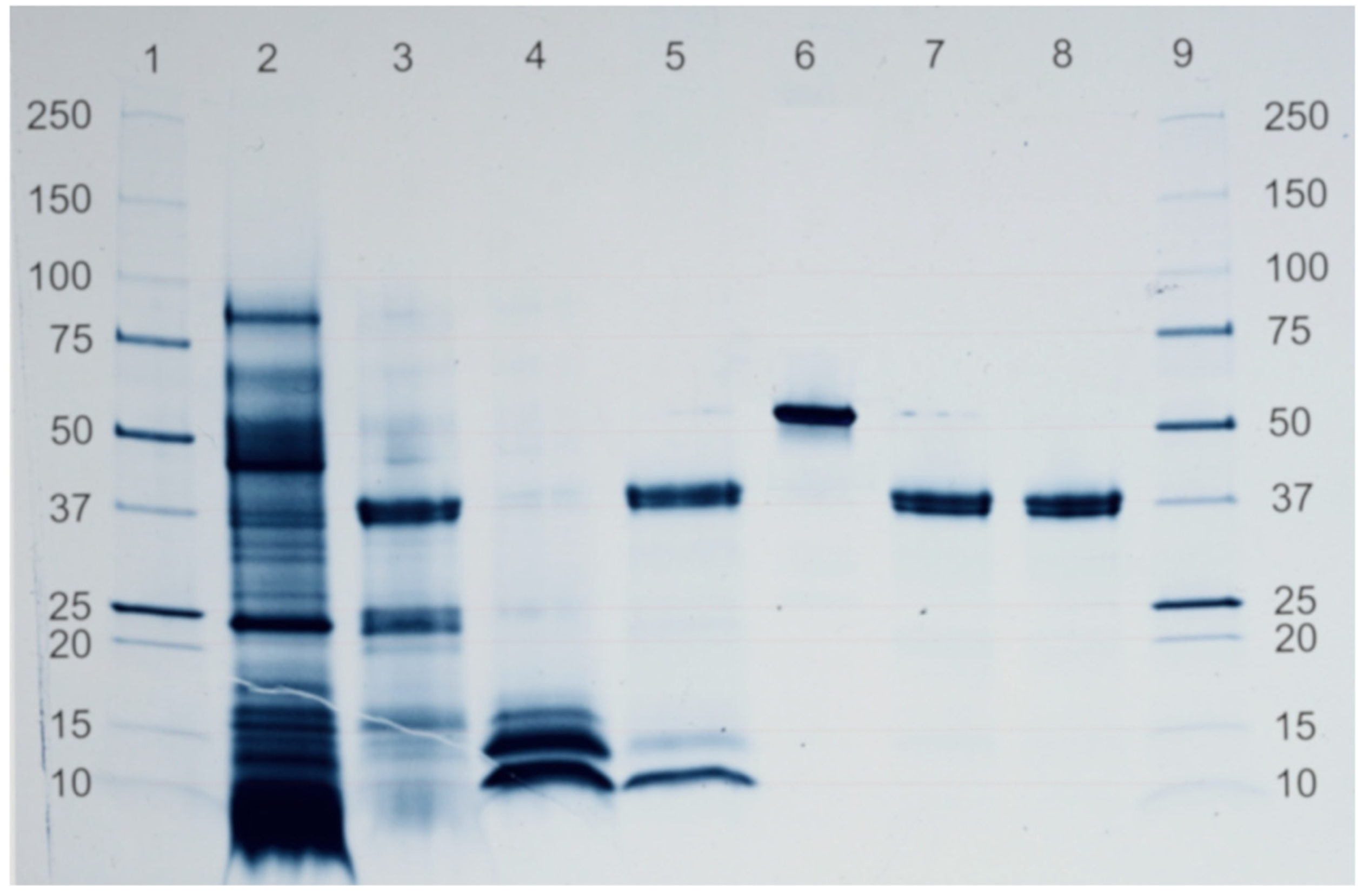
3.5. Electrophoretic Protein Profile and Digestibility
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Chatterjee, S.; Biswas, G.; Basu, S.K.; Acharya, K. Antineoplastic effect of mushrooms: A review. Aust. J. Crop. Sci. 2011, 5, 904–911. [Google Scholar]
- Ahmad, M.F. Ganoderma lucidum: Persuasive biologically active constituents and their health endorsement. Biomed Pharm. 2018, 107, 507–519. [Google Scholar] [CrossRef]
- Kong, M.; Yao, Y.; Zhang, H. Antitumor activity of enzymatically hydrolyzed Ganoderma lucidum polysaccharide on U14 cervical carcinoma-bearing mice. Int. J. Immunopathol. Pharmacol. 2019, 33, 205873841986948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharadwaj, S. Discovery of Ganoderma lucidum triterpenoids as potential inhibitors against Dengue virus NS2B-NS3 protease. Prep. Biochem. Biotechnol. 2019, 9, 19059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Marquez, E.; Lagunas-Martinez, A.; Bermudez-Morales, V.H.; Burgete-Garcia, A.I.; Leon-Rivera, I.; Montiel-Arcos, E.; Garcia-Villa, E.; Gariglio, P.; Madrid-Marina, V.V.; Ondarza-Vidaurreta, R.N. Inhibitory activity of Lingzhi or Reishi medicinal mushroom, Ganoderma lucidum (higher Basidiomycetes) on transformed cells by human papillomavirus. Int. J. Med. Mushrooms 2014, 16, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Lai, G.; Guo, Y.; Chen, D.; Tang, X.; Shuai, O.; Yong, T.; Wang, D.; Xiao, C.; Zhou, G.; Xie, Y.; et al. Alcohol Extracts From Ganoderma lucidum Delay the Progress of Alzheimer’s Disease by Regulating DNA Methylation in Rodents. Front. Farmacol. 2019, 10, 272. [Google Scholar] [CrossRef] [PubMed]
- Sargowo, D.; Ovianti, N.; Susilowati, E.; Ubaidillah, N.; Nugraha, A.W.; Proboretno, K.S.; Failasufi, M.; Ramadhan, F.; Wulandari, H.; Waranugraha, Y.; et al. The role of polysaccharide peptide of Ganoderma lucidum as a potent antioxidant against atherosclerosis in high risk and stable angina patients. Indian Heart J. 2018, 70, 608–614. [Google Scholar] [CrossRef]
- Meng, J.; Yang, B. Protective Effect of Ganoderma (Lingzhi) on Cardiovascular System. Adv. Exp. Med. Biol. 2019, 1182, 181–199. [Google Scholar] [PubMed]
- Zhao, C.; Fan, J.; Liu, Y.; Guo, W.; Cao, H.; Xiao, J.; Wang, Y.; Liu, B. Hepatoprotective activity of Ganoderma lucidum triterpenoids in alcohol-induced liver injury in mice, an iTRAQ-based proteomic analysis. Food Chem. 2019, 271, 148–156. [Google Scholar] [CrossRef]
- Ćirić, A.; Kruljević, I.; Stojković, D.; Fernandes, Â.; Barros, L.; Calhelha, R.C.; Ferreira, I.; Soković, M.; Glamočlija, J. Comparative investigation on edible mushrooms Macrolepiota mastoidea, M. rhacodes and M. procera: Functional foods with diverse biological activities. Food Funct. 2019, 10, 7678–7686. [Google Scholar] [CrossRef]
- Carocho, M.; Ferreira, I.; Morales, P.; Soković, M. Antioxidants and Prooxidants: Effects on Health and Aging. Oxid. Med. Cell Longev. 2018, 2018, 1472708. [Google Scholar] [CrossRef]
- Ferreira, I.C.; Heleno, S.A.; Reis, F.S.; Stojkovic, D.; Queiroz, M.J.; Vasconcelos, M.H.; Sokovic, M. Chemical features of Ganoderma polysaccharides with antioxidant, antitumor and antimicrobial activities. Phytochem 2015, 114, 38–55. [Google Scholar] [CrossRef] [Green Version]
- Hennicke, F.; Cheikh-Ali, Z.; Liebisch, T.; Macia-Vicente, J.G.; Bode, H.B.; Piepenbring, M. Distinguishing commercially grown Ganoderma lucidum from Ganoderma lingzhi from Europe and East Asia on the basis of morphology, molecular phylogeny, and triterpenic acid profiles. Phytochemistry 2016, 127, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Osuji, C.; Abubakar, S.; Mowobi, G. A review of pharmacological significance and need for proper authentication of Ganoderma lucidum. J. Sci. Tech. Advs. 2016, 1, 1–7. [Google Scholar]
- Amen, Y.M.; Zhu, Q.; Tran, H.B.; Afifi, M.S.; Halim, A.F.; Ashour, A.; Mira, A.; Shimizu, K. Lucidumol C, a new cytotoxic lanostanoid triterpene from Ganoderma lingzhi against human cancer cells. J. Nat. Med. 2016, 70, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Andoh, T.; Zhang, Q.; Yamamoto, T.; Tayama, M.; Hattori, M.; Tanaka, K.; Kuraishi, Y. Inhibitory effects of the methanol extract of Ganoderma lucidum on mosquito allergy-induced itch-associated responses in mice. J. Pharmacol. Sci. 2010, 114, 292–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curvetto, N.R.; Figlas, D.; Devalis, R.; Delmastro, S. Growth and productivity of different Pleurotus ostreatus strains on sunflower seed hulls supplemented with N-NH4+ and/or Mn(II). Bioresour. Technol. 2002, 84, 171–176. [Google Scholar] [CrossRef]
- Fan, C.Y.; Chien, Y.H.; Han, M.C. Use of stillage grain from a rice-spirit distillery in the solid state fermentation of Ganoderma lucidum. Process Biochem. 2003, 39, 21–26. [Google Scholar]
- Crisan, E.V.; Sands, A. Nutritional value. In The Biology and Cultivation of Edible Mushrooms; Chang, S.T., Hayes, W.A., Eds.; Academic Press, Inc.: New York, NY, USA, 1978; pp. 137–168. [Google Scholar]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E. Novel Total Antioxidant Capacity Index for Dietary Polyphenols and Vitamins C and E, Using Their Cupric Ion Reducing Capability in the Presence of Neocuproine: CUPRAC Method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Jimenez, P.; Tejero, J.; Cabrero, P.; Cordoba-Diaz, D.; Girbes, T. Differential sensitivity of D-galactose-binding lectins from fruits of dwarf elder (Sambucus ebulus L.) to a simulated gastric fluid. Food Chem. 2013, 136, 794–802. [Google Scholar] [CrossRef]
- Ozcariz, M.V. Aprovechamiento de Residuos Agroforestales, con Interés en los Originados en Explotaciones de Nuez Pecán (Caryaillinoinensis), por Medio del Cultivo de Hongos Saprófitos Saludables Ibéricos. Ph.D. Thesis, University of Valladolid, Valladolid, Spain, 2016. [Google Scholar]
- Jeurink, P.V.; Noguera, C.L.; Savelkoul, H.F.; Wichers, H.J. Immunomodulatory capacity of fungal proteins on the cytokine production of human peripheral blood mononuclear cells. Int. Immunopharmacol. 2008, 8, 1124–1131. [Google Scholar] [CrossRef]
- Kumakura, K.; Hori, C.; Matsuoka, H.; Igarashi, K.; Samejima, M. Protein components of water extracts from fruiting bodies of the reishi mushroom Ganoderma lucidum contribute to the production of functional molecules. J. Sci. Food Agric. 2018, 99, 529–535. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, S.H.; Dai, Y.C. Species clarification of the prize medicinal Ganoderma Mushroom Lingzhi. Fungal. Diversity 2012, 56, 49–62. [Google Scholar] [CrossRef]
- Ou, C.C.; Hsiao, Y.M.; Wang, W.H.; Ko, J.L.; Lin, M.Y. Stability of fungal immunomodulatory protein, FIP-gts and FIP-fve, in IFN-γ production. Food Agric. Immunol. 2009, 20, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Bastiaan-Net, S.; Chanput, W.; Hertz, A.; Zwittink, R.D.; Mes, J.J.; Wichers, H.J. Biochemical and functional characterization of recombinant fungal immunomodulatory proteins (rFIPs). Int. Immunopharmacol. 2013, 15, 167–175. [Google Scholar] [CrossRef]
- Tanaka, S.; Jeurink, K.; Tsuchiya, K.; Yamashita, A.; Murasugi, A.; Sakuma, S.; Tsunoo, H. Complete amino acid sequence of an immunomodulatory protein, Ling Zhi-8 (LZ-8): An immunomodulator from a fungus, Ganoderma lucidium, having similarity to immunoglobulin variable regions. J. Biol. Chem. 1989, 264, 16372–16377. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, T.; Tang, H.; Ren, Z.; Li, Q.; Jia, J.; Chen, H.; Fu, J.; Ding, S.; Hao, Q.; et al. Ganoderma lucidum Immune Modulator Protein rLZ-8 Could Prevent and Reverse Bone Loss in Glucocorticoids-Induced Osteoporosis Rat Model. Front. Pharmacol. 2020, 11, 731. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.W.; Guo, X.Y.; Zhang, X.C.; Guo, L.Q.; Lin, J.F. Optimization of Cultivation Conditions of Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (Agaricomycetes) for the Highest Antioxidant Activity and Antioxidant Content. Int. J. Med. Mushrooms 2019, 21, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Li, J.; Deng, K.; Ai, L. Effects of drying methods on the antioxidant activities of polysaccharides extracted from Ganoderma lucidum. Carbohydr. Polym. 2012, 87, 1849–1854. [Google Scholar] [CrossRef]
- Stojkovic, D.S.; Barros, L.; Calhelha, R.C.; Glamoclija, J.; Ciric, A.; van Griensven, L.J.; Sokovic, M.; Ferreira, I.C. A detailed comparative study between chemical and bioactive properties of Ganoderma lucidum from different origins. Int. J. Food Sci. Nutr. 2014, 65, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant properties of phenolic compounds occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Gasecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P.; Kozak, L. Phenolic and flavonoid content in Hericium erinaceus, Ganoderma lucidum, and Agrocybe aegerita under selenium addition. Acta Aliment. 2016, 45, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, K.; Riazipour, M. Ganoderma lucidum induces the expression of CD40/CD86 on peripheral blood monocytes. Iran. J. Immunol. 2009, 6, 87–91. [Google Scholar] [PubMed]
- Da, J.; Cheng, C.R.; Yao, S.; Long, H.L.; Wang, Y.H.; Khan, I.A.; Li, Y.F.; Wang, Q.R.; Cai, L.Y.; Jiang, B.H.; et al. A reproducible analytical system based on the multi-component analysis of triterpene acids in Ganoderma lucidum. Phytochemistry 2015, 114, 146–154. [Google Scholar] [CrossRef]
- Delzenne, N.M.; Bindels, L.B. Gut microbiota: Ganoderma lucidum, a new prebiotic agent to treat obesity? Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 553–554. [Google Scholar] [CrossRef]
- González-Matute, R.; Figlas, D.; Curvetto, N. Agaricus blazei production on non-composted substrates based on sunflower seed hulls and spent oyster mushroom substrate. World J. Microbiol. Biotechnol. 2011, 27, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Ogbe, A.O.; Obeka, A.D. Proximate, Mineral and Anti-Nutrient Composition of Wild Ganoderma lucidum: Implication on Its Utilization in Poultry Production. Iran. J. Appl. Animl. Sci. 2013, 3, 161–166. [Google Scholar]
- Lv, G.P.; Zhao, J.; Duan, J.A.; Tang, Y.P.; Li, S.P. Comparison of sterols and fatty acids in two species of Ganoderma. Chem. Cent. J. 2012, 6, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavishree, S.; Hemavathy, J.; Lokesh, B.R.; Shashirekha, M.N.; Rajarathnam, S. Fat and fatty acids of Indian edible mushrooms. Food Chem. 2008, 106, 597–602. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | Strain | Substrate | Colonization | First Harvest | Total Time | ||
|---|---|---|---|---|---|---|---|
| Formulation | Container | (cc) | (Days) | (Days) | (Days) | ||
| 1 | GLA | 20%T80%Ch | Jar | 200 | 28.0 ± 2.4 a | 100.75 ± 2.2 a | 128.7 ± 4.7 a |
| 1 | GLM | 20%T80%Ch | Jar | 200 | 26.7 ± 3.5 a | 99.5 ± 4.3 a | 126.5 ± 7.8 a |
| 1 | GLP | 20%T80%Ch | Jar | 200 | 35.6 ± 8.1 a | 106.0 ± 7.0 a | 141 ± 15 a |
| 2 | GLM | 20%C80%Qp | Bag | 4000 | 18.7 ± 2.3 | 79.7 ± 9.8 | 107.7± 19.6 |
| Analysis | G. lingzhi | G. lucidum |
|---|---|---|
| Dietary fibre (%) | 76.81 ± 3.46 a | 69.35 ± 3.12 b |
| Protein × 4.381 (%) | 7.47 ± 0.22 a | 11.70 ± 0.35 b |
| Carbohydrates (%) | 9.88 ± 1.04 a | 11.02 ± 1.16 a |
| Total sugars (%) | <1.00 * | <1.00 * |
| Ash (%) | 1.21 ± 0.06 a | 2.31 ± 0.12 b |
| Total Gross Fat (%) | 1.44 ± 0.10 a | 1.26 ± 0.09 a |
| Sodium (mg × 10−2 g) | 0.0050 ± 0.0006 a | 0.0026 ± 0.0003 b |
| Fatty Acid | G. lingzhi ±15% | G. lucidum ±15% |
|---|---|---|
| Capric acid | <0.05 * a | 0.09 b |
| Behenic acid | 0.4 a | 0.11 b |
| Erucic acid | <0.05 * a | 0.10 b |
| Arachidic Acid | 0.26 a | 0.15 b |
| Eicosatrienoic acid (11c,14c,17c) | <0.05 * a | 0.14 b |
| Margaric acid | 0.29 a | 0.19 b |
| Margaroleic acid | 0.52 a | 0.26 b |
| Octadecadienoic acid (9c, 12c) | <0.05 * a | 0.09 b |
| Octadecanoic/Stearic Acid | 1.3 a | 2.17 b |
| Octadecatrienoic acid (9c, 12c, 15c)/a-Linolenic acid | 0.1 a | 0.46 b |
| Octadecatrienoic acid (9t, 12c, 15t) | <0.05 * a | 0.09 b |
| Octadecenoic acid (11c)/cis-Vaccenic acid | 1.8 a | 1.24 b |
| Octadecenoic acid (9t)/Elaidic acid | <0.05 * a | 0.12 b |
| Tetracosanoic/Lignoceric Acid | 0.69 a | 0.21 b |
| Tetracosenoic acid (15c)/Nervonic acid | <0.05 * a | 0.14 b |
| Tetradecanoic Acid/Myristic | 0.21 a | 0.54 b |
| Trans fatty acids | 0.1 a | 0.31 b |
| Total Monounsaturated Fatty Acids | 37.5 a | 28.68 a |
| Total polyunsaturated fatty acids | 43.84 a | 49.93 a |
| Total saturated fatty acids | 18.64 a | 20.77 a |
| Author | [33] | [33] | [40] | [39] | Present Study | |
|---|---|---|---|---|---|---|
| Variable | ||||||
| Origin | Serbia ** | China * | Nigeria ** | NR * | Spain * | |
| Dietary Fibre (%) | NR | NR | 59.2% ± 0.9 | 66.8 | 69.4 ± 3.1 | |
| Proteins (%) | 11.3 ± 1.2 | 9.9 ± 0.3 | 17.3 ± 0.1 | 15.1 | 11.7 ± 0.4 | |
| Total Carbohydrates (%) | 81.5 ± 1.1 | 78.2 ± 0.2 | 65.8 ± 0.2 | 71.0 | 11.0 ± 1.2 | |
| Ash (%) | 2.8 ± 0.0 | 8.2 ± 0.1 | 8.7± 0.1 | NR | 2.3 ± 0.1 | |
| Total Gross Fat (%) | 4.4 ± 0.0 | 3.7 ± 0.0 | 1.6 ± 0.1 | 3.5 | 1.3 ± 0.1 | |
| Saturated FA (% T FA−1) | 15.7 ± 0.0 | 32.4 ± 0.2 | NR | 13.7 | 20.8 ± 3.1 | |
| Monounsaturated FA (% T FA−1) | 49.6 ± 0.0 | 25.2 ± 0.1 | NR | 60.9 | 28.7 ± 4.3 | |
| Polyunsaturated FA (% T FA−1) | 34.7 ± 0.0 | 42.4 ± 0.1 | NR | 25.4 | 49.9 ± 7.5 | |
| Sodium (mg × 10−2 g) | NR | NR | 236.5 ± 0.0 | NR | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraile-Fabero, R.; Ozcariz-Fermoselle, M.V.; Oria-de-Rueda-Salgueiro, J.A.; Garcia-Recio, V.; Cordoba-Diaz, D.; del P. Jiménez-López, M.; Girbés-Juan, T. Differences in Antioxidants, Polyphenols, Protein Digestibility and Nutritional Profile between Ganoderma lingzhi from Industrial Crops in Asia and Ganoderma lucidum from Cultivation and Iberian Origin. Foods 2021, 10, 1750. https://doi.org/10.3390/foods10081750
Fraile-Fabero R, Ozcariz-Fermoselle MV, Oria-de-Rueda-Salgueiro JA, Garcia-Recio V, Cordoba-Diaz D, del P. Jiménez-López M, Girbés-Juan T. Differences in Antioxidants, Polyphenols, Protein Digestibility and Nutritional Profile between Ganoderma lingzhi from Industrial Crops in Asia and Ganoderma lucidum from Cultivation and Iberian Origin. Foods. 2021; 10(8):1750. https://doi.org/10.3390/foods10081750
Chicago/Turabian StyleFraile-Fabero, Raúl, María V. Ozcariz-Fermoselle, Juan A. Oria-de-Rueda-Salgueiro, Veronica Garcia-Recio, Damian Cordoba-Diaz, María del P. Jiménez-López, and Tomás Girbés-Juan. 2021. "Differences in Antioxidants, Polyphenols, Protein Digestibility and Nutritional Profile between Ganoderma lingzhi from Industrial Crops in Asia and Ganoderma lucidum from Cultivation and Iberian Origin" Foods 10, no. 8: 1750. https://doi.org/10.3390/foods10081750