
Improved Storage Properties and Cellular Uptake of Casticin-Loaded Nanoemulsions Stabilized by Whey Protein-Lactose Conjugate

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Materials and Chemicals
2.2. Preparation and Characterization of WPI-Lactose Maillard Conjugate
2.2.1. Preparation of WPI-Lactose Maillard Conjugate (MWPI)
2.2.2. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
2.2.3. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.3. Preparation and Characterization of MWPI-Stabilized Nanoemulsions
2.3.1. Preparation of the WPI/MWPI-Stabilized Nanoemulsion
2.3.2. Storage Stability Test
2.4. Fabrication of Casticin-Loaded Nanoemulsions
2.4.1. The Solubility of Casticin in Oil
2.4.2. Preparation of Casticin-Loaded Nanoemulsion
2.4.3. Spray Drying of the Casticin-Loaded Nanoemulsion
2.4.4. Determination of Casticin Content in Nanoemulsion
2.4.5. Stability of Casticin-Loaded Nanoemulsion in the Culture Medium
2.5. Inhibitor Effects of Casticin-Loaded Nanoemulsion on Cancer Cells
2.6. Cellular Uptake of the Casticin-Loaded Nanoemulsion
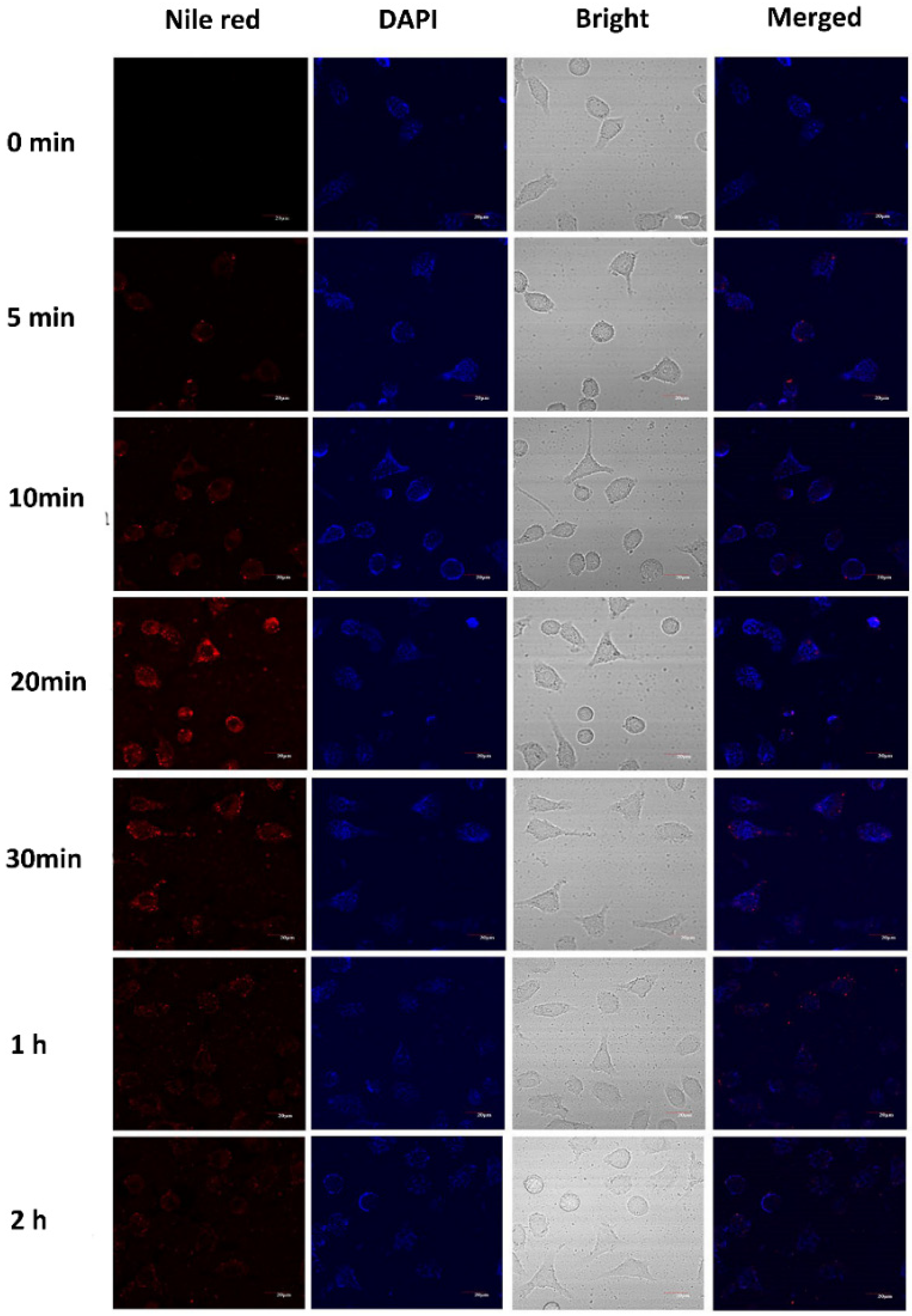
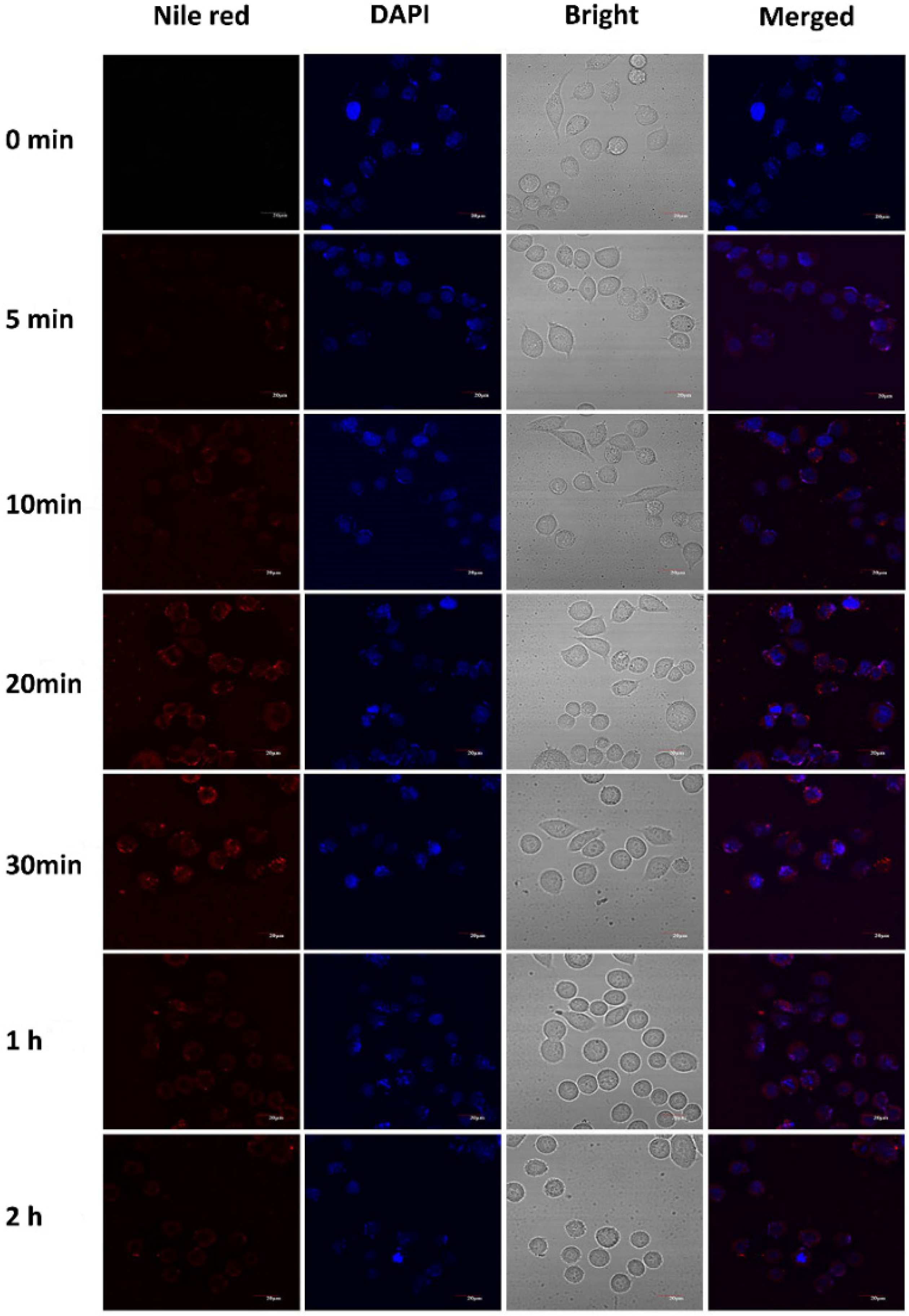
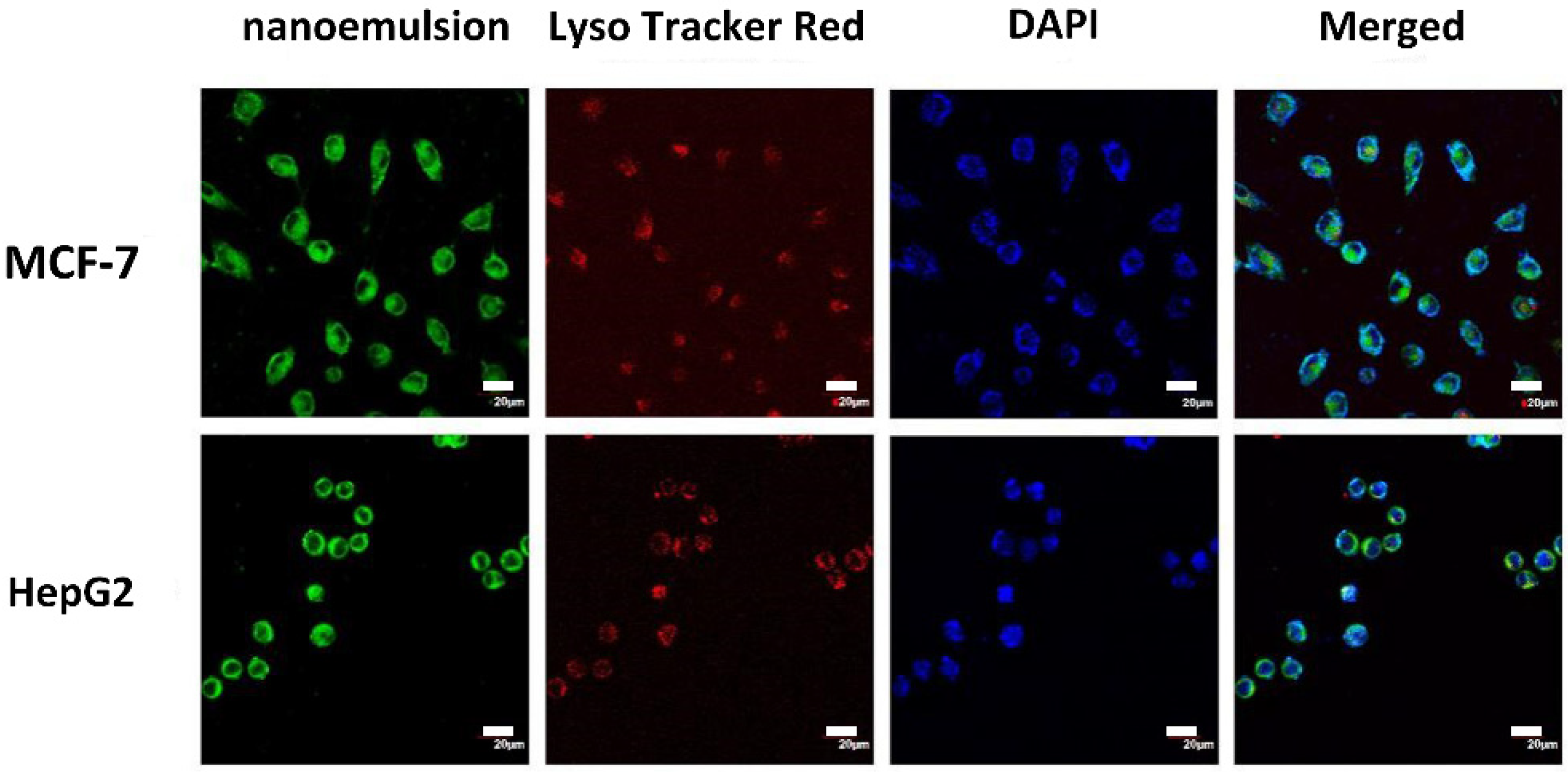
2.7. Localization of Casticin-Loaded Nanoemulsion in Cells
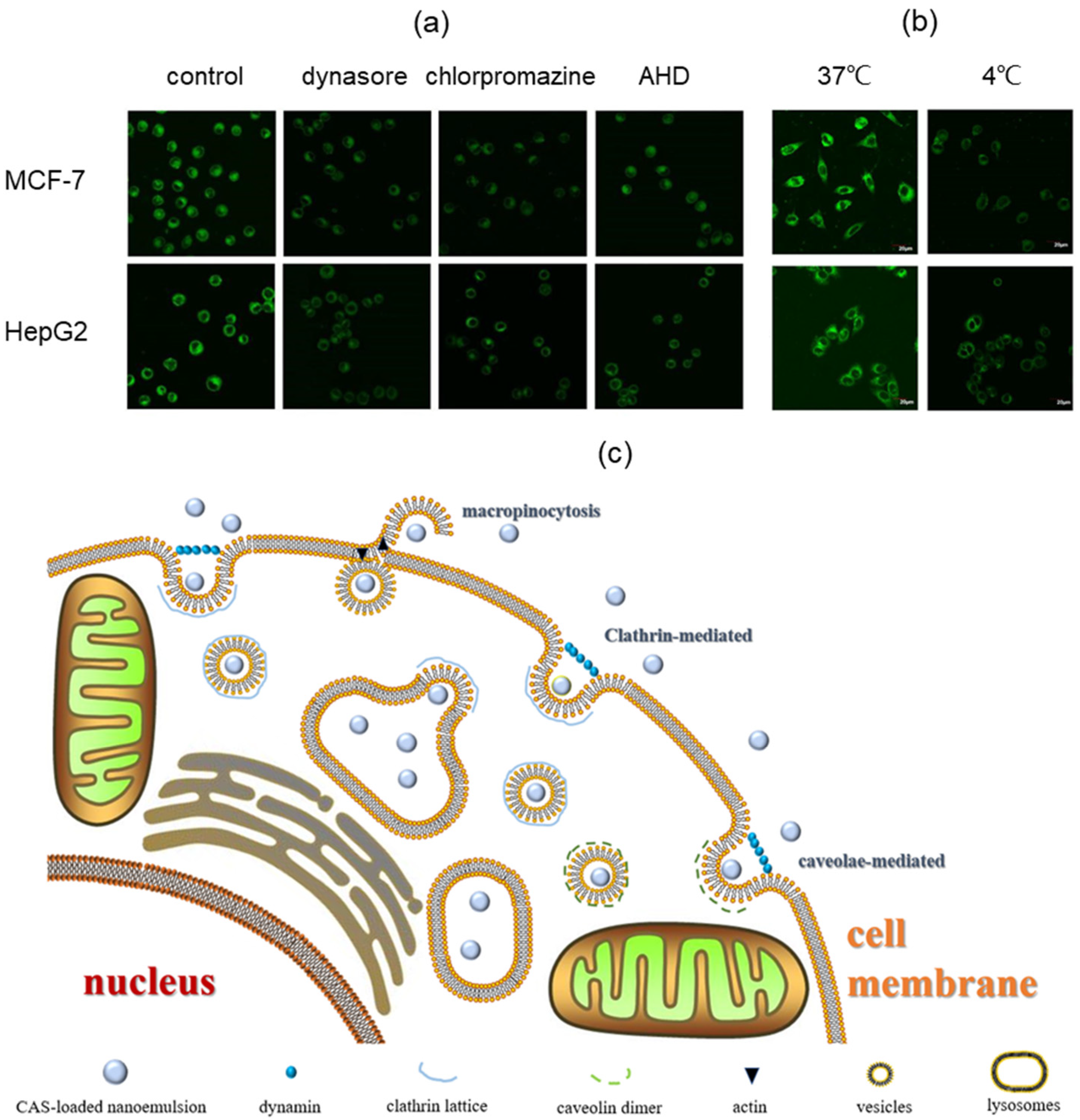
2.8. Cellular Uptake Mechanism of Casticin-Loaded Nanoemulsion
2.9. Statistical Analysis
3. Results and Discussion
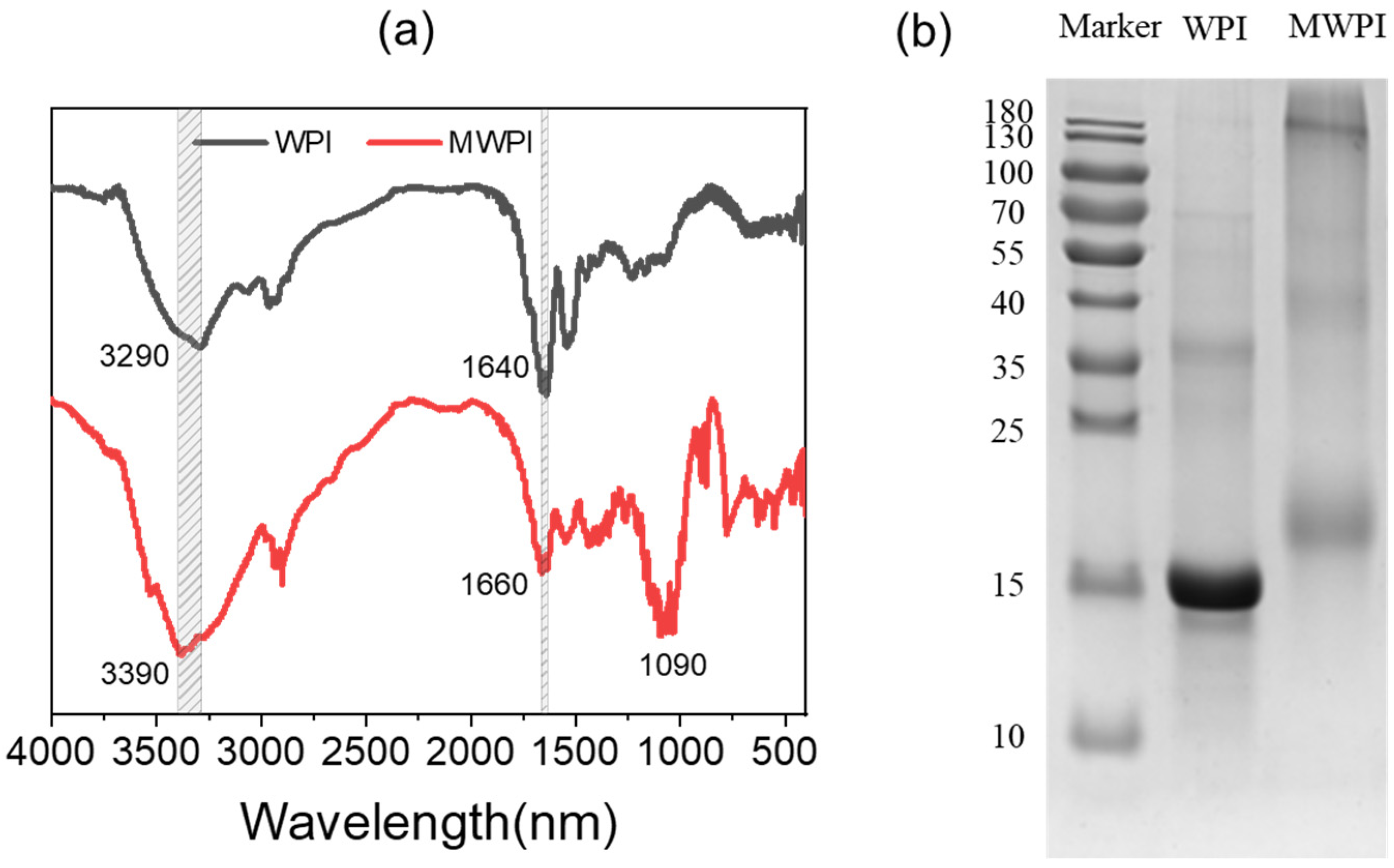
3.1. Characterization of MWPI
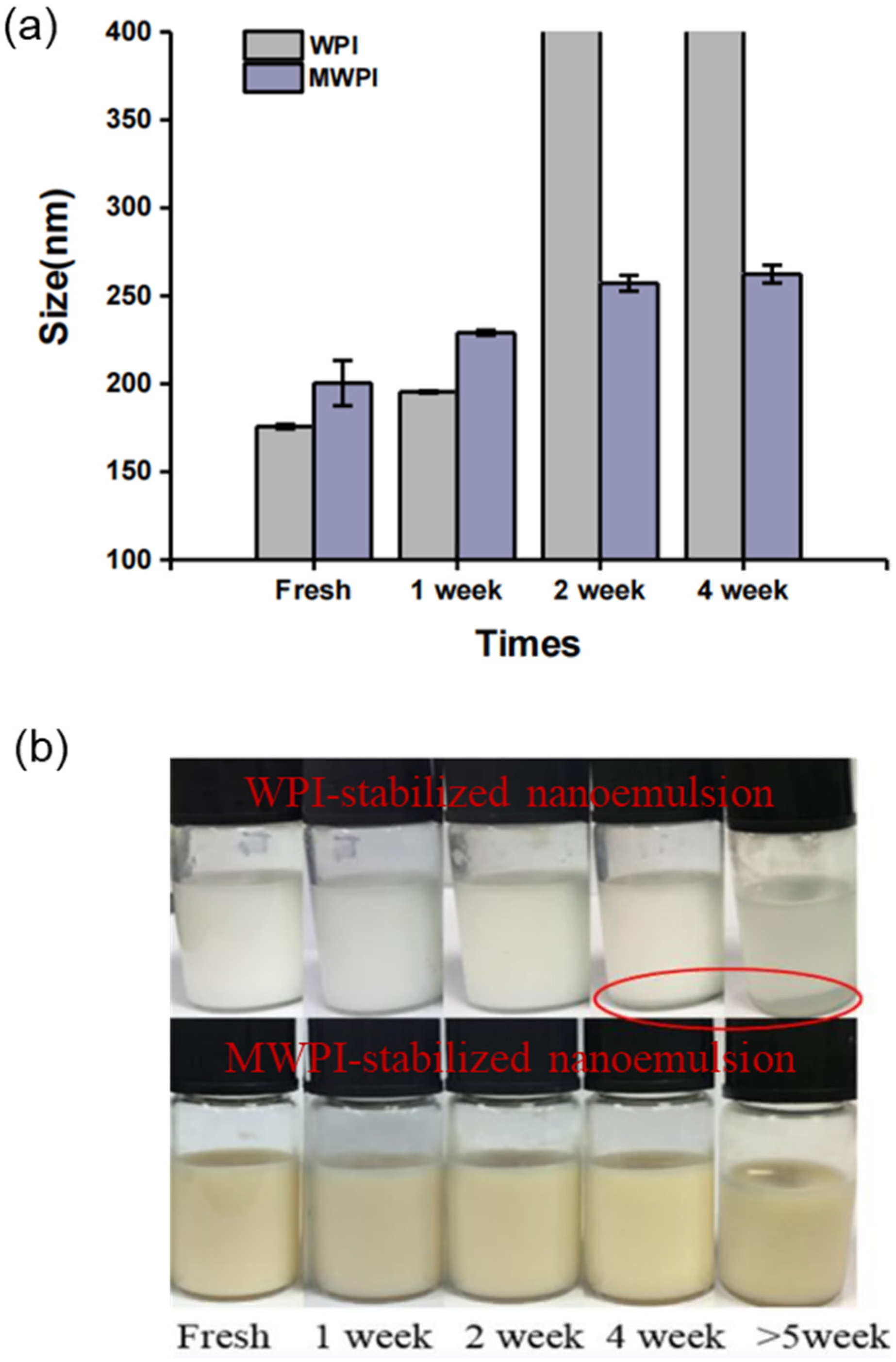
3.2. Storage Stability of WPI and MWPI-Stabilized Nanoemulsions
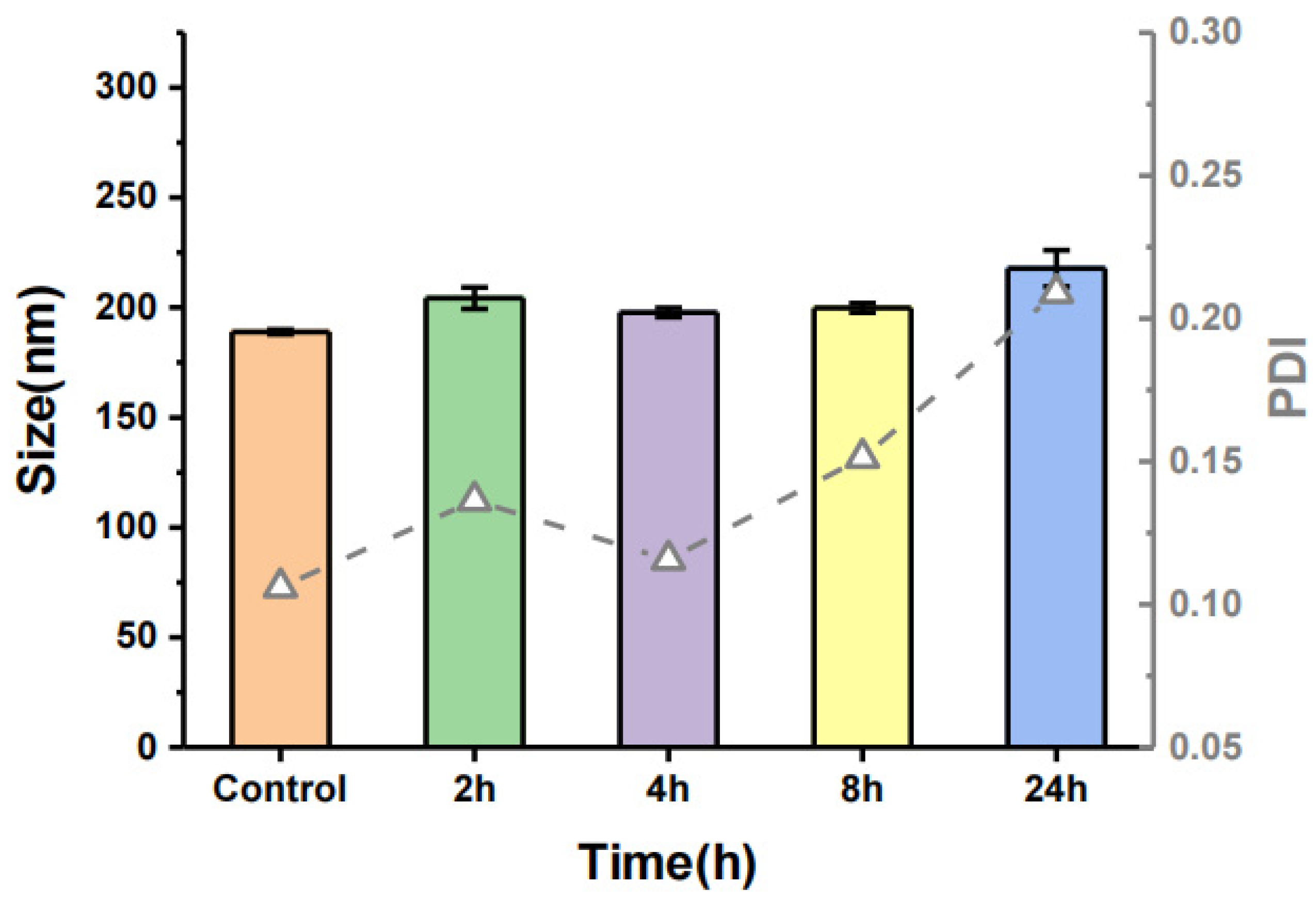
3.3. Characteristics of Casticin-Loaded Nanoemulsion
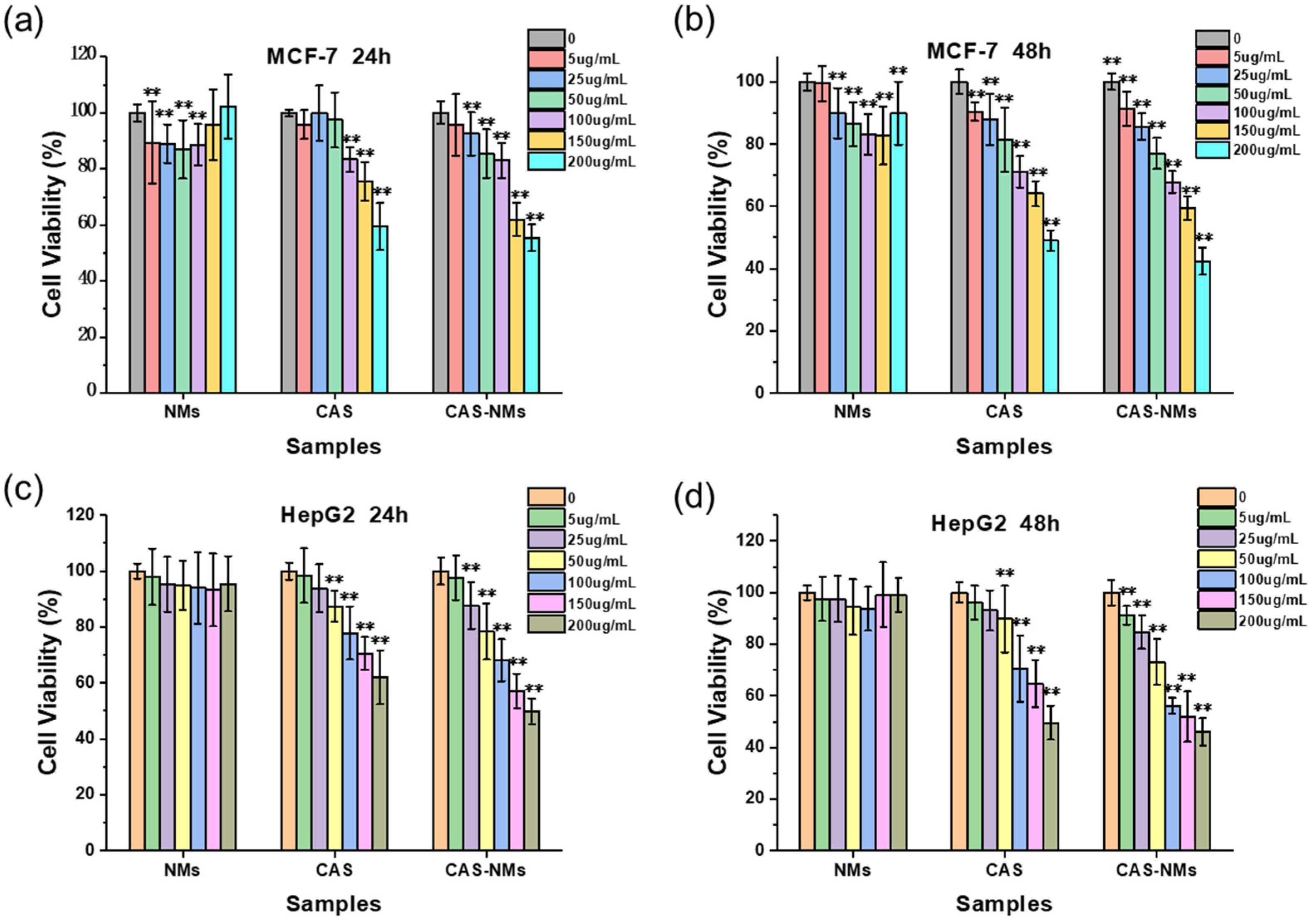
3.4. Cell Anticancer Activity of the Casticin-Loaded Nanoemulsion
3.5. Cellular Uptake of the Casticin-Loaded Nanoemulsions
3.6. Cellular Uptake Mechanism
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haidara, K.; Zamir, L.; Shi, Q.; Batist, G. The flavonoid Casticin has multiple mechanisms of tumor cytotoxicity action. Cancer Lett. 2006, 242, 180–190. [Google Scholar] [CrossRef]
- Lee, H.; Jung, K.; Lee, H.; Park, S.; Choi, W.; Bae, H. Casticin, an active compound isolated from Vitex Fructus, ameliorates the cigarette smoke-induced acute lung inflammatory response in a murine model. Int. Immunopharmacol. 2015, 28, 1097–1101. [Google Scholar] [CrossRef]
- Hu, Y.; Xin, H.L.; Zhang, Q.Y.; Zheng, H.C.; Rahman, K.; Qin, L.P. Anti-nociceptive and anti-hyperprolactinemia activities of Fructus Viticis and its effective fractions and chemical constituents. Phytomed. Int. J. Phytother. Phytopharm. 2007, 14, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Yang, J.; Yang, C.; Jiang, Y.; Zhou, Y.; Yu, B.; Yang, H. Vitexicarpin induces apoptosis in human prostate carcinoma PC-3 cells through G2/M phase arrest. Asian Pac. J. Cancer Prev. 2012, 13, 6369–6374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramchandani, S.; Naz, I.; Lee, J.H.; Khan, M.R.; Ahn, K.S. An Overview of the Potential Antineoplastic Effects of Casticin. Molecules 2020, 25, 1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, E.; Wong, S.K.; Chan, H.T. Casticin from Vitex species: A short review on its anticancer and anti-inflammatory properties. J. Integr. Med. 2018, 16, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Fani, A.; Soliva-Fortuny, R.; Martín-Belloso, O. Nanoemulsions as edible coatings. Curr. Opin. Food Sci. 2017, 15, 43–49. [Google Scholar] [CrossRef]
- Mcclements, D.J.; Rao, J. Food-Grade Nanoemulsions: Formulation, Fabrication, Properties, Performance, Biological Fate, and Potential Toxicity. Crit. Rev. Food Technol. 2011, 22, 285–330. [Google Scholar] [CrossRef]
- Gao, Y.; Xie, J.; Chen, H.; Gu, S.; Zhao, R.; Shao, J.; Jia, L. Nanotechnology-based intelligent drug design for cancer metastasis treatment. Biotechnol. Adv. 2014, 32, 761–777. [Google Scholar] [CrossRef]
- Huang, Y.; He, L.; Liu, W.; Fan, C.; Zheng, W.; Wong, Y.S.; Chen, T. Selective cellular uptake and induction of apoptosis of cancer-targeted selenium nanoparticles. Biomaterials 2013, 34, 7106–7116. [Google Scholar] [CrossRef]
- Wei, Z.; Yang, W.; Fan, R.; Yuan, F.; Gao, Y. Evaluation of structural and functional properties of protein–EGCG complexes and their ability of stabilizing a model β-carotene emulsion. Food Hydrocoll. 2015, 45, 337–350. [Google Scholar] [CrossRef]
- Cui, Z.; Kong, X.; Chen, Y.; Zhang, C.; Hua, Y. Effects of rutin incorporation on the physical and oxidative stability of soy protein-stabilized emulsions. Food Hydrocoll. 2014, 41, 1–9. [Google Scholar] [CrossRef]
- Lei, L.; Zhang, Y.; He, L.; Wu, S.; Li, B.; Li, Y. Fabrication of nanoemulsion-filled alginate hydrogel to control the digestion behavior of hydrophobic nobiletin. LWT Food Sci. Technol. 2017, 82, 260–267. [Google Scholar] [CrossRef]
- Wan, J.; Li, D.; Song, R.; Shah, B.R.; Li, B.; Li, Y. Enhancement of physical stability and bioaccessibility of tangeretin by soy protein isolate addition. Food Chem. 2017, 221, 760. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, W.; Qin, X.; Zhong, Q. Pickering emulsions stabilized by amphiphilic anisotropic nanofibrils of glycated whey proteins. Food Hydrocoll. 2019, 101, 105503. [Google Scholar] [CrossRef]
- Mengíbar, M.; Miralles, B.; Heras, Á. Use of soluble chitosans in Maillard reaction products with β-lactoglobulin. Emulsifying and antioxidant properties. LWT Food Sci. Technol. 2017, 75, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Nagaraju, P.G.; Sindhu, P.; Dubey, T.; Chinnathambi, S.; Priyadarshini, C.G.P.; Rao, J.P. Influence of sodium caseinate, maltodextrin, pectin and their Maillard conjugate on the stability, in vitro release, anti-oxidant property and cell viability of eugenol-olive oil nanoemulsions. Int. J. Biol. Macromol. 2021, 183, 158–170. [Google Scholar] [CrossRef]
- Qi, P.X.; Xiao, Y.; Wickham, E.D. Changes in physical, chemical and functional properties of whey protein isolate (WPI) and sugar beet pectin (SBP) conjugates formed by controlled dry-heating. Food Hydrocoll. 2017, 69, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.D.; Li, C.; Bin, Z.; Huang, Q.; You, L.J.; Chen, C.; Fu, X.; Liu, R.H. Physicochemical properties and bioactivity of whey protein isolate-inulin conjugates obtained by Maillard reaction. Int. J. Biol. Macromol. 2020, 150, 326–335. [Google Scholar] [CrossRef]
- Xu, D.; Wang, X.; Jiang, J.; Yuan, F.; Gao, Y. Impact of whey protein-Beet pectin conjugation on the physicochemical stability of β-carotene emulsions. Food Hydrocoll. 2012, 28, 258–266. [Google Scholar] [CrossRef]
- Liu, G.; Wang, Q.; Hu, Z.; Cai, J.; Qin, X. Maillard-Reacted Whey Protein Isolates and Epigallocatechin Gallate Complex Enhance the Thermal Stability of the Pickering Emulsion Delivery of Curcumin. J. Agric. Food Chem. 2019, 67, 5212–5220. [Google Scholar] [CrossRef]
- Liu, G.; Li, W.; Qin, X.; Zhong, Q. Flexible protein nanofibrils fabricated in aqueous ethanol: Physical characteristics and properties of forming emulsions of conjugated linolenic acid. Food Hydrocoll. 2021, 114, 106573. [Google Scholar] [CrossRef]
- Huang, S.; He, J.; Cao, L.; Lin, H.; Zhong, Q. Improved Physicochemical Properties of Curcumin-Loaded Solid Lipid Nanoparticles Stabilized by Sodium Caseinate-Lactose Maillard Conjugate. J. Agric. Food Chem. 2020, 68, 7072–7081. [Google Scholar] [CrossRef]
- Jia, C.; Cao, D.; Ji, S.; Lin, W.; Muhoza, B. Whey protein isolate conjugated with xylo-oligosaccharides via maillard reaction: Characterization, antioxidant capacity, and application for lycopene microencapsulation. LWT Food Sci. Technol. 2019, 118, 108837. [Google Scholar] [CrossRef]
- Zhang, Q.; Neoh, K.G.; Xu, L.; Lu, S.; Kang, E.T.; Mahendran, R.; Chiong, E. Functionalized mesoporous silica nanoparticles with mucoadhesive and sustained drug release properties for potential bladder cancer therapy. Langmuir 2014, 30, 6151–6161. [Google Scholar] [CrossRef]
- Peng, D.; Zhang, J.; Liu, Q.; Taylor, E.W. Size effect of elemental selenium nanoparticles (Nano-Se) at supranutritional levels on selenium accumulation and glutathione S-transferase activity. J. Inorg. Biochem. 2007, 101, 1457–1463. [Google Scholar] [CrossRef] [PubMed]
- Wumaier, M.; Yao, T.; Hu, X.; Hu, Z.; Shi, S. Luminescent Ru(II)-thiols modified silver nanoparticles for lysosome targeted theranostics. Dalton Trans. 2019, 48, 10393–10397. [Google Scholar] [CrossRef]
- Kam, N.W.S.; Dai, H. Carbon Nanotubes as Intracellular Protein Transporters: Generality and Biological Functionality. J. Am. Chem. Soc. 2005, 127, 6021–6026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Li, J.; Li, Q.; Huang, Q.; Shi, J.; Yan, H.; Fan, C. Single-particle tracking and modulation of cell entry pathways of a tetrahedral DNA nanostructure in live cells. Angew. Chem. Int. Ed. Engl. 2014, 53, 7745–7750. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Mao, Z.; Gao, C. Colloidal particles for cellular uptake and delivery. J. Mater. Chem. 2009, 19, 3108–3115. [Google Scholar] [CrossRef]
- Zhang, L.W.; Yang, J.; Barron, A.R.; Monteiro-Riviere, N.A. Endocytic mechanisms and toxicity of a functionalized fullerene in human cells. Toxicol. Lett. 2009, 191, 149–157. [Google Scholar] [CrossRef]
- Hillaireau, H.; Couvreur, P. Nanocarriers’ entry into the cell: Relevance to drug delivery. Cell. Mol. Life Sci. 2009, 66, 2873–2896. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhao, Y.; Liu, Y.; Chang, X.; Chen, C.; Zhao, Y. Cellular Uptake, Intracellular Trafficking, and Cytotoxicity of Nanomaterials. Small 2011, 7, 1322–1337. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, H.; Yanagihara, K.; Araki, N.; Mukae, H.; Sakamoto, N.; Izumikawa, K.; Seki, M.; Miyazaki, Y.; Hirakata, Y.; Mizuta, Y. Single-cell observation of phagocytosis by human blood dendritic cells. Jpn. J. Infect. Dis. 2008, 61, 294–297. [Google Scholar] [CrossRef]
- Duncan, R.; Richardson, S.C.W. Endocytosis and intracellular trafficking as gateways for nanomedicine delivery: Opportunities and challenges. Mol. Pharm. 2012, 9, 2380. [Google Scholar] [CrossRef]
- Macia, E.; Ehrlich, M.; Massol, R.; Boucrot, E.; Brunner, C.; Kirchhausen, T. Dynasore, a Cell-Permeable Inhibitor of Dynamin. Dev. Cell 2006, 10, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.C.; Wu, C.Y.; Chen, C.F. Purification of Chlorpromazine-Sensitive GTPase From Rat Cerebral Cortex. Prep. Biochem. 1995, 25, 183–195. [Google Scholar] [CrossRef]
- Koivusalo, M.; Welch, C.; Hayashi, H.; Scott, C.C.; Kim, M.; Alexander, T.; Touret, N.; Grinstein, H.S. Amiloride Inhibits Macropinocytosis by Lowering Submembranous pH and Preventing Rac1 and Cdc42 Signaling. J. Cell Biol. 2010, 188, 547–563. [Google Scholar] [CrossRef] [Green Version]
- Greulich, C.; Diendorf, J.; Simon, T.; Eggeler, G.; Epple, M.; Koeller, M. Uptake and intracellular distribution of silver nanoparticles in human mesenchymal stem cells. Acta Biomater. 2011, 7, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, Y.; Ge, M.; Zhou, G.; Sun, W.; Liu, D.; Liang, X.J.; Zhang, J. A Distinct Endocytic Mechanism of Functionalized-Silica Nanoparticles in Breast Cancer Stem Cells. Sci. Rep. 2017, 7, 16236. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Lei, M.; Huang, W.; Liu, G.; Jiang, F.; Peng, D.; Huang, Q.; Jin, W. Improved Storage Properties and Cellular Uptake of Casticin-Loaded Nanoemulsions Stabilized by Whey Protein-Lactose Conjugate. Foods 2021, 10, 1640. https://doi.org/10.3390/foods10071640
Zhang B, Lei M, Huang W, Liu G, Jiang F, Peng D, Huang Q, Jin W. Improved Storage Properties and Cellular Uptake of Casticin-Loaded Nanoemulsions Stabilized by Whey Protein-Lactose Conjugate. Foods. 2021; 10(7):1640. https://doi.org/10.3390/foods10071640
Chicago/Turabian StyleZhang, Bei, Miao Lei, Wenjing Huang, Gang Liu, Fangcheng Jiang, Dengfeng Peng, Qingrong Huang, and Weiping Jin. 2021. "Improved Storage Properties and Cellular Uptake of Casticin-Loaded Nanoemulsions Stabilized by Whey Protein-Lactose Conjugate" Foods 10, no. 7: 1640. https://doi.org/10.3390/foods10071640