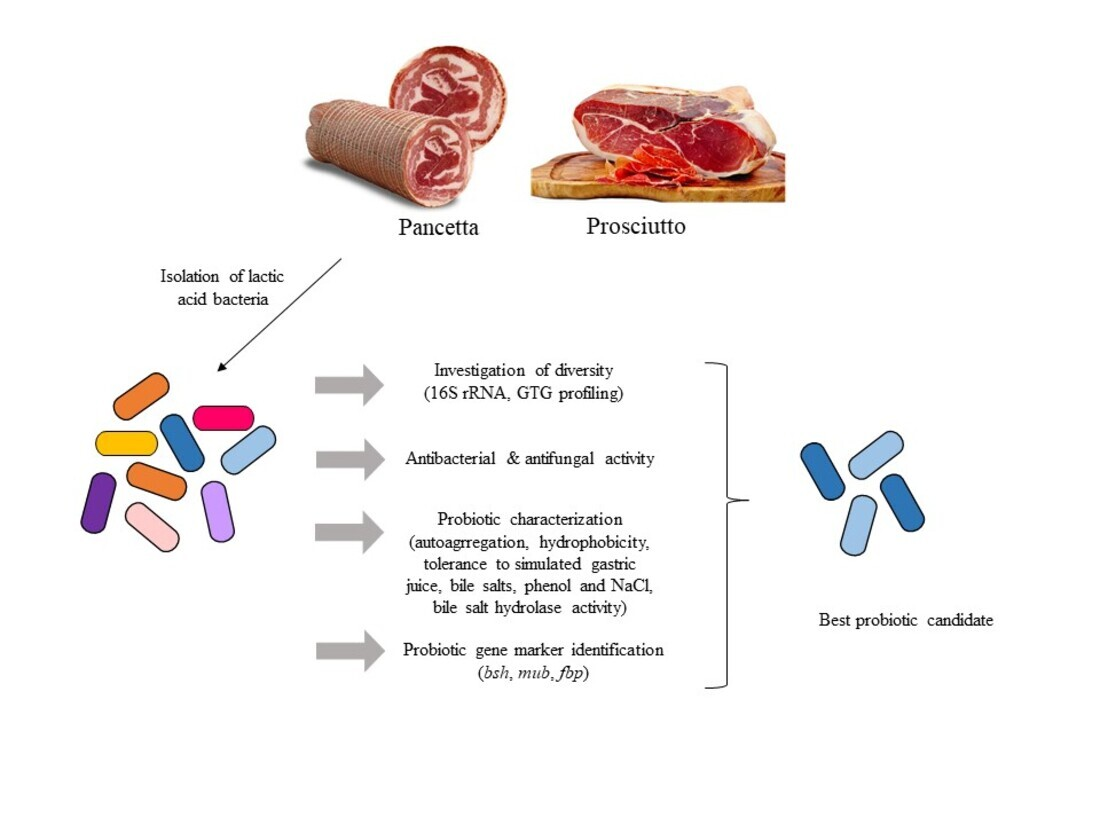
3.1. Diversity of LAB in Pancetta and Prosciutto
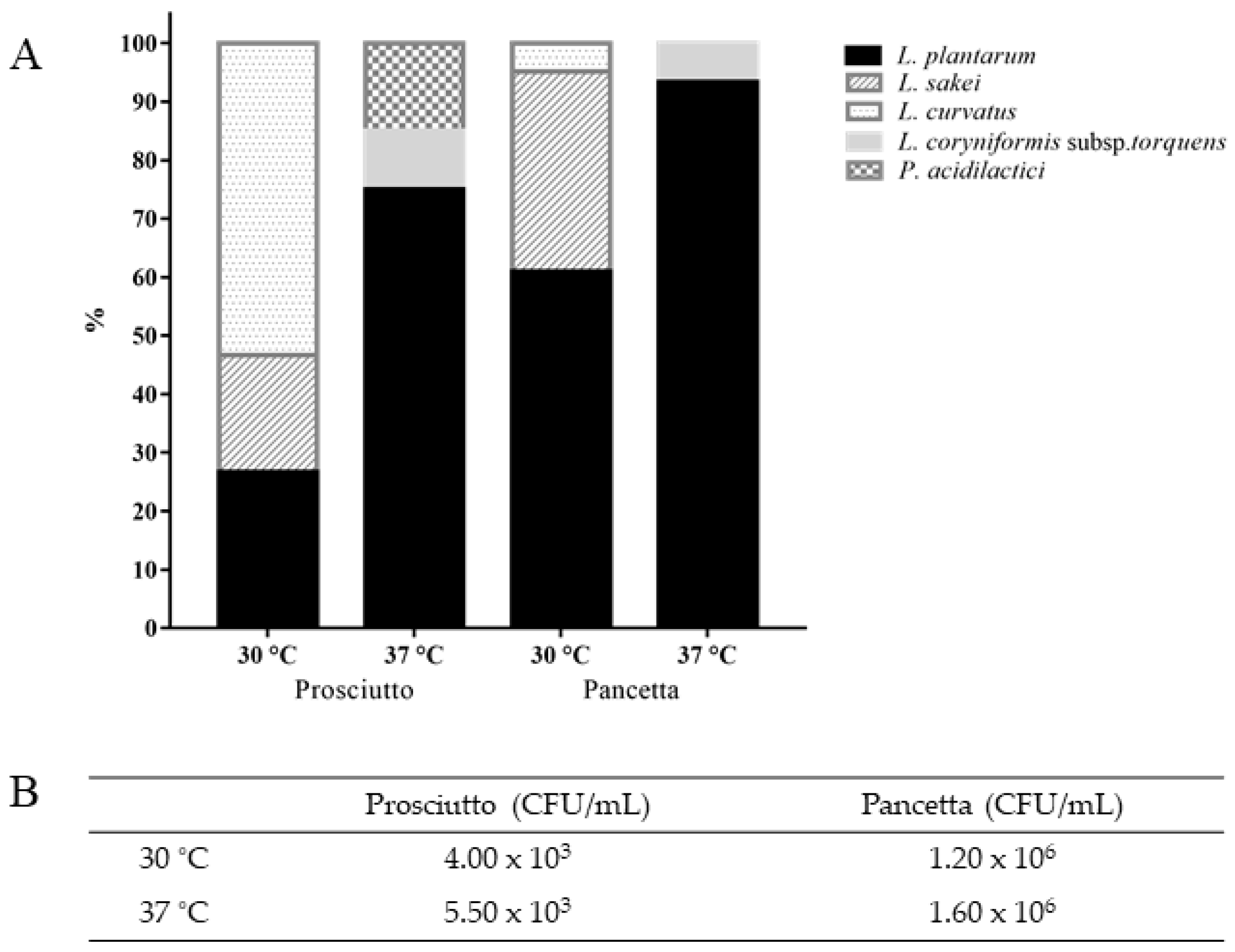
Fermented meats are reported to be a rich source of LAB and we therefore evaluated and compared the microbial composition of two Italian fermented meat products. This was achieved through a combination of classical microbiological and molecular approaches, permitting an evaluation of the diversity and distribution of LAB in prosciutto and pancetta. MRS was used as a selective medium to isolate potential probiotic LAB. Prior to the isolation of individual colonies, the number of viable colonies (CFU/mL) of both products was counted on MRS agar at 30 °C and 37 °C. The prosciutto sample was observed to host approximately 10
3 CFU/mL, while significantly higher counts were observed for pancetta at approximately 10
6 CFU/mL at both temperatures (
Figure 1B). A total of 35 isolates from prosciutto and 71 isolates from pancetta were retrieved for 16S rRNA gene sequence analysis.
Analysis of the isolates derived from prosciutto at 30 °C suggests a dominance of
L. curvatus (8/15 or 53%), in addition to
L. plantarum (4/15 or 27%) and
L. sakei (3/15 or 20%) (
Figure 1A). At an incubation temperature of 37 °C, however, the majority of the isolated LAB population was represented by
L. plantarum (15/20 or 75%) with a significantly lower percentage of
P. acidilactici (3/20 or 15%) and
Loigolactobacillus coryniformis subsp.
torquens (2/20 or 10%). In the pancetta sample, the dominant LAB species appeared to be
L. plantarum regardless of the incubation temperature (
Figure 1A). There was a comparatively higher proportion of
L. sakei (14/41 or 34%) compared to
L. curvatus (2/41 or 5%) at 30 °C while a small subpopulation of
L. coryniformis subsp.
torquens (2/30 or 7%) was identified specifically at 37 °C. Overall,
L. plantarum was the predominant LAB species in both meat products and prosciutto appeared to exhibit a higher diversity of organisms depending on the incubation temperature.
L. coryniformis subsp.
torquens and
P. acidilactici were identified specifically at 37 °C, whereas
L. sakei and
L. curvatus were determined to be more dominant at 30 °C (
Figure 1A).
L. plantarum isolates were obtained at both 30 °C and 37 °C, which likely reflects this species’ versatility and adaptability to grow at different temperatures.
Long fermentation and ripening times expose LAB to prolonged stress, including acid stress, oxidation and starvation, which in turn affects their survival in specific food niches. The accumulation of lactic acid produced by LAB could exert physiological changes [
41], which may cause the collapse of the proton motive forces as it acidifies the cytoplasm, resulting in cell death [
42].
L. plantarum is regarded as one of the most versatile and industrially important LAB due to its useful properties, a high survivability and being functional in a range of fermented food niches [
43].
L. plantarum was also observed to be among the dominant LAB species found in fermented meat products of different geographical regions, including Asia [
44,
45], South America [
46] and Africa [
47]. It is generally accepted that
L. plantarum is highly tolerant against acid and alkali stress and some strains of
L. plantarum were reported to be able to withstand heat, oxidative strees and starvation stress [
48,
49,
50].
A previous study of LAB diversity in fermented meats produced in a range of European countries highlighted a dominance of
L. sakei in a variety of fermented meat products [
51]. The second most predominant species was
P. pentosaceus (for Belgium and Germany) or
L. curvatus (for Spain, France and Italy).
P. acidilactici and
L. coryniformis subps.
torquens species have also been found in fermented meat products [
52,
53,
54,
55], although they were not commonly identified as the dominant LAB species surviving in European fermented meats. The findings in the present study concur with what has been reported in literature as
Figure 1A demonstrated low percentage of
L. coryniformis subps.
torquens (10% in prosciutto and 5% in pancetta) and
P. acidilactici (15% in prosciutto). However, it is noteworthy that the dominant LAB components identified in the present study differed from those observed in the pan-European study, suggesting a product-specific microflora.
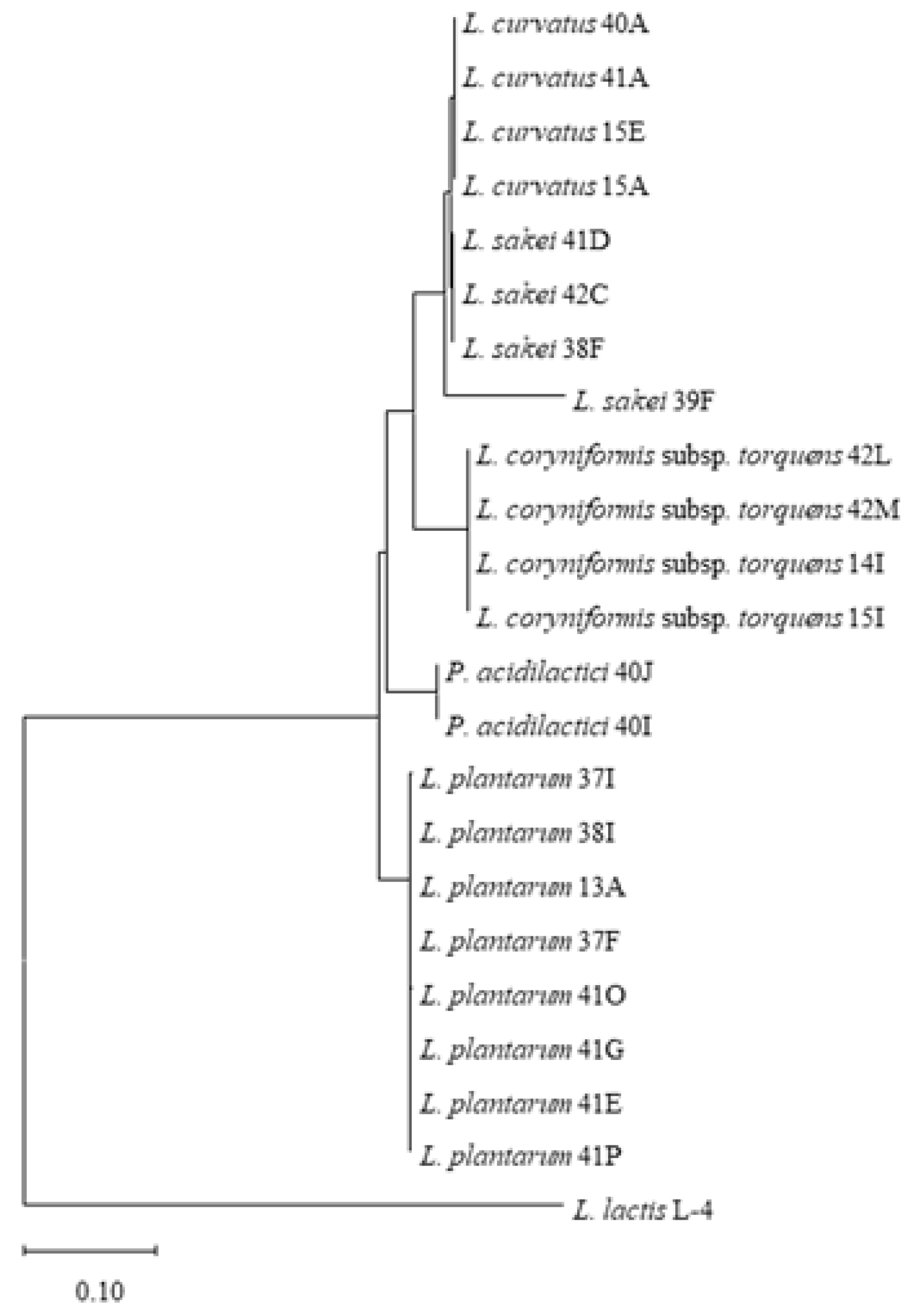
Based on the 16S rRNA gene sequence data, food source and incubation temperature (
Table S2), we selected 22 representative LAB isolates and subjected these strains to genetic fingerprinting analysis (GTG)
5 (
Figure S1), thereby yielding species level profiles which confirmed the 16S rRNA speciation results of the selected isolates. Isolates of
L. plantarum (lanes 1–8),
L. coryniformis subsp.
torquens (lanes 17–20) and
P. acidilactici (lanes 21–22) were shown to exhibit species-specific profiles. Furthermore, (GTG)
5 profiles of
L. sakei strains 38F and 39F (lanes 11–12) were shown to be highly similar while that of
L. sakei 41D was shown to lack a band (~1400 bp), whereas the
L. sakei 42C strain produced a slightly smaller band of less than 1400 bp, suggesting that these may be distinct isolates. While the genetic fingerprinting profiles of
L. curvatus 40A, 41A and 15E (lane 13, 14 and 16) were shown to be highly similar,
L. curvatus 15A exhibits an additional distinct amplicon (~5000 bp). The GTG species level profiling was consistent with phylogenetic analysis of the 16S rRNA gene sequences of these strains with four distinct clusters representing the five LAB species (
L. sakei and
L. curvatus cluster together in the phylogenetic tree) identified in this study (
Figure 2 and
Figure S1).
3.2. Organic Acid Production and Antagonistic Activity
LAB may possess a competitive advantage to survive through the production of antimicrobial compounds including organic acids. LAB may produce more than one type of organic acid (although primarily lactic acid) and this may vary by species and even strain. In order to evaluate the organic acid production profiles of 22 representative isolates, the cell-free supernatant (CFS) of fresh overnight cultures of the selected strains were analyzed by HPLC. Representative isolates of
L. plantarum,
L. sakei,
L. curvatus,
L. coryniformis subsp.
torquens and
P. acidilactici were analyzed in this manner. Perhaps unsurprisingly, the isolates were observed to primarily produce lactic and acetic acids (
Figure 3,
Table S3). Among the five different species of LAB, strains of
L. plantarum produced the highest concentration of lactic acid compared to other species, with
L. plantarum 41O and 41P producing significantly (
p < 0.001) higher lactate of 157.09 ± 0.29 mmol/L and 158.02 ± 0.22 mmol/L, respectively, compared to
L. plantarum 41G, 41E, 13A, 37F, 38I and 37I (106.67 to 138.91 mmol/L).
L. curvatus (71.14 to 78.07 mmol/L) and
L. sakei (68.75 to 81.90 mmol/L) were significantly weaker lactic acid producers compared to
P. acidilactici (~88 mM). In general,
L. plantarum species also formed significantly (
p < 0.001) higher acetate compared to other species, with
L. plantarum 41O, 41P, 37I and 38I (ranging from 45.16 to 46.77 mmol/L) being the strongest than compared to
L. plantarum 41G, 41E, 13A and 37F (ranging from 40.94 to 43.82 mmol/L). The concentration of acetate produced by
L. sakei did not significantly (
p > 0.05) differ compared to
L. coryniformis subsp.
torquens,
P. acidilactici and
L. curvatus. Furthermore, propionate was not produced by the tested isolates except for
L. plantarum at a range of 2.81 to 4.90 mmol/L.
The inhibitory activity of the selected 22 LAB isolates was assessed using the spot-on-lawn antimicrobial assay against several indicator organisms associated with food spoilage and/or food poisoning. The inhibitory activity of LAB may be due to the production of organic acids and/or bacteriocins and thus it was aimed to discern the source of antimicrobial activity where relevant. The nature of inhibitory activity was classified into two types (shown in
Figure S2): If a hazy undefined zone was observed, then the inhibitory effect was predicted to be primarily due to organic acid production, whereas if a clear defined zone was observed it was presumed to be due to bacteriocins [
56]. In summary, all
L. plantarum and
L. sakei isolates exhibited inhibitory activity against all indicators tested in the spot-on-lawn assays (
L. cremoris HP;
L. paramesenteroides NCDO869;
L. mesenteroides NCDO2028;
K. aerogenes NCIMB10102;
E. faecium NCIMB11508;
P. aeruginosa PA01;
L. innocua UCC3 and
S. aureus NCDO949) (
Table 1).
L. curvatus 40A and 41A exhibited weak inhibitory activity against
L. cremoris HP and
L. curvatus 15A and 15E presented a weak antagonistic effect against
S. aureus NCDO944 alone among the tested indicator strains.
L. coryniformis subsp.
torquens 42L and 42M demonstrated weak to moderate inhibitory activity against all indicators, whereas 14I and 15I showed similar activity against all indicators except for
L. cremoris HP and
E. aerogenes NCIMB10172 (
Table 1).
L. innocua UCC3,
E. faecium NCIMB11508 and
S. aureus NCDO944 were strongly inhibited by
P. acidilactici 40J and 40I (
Table 1).
The effectiveness of organic acids as a natural preservative in contributing to food protection and safety against a range of spoilage and pathogenic microorganisms has been well studied [
57,
58,
59]. Bacterial genera associated with food spoilage, such as
Lactococcus,
Leuconostoc,
Klebsiella and
Pseudomonas [
60,
61], and pathogenic bacteria associated with food poisoning, such as
Listeria and
Staphylococcus [
62,
63], were used as indicators in this study (
Table 1). Among the 22 strains tested, 14 isolates of
L. plantarum,
L. sakei and
P. acidilactici inhibited all indicators tested, whereas
L. curvatus and
L. coryniformis subsp.
torquens showed antagonistic ability at least against one indicator tested. Additionally, the antifungal potential of the isolates was evaluated. All isolates of
L. plantarum,
P. acidilactici and
L. coryniformis subsp.
torquens inhibited
P. expansum DSM1282 (
Table 1), but none of the isolates were active against
P. digitatum DSM2731.
P. expansum is recognized as one of the most dominant post-harvest pathogens in fruits and vegetables mainly in pomaceous fruits [
64]. While the compound(s) that exert the anti-fungal activity are not defined within the scope of this study, previous studies have reported the anti-fungal activity of phenyllactic acid produced by
L. plantarum [
65] and a bacteriocin-like compound produced by
P. pentosaceous [
66].
Findings from this study indicate that the presence of LAB in prosciutto and pancetta play a crucial role in controlling spoilage and/or pathogenic microorganisms through the production of organic acids and, in some cases, bacteriocins. However, it is important to note that organic acids and bacteriocins may not be the sole contributing factors, as LAB may also produce a variety of antimicrobial compounds including hydrogen peroxide, carboxylic acid, diacetyl and reuterin [
67].
3.3. Bacteriocin Activity and Associated Gene Clusters
In order to further validate that the inhibitory effect exerted by LAB isolates were not due to the reduced pH, well diffusion assays using pH neutralized and catalase treated (to remove H
2O
2 effect) CFS from selected representative isolates—
L. plantarum 13A, 41G, 38I, 41P and
P. acidilactici 40J (
Table 2)—were performed. These representative isolates were selected based on the product origin and the incubation temperature they were isolated from, as shown in
Table S2. These assays confirmed that
L. plantarum 13A, 41G, 38I and 41P produced bacteriocins that were effective against
L. cremoris HP, MG1363, NZ9000 and
L. paramesenteroides NCDO869, while that produced by
P. acidilactici 40J was effective against
L. mesenteroides NCDO2028,
E. faecium NCIMB11508 and
L. innocua strains UCC3, DPC3565, DPC3566 and DPC3567 (
Table 2). Growth of
K. aerogenes NCIMB10102,
P. aeruginosa PA01 and
S. aureus NCDO949 was not inhibited by the three tested isolates, which suggests that the inhibitory activity against these indicators was due to organic acids (
Table 1). In addition, the compounds produced by
L. plantarum 13A, 41G, 38I, 41P and
P. acidilactici 40J were sensitive to proteinase K, which further substantiated their proteinaceous nature.
In order to identify bacteriocin-encoding gene clusters responsible for the observed antimicrobial activity, the genomes of five isolates were sequenced and assembled. Potential bacteriocin gene clusters in these five isolates were detected based on in silico analysis of retrieved contigs with BAGEL4 and antiSMASH. Based on this analysis, it is proposed that
P. acidilactici 40J produces a pediocin-like bacteriocin with genes encoding the putative structural peptide (PedA); the immunity protein PedB; the transport proteins, PedC and PedD with 99 to 100% identity to those that encode pediocin PA1 produced by
P. acidilactici K10 (
Figure S3). Additionally, the organization of the gene clusters in
P. acidilactici 40J and K10 are identical. The bacteriocin produced by this strain (40J), which is likely pediocin, was effective against
Listeria,
Enterococcus and
Leuconostoc (
Table 2) and in agreement with the literature pertaining to pediocin PA1 [
68].
Different strains of
L. plantarum have been reported to produce bacteriocins with varied structural properties and antagonistic activities. For instance,
L. plantarum B21 was previously demonstrated to be effective against a broad range of LAB and a few non-LAB species (
Listeria monocytogenes and
Clostridium perfringens) [
69], whereas
L. plantarum LPL-1 was effective against a broad range of non-LAB species (
L. monocytogenes,
S. aureus,
E. faecalis,
B. amyloliquefaciens and
B. pumilus) and several strains of
Lactococcus and
Lactobacillus [
70].
L. plantarum NI326 produces the cyclic bacteriocin plantaricyclin A, which shared 99 to 100% similarity to the bacteriocin encoding cluster of
L. plantarum 13A, 38I and 41G isolated in the present study (
Figure S3), that is effective against some strains of
Lactococcus,
L. bulgaricus and
Alicyclobacillus acidoterrestris but is not effective against
L. plantarum,
L. brevis,
Pediococcus,
Listeria,
Escherichia,
Staphylococcus,
Streptococcus,
Klebsiella and
Bacillus [
71]. Based on comparative analysis of the identified bacteriocin encoding gene clusters, it is likely that 41P produces novel bacteriocin(s), which is potentially a derivative of plantaricyclin A since the only identifiable bacteriocin-encoding region in the strain’s genome (and which is plasmid-encoded) possessed only 42% similarity to plantaricyclin A. However, further investigations through purification and identification using mass spectrometry are required to confirm its potential novelty.
3.4. Antibiotic Susceptibility and Screening for Probiotic Candidates
Horizontal transfer of antibiotic resistance genes, particularly those within mobile genetic elements (transposons and plasmids) deserve particular attention due to the risk of resistance transfer to pathogenic organisms [
72]. The determination and comparison of antibiotic susceptibility patterns in a number of representative strains of each species are major steps in differentiating between intrinsic and acquired antibiotic resistance in probiotic bacteria [
72]. Antibiotic resistance tests were first performed as a preliminary screen for potential LAB strains amongst the 22 isolates (
Table 3). It is generally known that
Lactobacillus sp. are intrinsically resistant to gentamicin, streptomycin, nalidixic acid and vancomycin [
73,
74].
L. curvatus 40A displayed widespread antibiotic-resistance amongst the
L. curvatus strains tested, while 42C was the most sensitive amongst the
L. sakei isolates.
L. plantarum isolates 37F and 41G were the most antibiotic-sensitive; 37I, 38I and 41P were moderately sensitive, while 13A, 41O and 41E were resistant to several antibiotics. Nonetheless, 37F, 41G, 38I, 41P and 13A, which exhibited distinct sensitivity/resistance profiles, were selected for further probiotic tests since all
L. plantarum isolates were bacteriocin producers (
Table 1). Additionally,
P. acidilactici 40J,
L. coryniformis subsp.
torquens 42L, 42M and 14I were selected for further analysis.
In this study, hexane was used to evaluate hydrophobicity of LAB cell surface. This method is considered important in determining probiotic bacterial adhesion capacity to epithelial host cells [
5,
75].
L. coryniformis subsp.
torquens and
L. plantarum displayed significantly (
p < 0.05) greater % hydrophobicity (ranging from 31.2 to 42.2%) compared to
L. curvatus,
L. sakei and
P. acidilactici (ranging from 4.9 to 18.1%) (
Table 4). Isolates 42L, 14I, 41P, 41G and 38I did not differ significantly (
p > 0.05) and showed highest overall % hydrophobicity. According to the classification proposed by Colloca et al. [
76], bacterial hydrophobicity can be considered low (0 to 34%), moderate (35 to 69%) or high (70 to 100%). Altogether, the hydrophobicity values for meat isolates could be considered moderate when compared to those previously reported.
The auto-aggregation capacity of bacteria has also been associated with higher adhesion to epithelial cells and persistence in the intestine [
77]. All isolates tested in this study were shown to exhibit auto-aggregation during the first 3 h (from 9.7 to 15.6%), which increased with time after 24 h (from 28.9 to 69.0%) (
Table 4). In particular,
L. plantarum 41P, 38I, 37F and 41G did not significantly (
p > 0.05) differ and showed the highest auto-aggregation levels (63.6 ± 2.2 to 69.0 ± 4.2%, t = 24 h) compared to all other isolates. Autoaggregation % of
L. plantarum strains reported in this study was similar to several lactobacilli and
L. plantarum strains reported by other studies [
77,
78,
79]. Thus, autoaggregation levels of meat isolates in this study are generally similar or slightly higher to the probiotic candidates reported in the literature.
The abilities to tolerate gastric juice (stomach phase) and bile salt (intestinal phase) are important properties of probiotics to ensure their viability and survival in the human gastrointestinal tract [
80]. It was revealed that
L. curvatus and
L. sakei do not survive simulated gastric juice conditions after 1 h incubation and
L. coryniformis subsp.
torquens is unable to withstand such conditions following 2 h exposure (
Table 5). However, they are able to survive 0.3% bile salt exposure for a period of 3 h with approximately 5-fold to 9-fold viable count reduction (
Table 6). In contrast,
L. plantarum is able to withstand simulated gastric juice to some degree with 10-fold to 15-fold reduction after 1 h, but with declined survival (ranging from 10
4 to 10
2 CFU/mL) after 2 h and 3 h (
Table 5). These isolates appear to be more robust than previous reports on this species [
78]. Similarly,
L. plantarum strains isolated in this study were highly tolerant to bile salt stress, remainig viable at 10
8 CFU/mL (5 to 6-fold reduction) after 3 h (
Table 6). Furthermore,
P. acidilactici remained viable at 10
5 to 10
4 CFU/mL after a challenge with simulated gastric juice for 1 h to 3 h (
Table 5) and an 8-fold reduction in survival was observed after a challenge with bile salt for 3 h (
Table 6). It was reported that the median of gastric emptying half time in healthy adults is approximately 1 h [
81], but there are considerable variabilities among individuals [
82]. Therefore,
L. coryniformis subsp.
torquens could still be considered as a probiotic candidate. In summation, all isolates display high tolerance to 0.3% bile salt (displayed by less than 10-fold reduction in survival after 3 h), but only
L. plantarum and
P. acidilactici strains are able to survive more than 2 h exposure to simulated gastric juice.
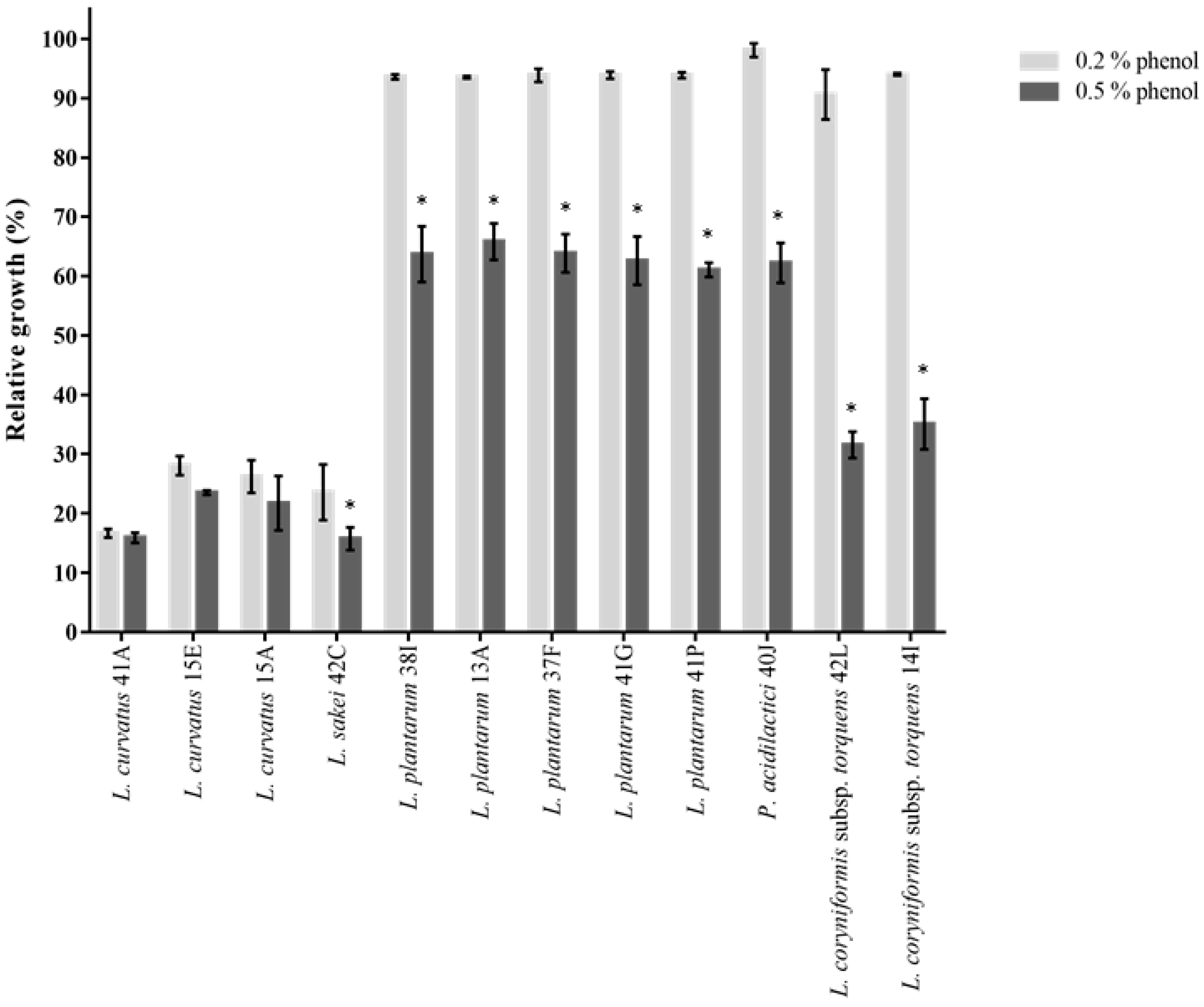
Probiotics also need to survive toxic metabolites, particularly phenols produced during the digestion process. Amino acids derived from the diet or endogenous proteins can be deaminated by bacteria in the gastrointestinal tract which results in the formation of phenols [
26]. All strains of
L. plantarum,
P. acidilactici and
L. coryniformis subsp.
torquens generally exhibited almost unimpaired growth in the presence of 0.2% phenol (>90% growth compared to untreated control) (
Figure 4). Upon exposure to 0.5% phenol, their growth was significantly (
p < 0.05) decreased from 61 to 65% (for 38I, 13A, 37F, 41G and 41P), 62% (for 40J), 31 and 35% (for 42L and 14I, respectively). These values were arguably higher compared to what was reported in other studies [
27,
78].
L. curvatus and
L. sakei were nonetheless shown to exhibit low survival to 0.2% phenol (from 16 to 28%) and 0.5% phenol (from 15 to 23%) stress (
Figure 4). It is important to note that phenol content in foods (ranging from flour, oil, raw beans, cocoa, fruits and wine) greatly varies from 0.0002 to 3.6% (
w/
w) [
83]. Furthermore, its bioavailability in the human gut greatly varies depending on several factors, such as the individual’s diet, the matrix of polyphenol-rich foods which may affect intestinal absorption and, likewise, the interaction of phenols with nutrients taken from the same meal could result in alteration in their absorption [
83]. For these reasons, 0.2% and 0.5% phenol concentrations (selected as the average representative phenol content present in foods) were tested against a panel of probiotic candidates.
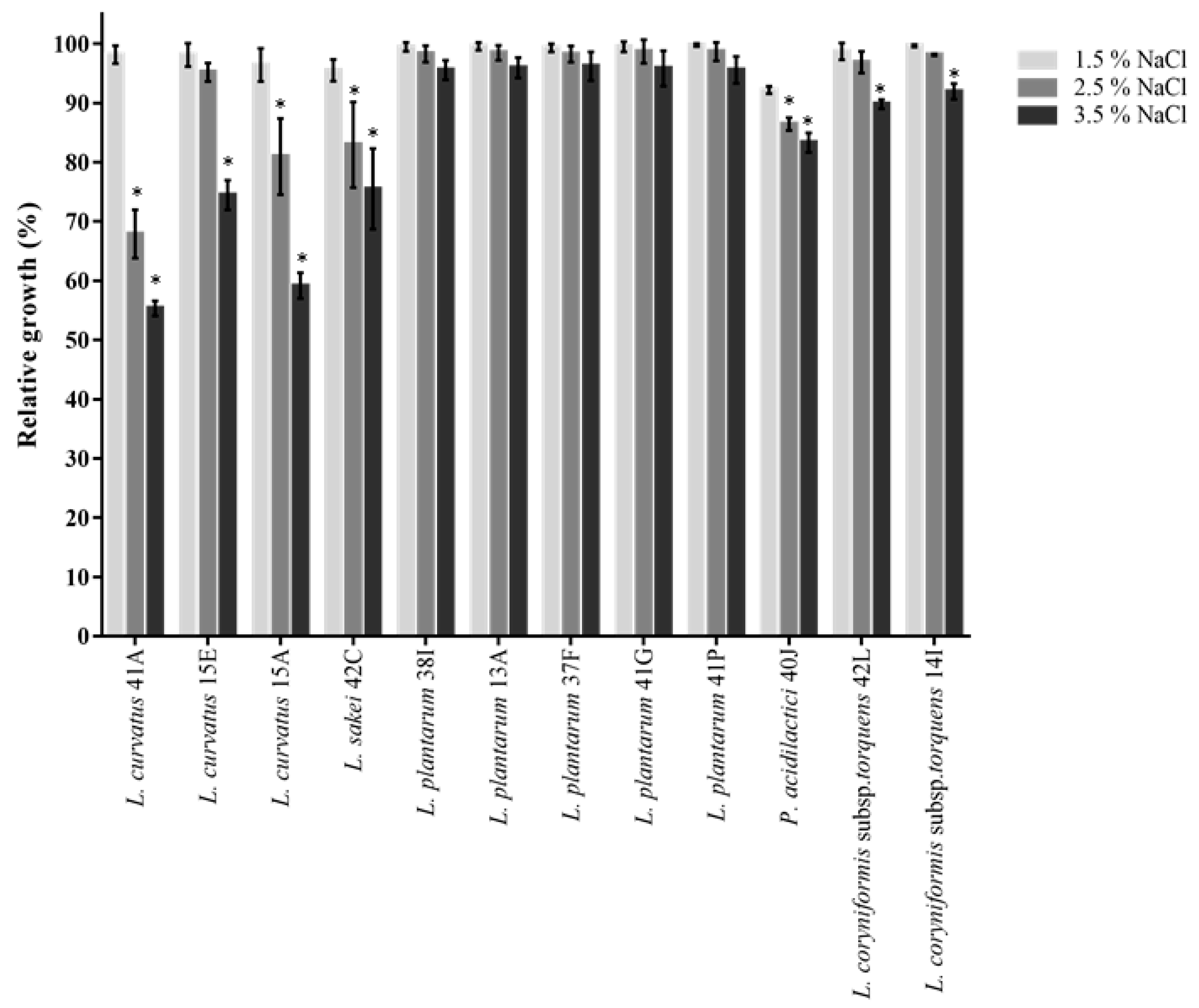
During industrial processing, probiotic LAB may encounter osmotic stress due to changes in solute concentration in the environment, which could result in cellular hydration and dehydration and this may negatively affect the survival rate and influence metabolic activities of probiotics [
84]. Probiotic strains could be added to fermented foods containing varying concentrations of salt. Thus, tolerance to NaCl may offer a competitive advantage for probiotics. For example, spontaneous sauerkraut fermentations operate optimally with a concentration of 2.5% (
w/
w) NaCl [
85], while 2 to 3% salt in meat fermentation was demonstrated to improve meat texture [
86]. All tested isolates were resistant to 1.5% NaCl, with
L. plantarum being the most resistant to higher NaCl concentrations (
Figure 5). Relatively high tolerance to 2.5% and 3.5% NaCl exposure was also observed for all isolates of
L. coryniformis subsp.
torquens (89 to 98%),
P. acidilactici 40J (83 to 86%) and
L. curvatus 15E (74 to 95%). Overall,
L. curvatus 41A, 15A and
L. sakei 42C demonstrated moderate tolerance to 2.5% (67 to 83% growth) and 3.5% (55 to 75% growth).
Current literature asserts that BSH activity offers a selective advantage for probiotic strains to survive and perform in the intestinal milieu [
87]. However, it is debatable whether BSH activity provides desirable effects for the human host, as there is concern over the safety of administering BSH-positive probiotics. However, research has shown that in the case of bifidobacteria and lactobacilli, BSH-positive probiotics may not be able to dehydroxylate deconjugated bile salts and thus the majority of products resulting from BSH activity may be precipitated and excreted in feces [
87]. BSH activity was assessed on selected probiotic candidates in the present study by growing them on MRS agar containing 0.5% TDCA and it was found that strains 38I, 41G, 42L and 14I produced BSH and
bsh gene homologues were identified in the genome of these strains (
Table S4). However, a
bsh gene homologue was also identified in the genome of
P. acidilactici 40J, which indicates that there could be a low level of BSH expression under the circumstances the bacteria were grown in or that the plate assay containing TDCA might not be the most appropriate method to test BSH activity in this strain. Other studies suggested the use of thin layer chromatography (TLC) assay as a more sensitive qualitative method to identify BSH-positive LAB [
88]. The presence of other probiotic gene markers such as
mub (mucin binding protein) and
fbp (fibronectin binding protein), which are known to play a role in promoting probiotic adherence to the host gastrointestinal tract, was also investigated [
89]. Strains 38I, 41G, 40J, 42L and 14I were found to possess homologues of the
mub and
fbp genes (
Table S4).


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}