
Lactobacillus fermentum SMFM2017-NK4 Isolated from Kimchi Can Prevent Obesity by Inhibiting Fat Accumulation

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Lactic Acid Bacteria
2.2. Measurement of α-Glucosidase Inhibition Activity
- AbsS: absorbance of a sample
- AbsSB: absorbance of a blank of the sample
- AbsC: absorbance of a control sample
- AbsCB: absorbance of a blank of the control sample
2.3. Measurement of Pancreatic Lipase Inhibition Activity
- AbsS: absorbance of a sample
- AbsSB: absorbance of a blank of the sample
- AbsRC: absorbance of a control sample
- AbsB: absorbance of a blank of the control sample
2.4. Measurement of Inhibition for 3T3-L1 Adipocyte Differentiation
2.5. Development of Obese Mouse Model and Administration of Lactic Acid Bacteria
2.6. Analysis of Serum Cholesterol and Lipid
2.7. Pathological Analysis of Adipose and Liver Tissue
2.8. Transcriptome Analysis of the Liver Tissue
2.9. Immunoblotting
2.10. Statistical Analysis
3. Results/Discussion
3.1. Inhibition of α-Glucosidase Activity by Lactic Acid Bacteria
3.2. Inhibition of Pancreatic Lipase Activity by Lactic Acid Bacteria
3.3. Inhibition of 3T3-L1 Cell Differentiation by Lactic Acid Bacteria
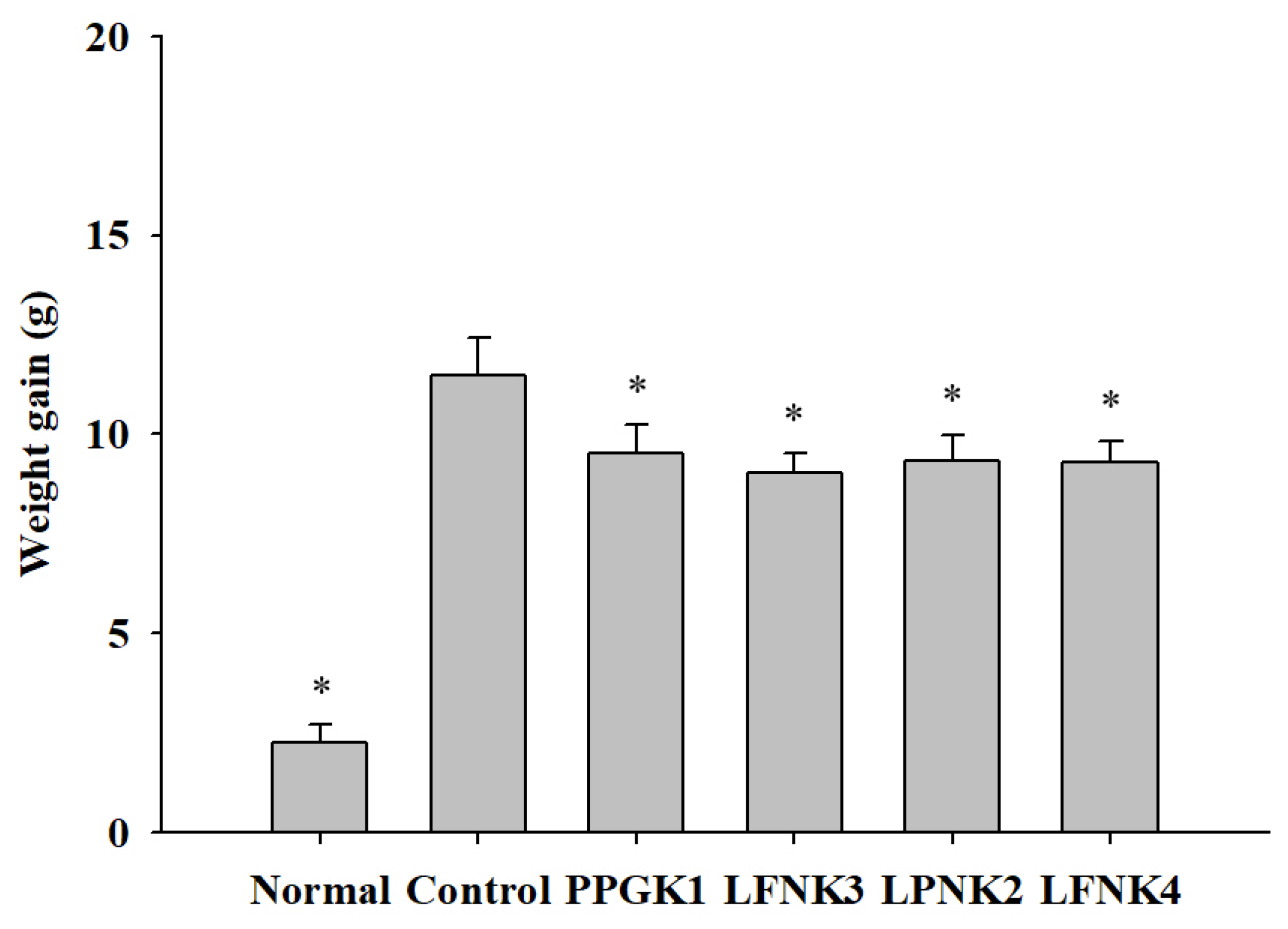
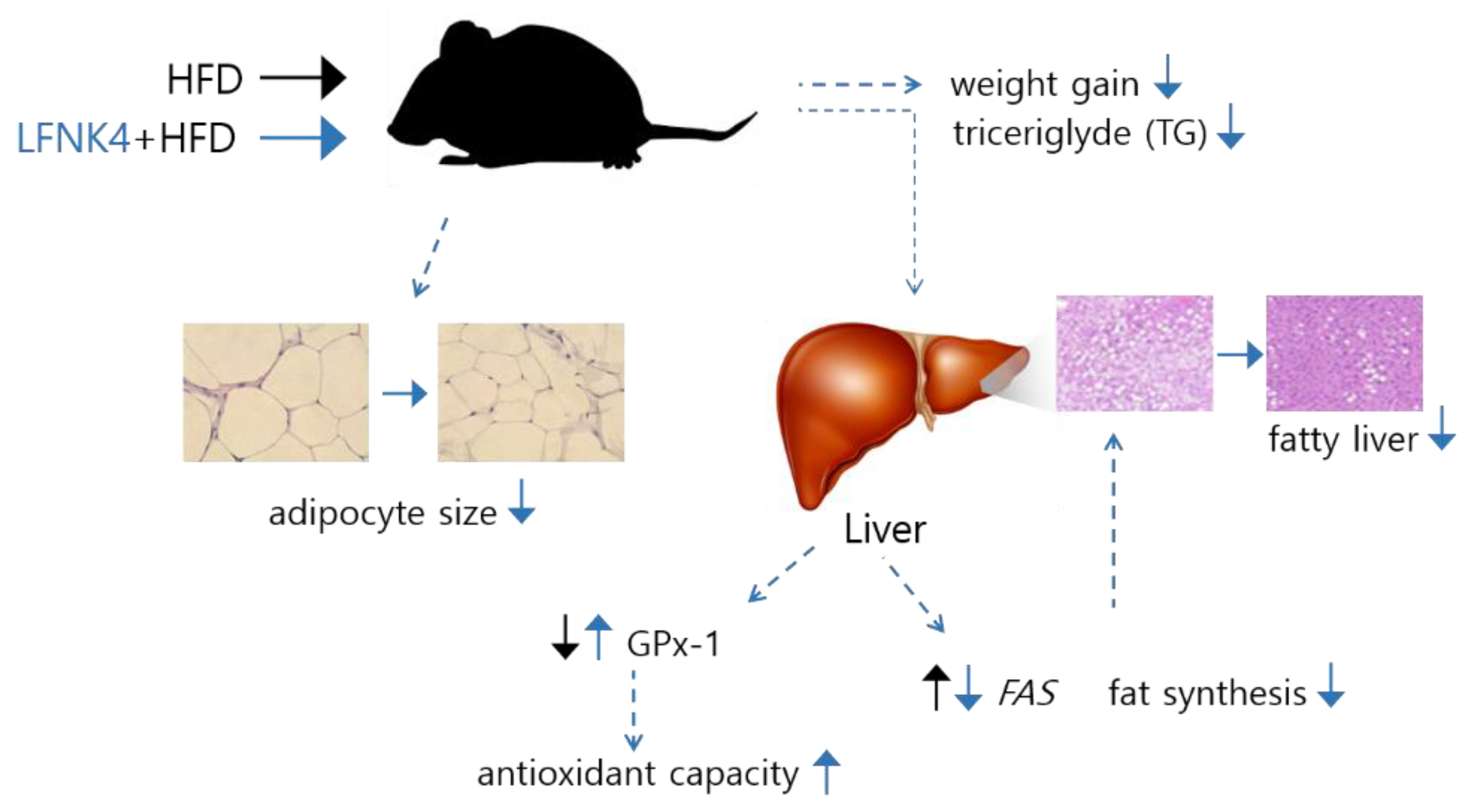
3.4. Weight Gain of the Obese Mouse Model
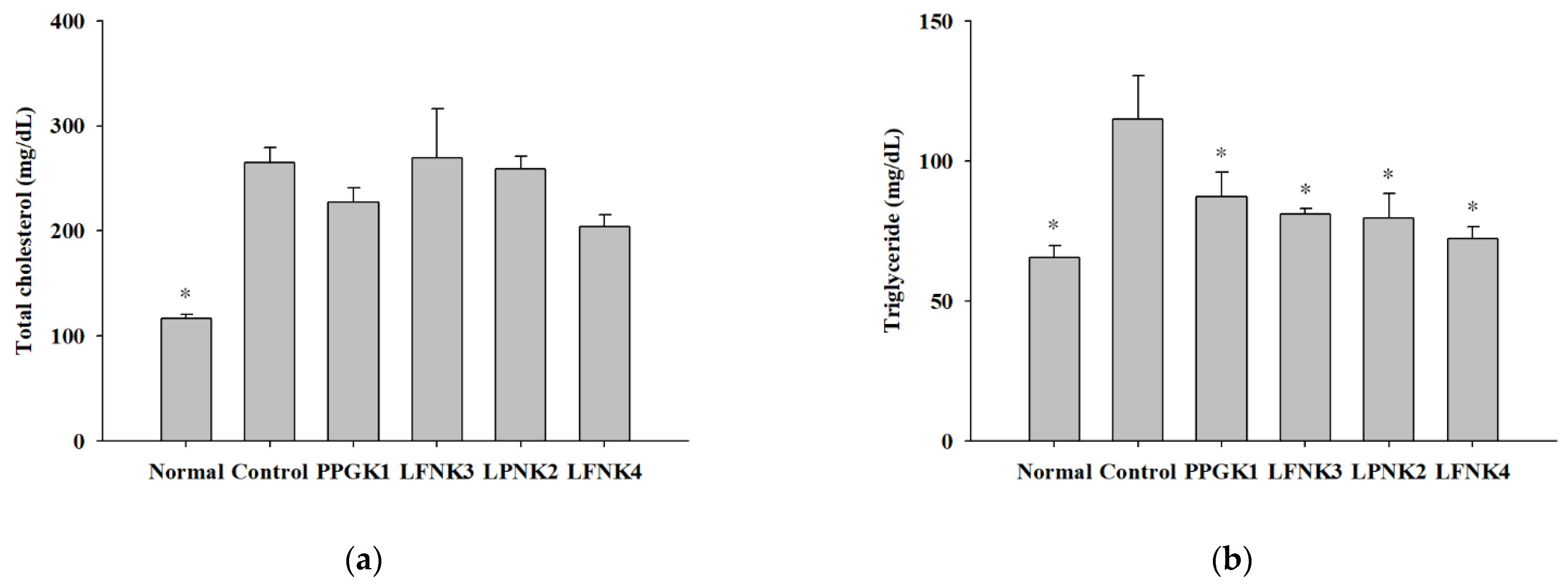
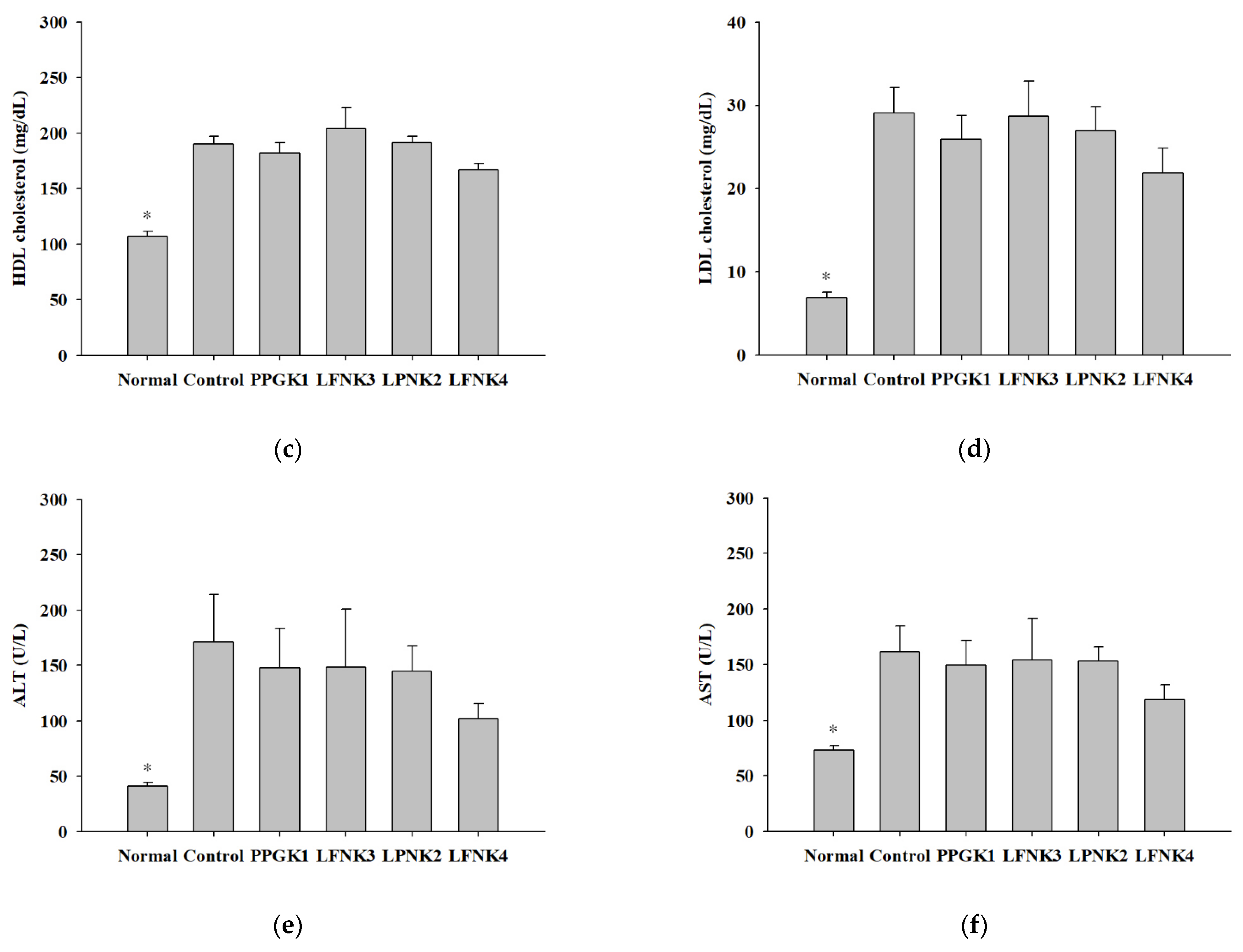
3.5. Indicators of Lipid and Inflammation-Related Markers in Blood Serum
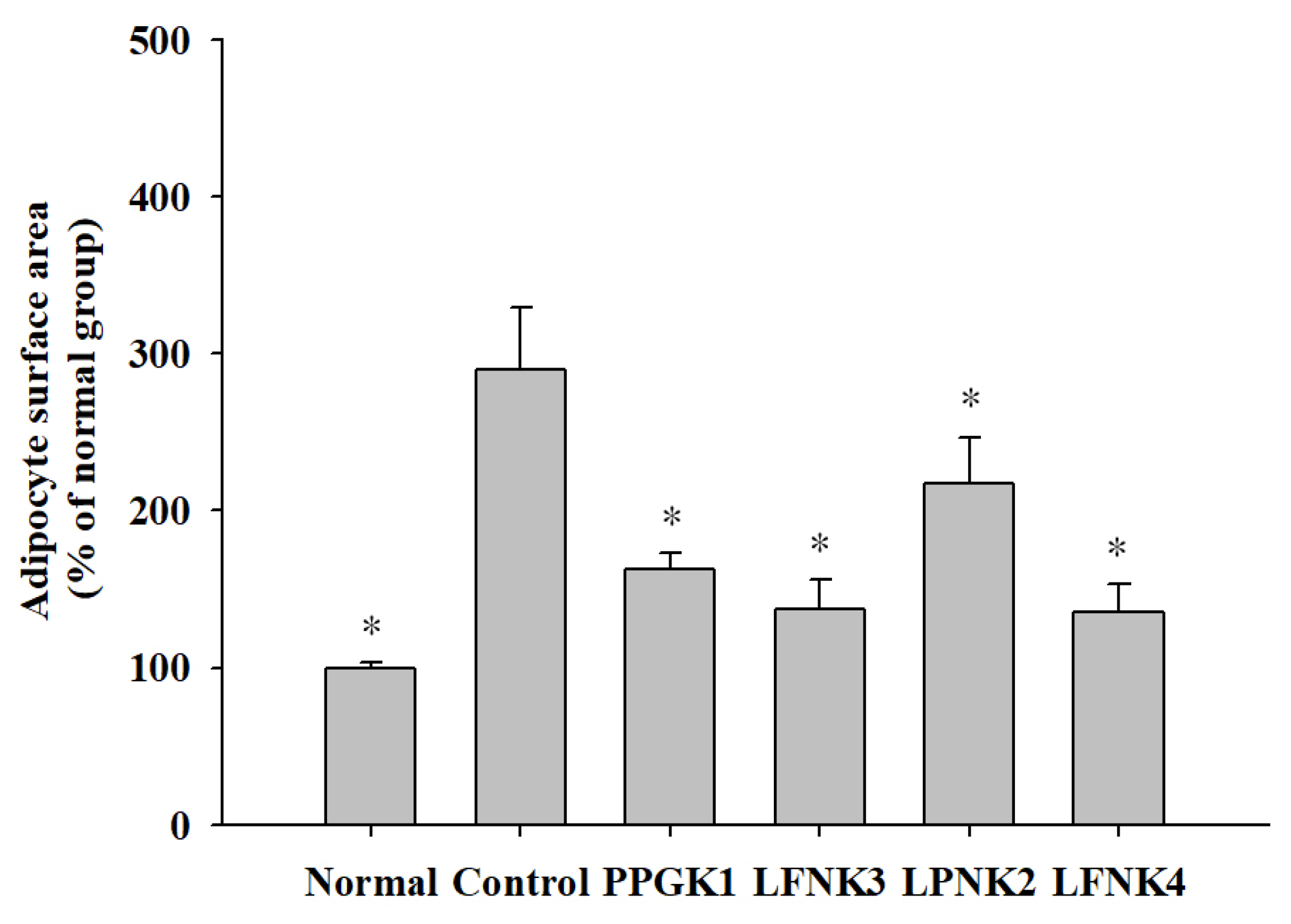
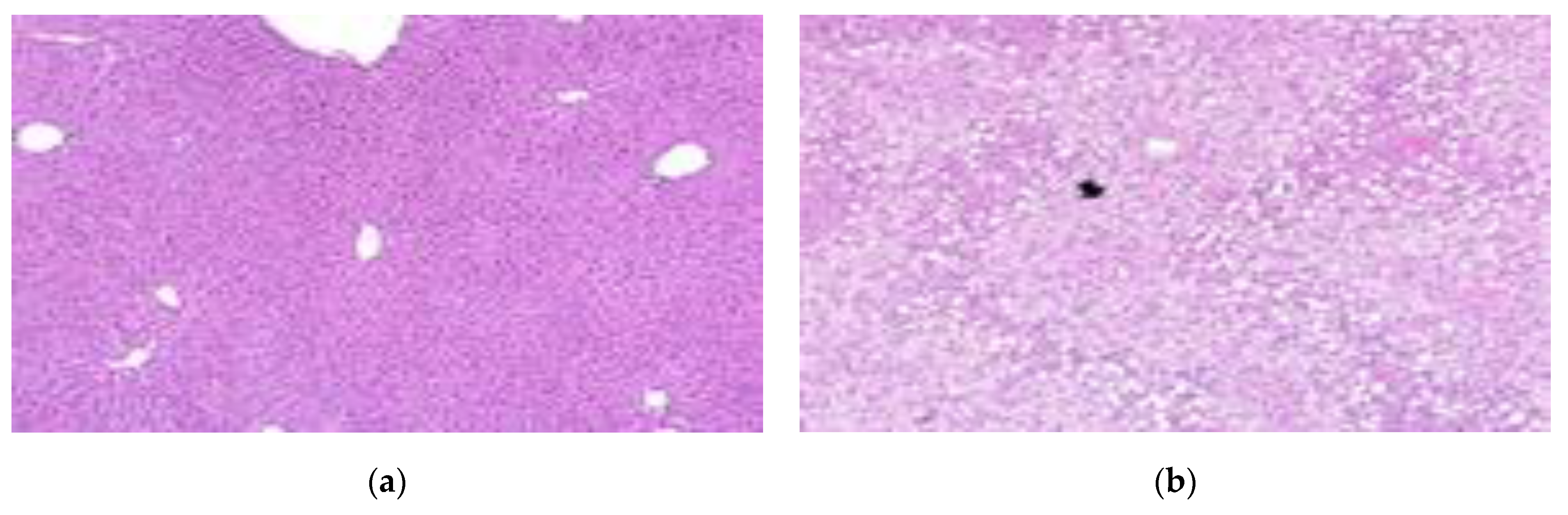
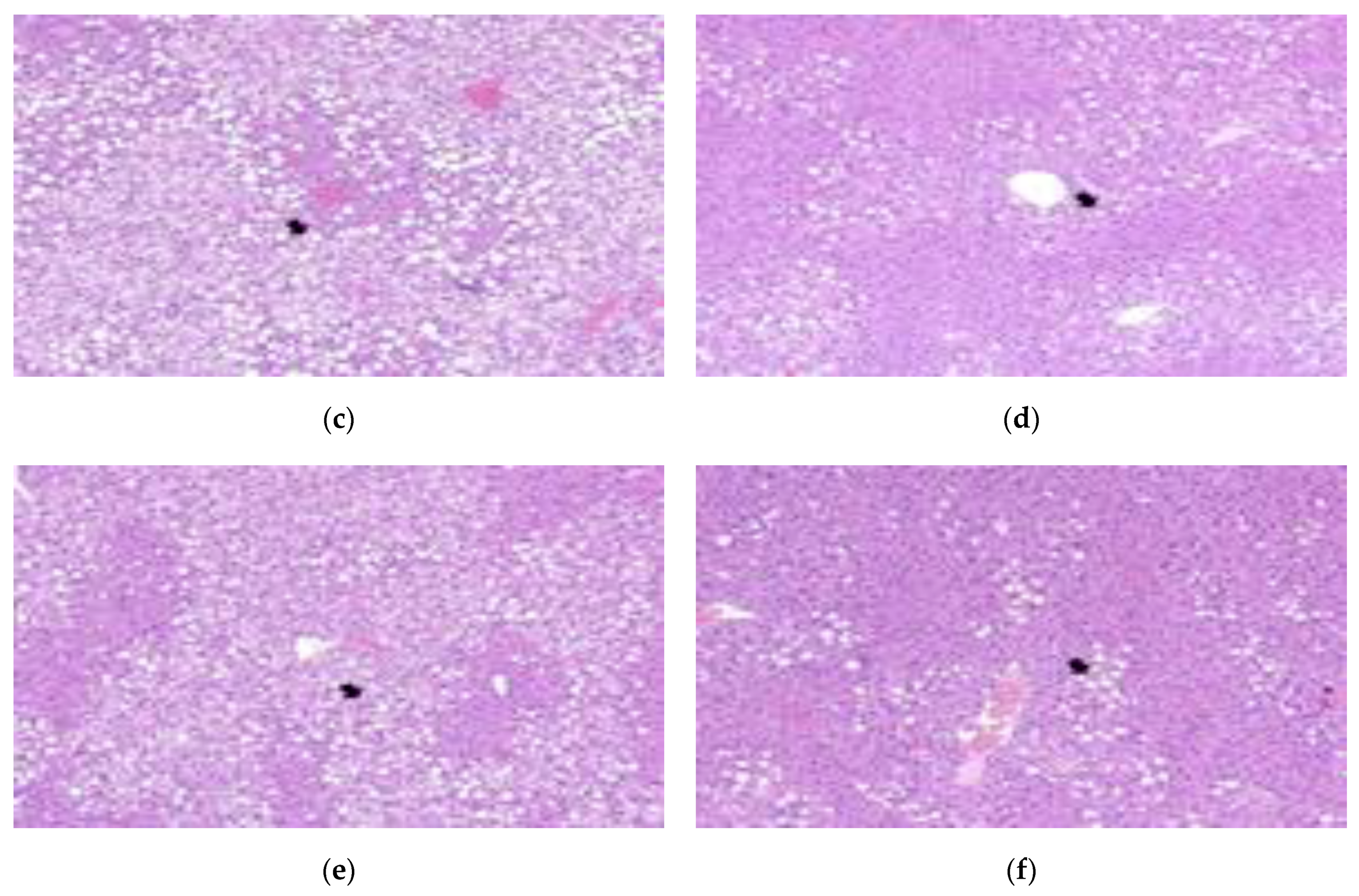
3.6. Pathological Features of Adipose and Liver Tissue
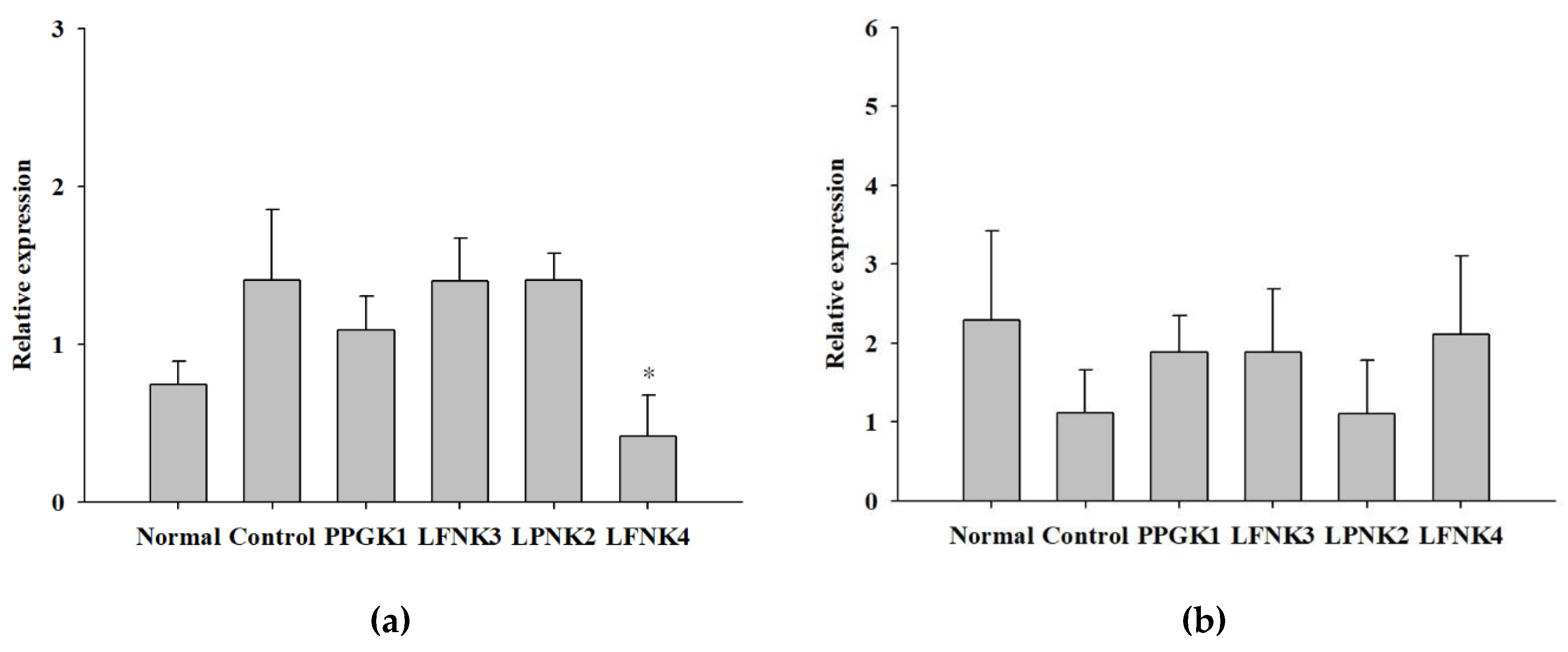
3.7. Transcriptome in the Liver Tissue
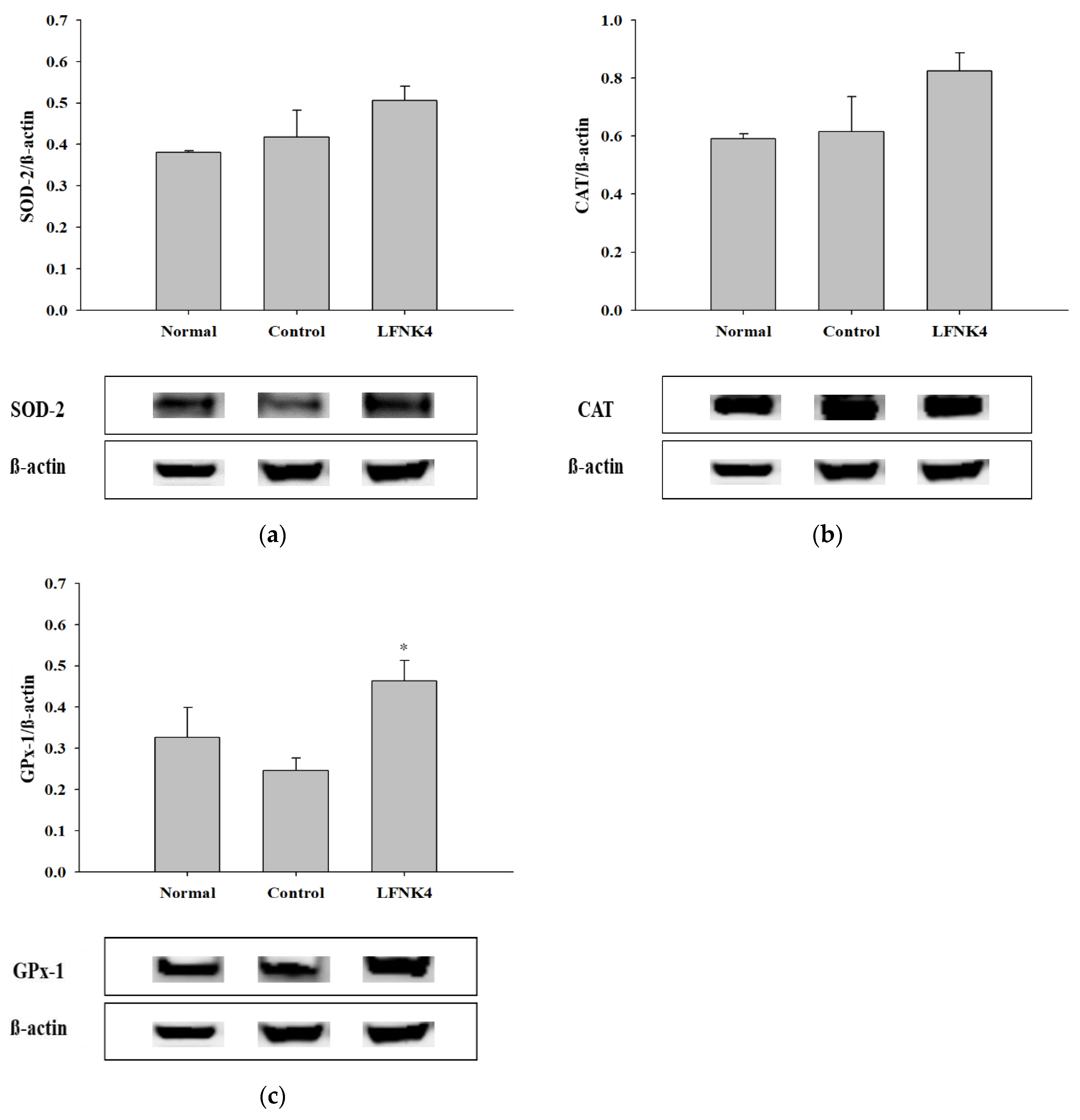
3.8. Protein Expression Related to Antioxidant Enzymes in the Liver Tissue
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Kim, S.K.; Lim, S.D. Separation and purification of lipase inhibitory peptide from fermented milk by Lactobacillus plantarum Q180. Food Sci. Anim. Resour. 2020, 40, 87–95. [Google Scholar] [CrossRef]
- Lee, J.S.; Hyun, I.K.; Yoon, J.W.; Seo, H.J.; Kang, S.S. Bioconversion products of whey by lactic acid bacteria exert anti-adipogenic effect. Food Sci. Anim. Resour. 2021, 41, 145–152. [Google Scholar] [CrossRef] [PubMed]
- McGee, D.L. Body mass index and mortality: A meta analysis based on person-level data from twenty-six observational studies. Ann. Epidemiol. 2005, 15, 87–97. [Google Scholar] [CrossRef] [PubMed]
- KCDC (Korea Centers for Disease Control and Prevention). Trends in Prevalence of Obesity among Korean Adults 2007–2016. 2018. Available online: http://www.cdc.go.kr/board.es?mid=a20602010000&bid=0034&act=view&list_no=140978 (accessed on 29 June 2020).
- Lee, K.S.; Kim, M.G.; Lee, N.Y. Antimicrobial effect of the extracts of Cactus Cheonnyuncho (Opuntiahumifusa) against food borne pathogens. J. Korean Soc. Food. Nutr. 2004, 33, 1268–1272. [Google Scholar]
- Lee, Y.A.; Kim, M.H. The effect of sea tangle extract on serum lipid level in ovariectomized rats. J. Life Sci. 2008, 18, 249–254. [Google Scholar] [CrossRef]
- Lee, S.J.; Park, H.J.; Song, Y.O.; Jang, S.H.; Goo, J.; Ko, Y.G.; Cho, J.H. Antioxidant activity and anti-obesity effect of Coprinus comatus in Zucker rat (fa/fa). Korean J. Vet. Serv. 2014, 37, 51–58. [Google Scholar] [CrossRef] [Green Version]
- De Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef] [PubMed]
- Wulandari, E.; Yurmiati, H.; Subroto, T.; Suradi, K. Quality and probiotic lactic acid bacteria diversity of rabbit meat Bekasam-fermented meat. Food Sci. Anim. Resour. 2020, 40, 362–376. [Google Scholar] [CrossRef]
- Kariyawasam, K.M.G.M.M.; Yang, S.J.; Lee, N.K.; Paik, H.D. Probiotic properties of Lactobacillus brevis KU200019 and synergistic activity with fructooligosaccharides in antagonistic activity against foodborne pathogens. Food Sci. Anim. Resour. 2020, 40, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, K.; He, F.; Yoda, K.; Hiramatsu, M. Potent effects of, and mechanisms for, modification of cross-talk between macrophages and adipocytes by lactobacilli. Microbiol. Immunol. 2012, 56, 847–854. [Google Scholar] [CrossRef]
- Choi, H.J.; Oh, Y.J.; Jang, J.Y.; Lee, J.E.; Kwon, M.S.; Lim, S.K. Weissella cibaria WIKIM28 Having Anti-Obesity Activity and Composition for Comprising the Same. KR Patent 1834383, 26 February 2018. [Google Scholar]
- Choi, H.J.; Oh, Y.J.; Jang, J.Y.; Lee, J.E. Weissella confusa WIKIM51 Having Anti-Obesity Activity and Composition for Comprising the Same. KR Patent 1834379, 26 February 2018. [Google Scholar]
- Seo, J.Y.; Kim, J.R.; Yoo, B.W. Interpretation of liver function test. Korean J. Fam. Pract. 2012, 2, 207–213. [Google Scholar]
- Kwon, J.Y.; Cheigh, H.S.; Song, Y.O. Weight reduction and lipid lowering effects of kimchi lactic acid powder in rats fed high fas diets. Korean J. Food. Sci. Technol. 2004, 36, 1014–1019. [Google Scholar]
- Jang, H.J. Potential Use of Lactic Acid Bacteria Isolated from Kimchi as Probiotics. Master’s Thesis, Sookmyung Women’s University, Seoul, Korea, 2018. [Google Scholar]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Ana, M.R.; Thomas, P.B. Lipase inhibitory activity of skim milk fermented with different strains of lactic acid bacteria. J. Funct. Foods 2019, 60, 103413. [Google Scholar]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Kim, K.A.; Gu, W.; Lee, I.A.; Joh, E.H.; Kim, D.H. High fat diet-induced gut microbiota exacerbates infalmmation and obesity in mice via the TLR4 signaling pathway. PLoS ONE 2012, 7, e47713. [Google Scholar]
- Ritze, Y.; Bárdos, G.; Claus, A.; Ehrmann, V.; Berqheim, I.; Schwiertz, A.; Bischoff, S.C. Lactobacillus rhamnosus GG protects against non-alcholic fatty liver disease in mice. PLoS ONE 2014, 9, e80169. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.R.; Kim, Y.J.; Park, D.Y.; Jung, U.J.; Jeon, S.M.; Ahn, Y.T.; Huh, C.S.; McGregor, R.; Choi, M.S. Probiotics L. plantarum and L. curvatus in combination alter hepatic lipid metabolism and suppress diet-induced obesity. Obesity 2013, 21, 2421–2603. [Google Scholar] [CrossRef] [PubMed]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [CrossRef]
- Bertozzi, C.R.; Kiessling, L.L. Chemical glycobiology. Science 2001, 23, 2357–2364. [Google Scholar] [CrossRef] [Green Version]
- Ko, K.H.; Kim, J.Y.; Kim, Y.G.; Oh, K.Y.; Goo, Y.M.; Son, Y.H.; Park, B.R.; Kim, G.S.; Lim, J.H.; Nam, S.H. Isolation and identification of α-glucosidase inhibitory compounds from artemisia annua L. leaves and stems. J. Agric. Life Sci. 2016, 50, 161–173. [Google Scholar] [CrossRef]
- Yoshio, K. Obesity and Related Diseases; Koonja Publishing Company: Seoul, Korea, 2005; pp. 691–692. [Google Scholar]
- EI-shiekh, R.A.; AI-Mahdy, D.A.; Hifnawy, M.S.; Abdel-Sattar, E.A. In-vitro screening of selected traditional medicinal plants for their anti-obesity and anti-oxidant activities. J. Bot. 2019, 123, 43–50. [Google Scholar]
- Kim, S.K.; Lim, S.D. Physiological characteristics and anti-obesity effect of milk fermented by Lactobacillus plantarum KI134. J. Dairy Sci. Biotechnol. 2020, 38, 207–221. [Google Scholar] [CrossRef]
- Charles, M. Fat digestion and absorption. In Encyclopeida of Gastroenterology; Leonard, R.J., Ed.; Academic Press: Cambridge, MA, USA, 2004; pp. 23–30. [Google Scholar]
- Bray, G.A. Medications for obesity: Mechanisms and applications. Clin. Chest Med. 2009, 30, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Lee, S.Y.; Lee, A.R. The effect of Selaginella tamariscina on inhibition of pancreatic lipase and lipid accumulation. J. Korean Food. Nutr. 2018, 32, 27–32. [Google Scholar]
- Kang, C.H.; Jeong, Y.A.; Han, S.H.; Kim, J.S.; Kim, Y.G.; Park, H.M.; Choi, S.I.; Paek, N.S. In vitro probiotic evaluation of potential antiobesity lactic acid bacteria isolated from human vagina and shellfish. KSBB J. 2018, 33, 161–167. [Google Scholar] [CrossRef]
- Kim, E.J.; Kang, M.J.; Seo, Y.B.; Nam, S.W.; Kim, G.D. Acer okamotoanum Nakai leaf extract inhibits adipogenesis via suppressing expression of PPAR γ and C/EBP α in 3T3-L1 cells. J. Microbiol. Biotechnol. 2018, 28, 1645–1653. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Jeong, J.E.; Moon, S.H.; Park, K.Y. Anti-obesity effect of Bacillus subtilis KC-3 fermented soymilk in 3T3-L1 adipocytes. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1126–1131. [Google Scholar]
- Song, M.Y.; Bose, S.; Kim, H.J. Effect of probiotics-fermented Samjunghwan on differentiation in 3T3-L1 preadipocytes. J. Korean Soc. Food Sci. Nutr. 2013, 42, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Lim, S.D. Physiological characteristics and anti-obesity effect of Lactobacillus plantarum K6 isolated from Kimchi. J. Milk Sci. Biotechnol. 2017, 35, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Park, H.E.; Do, K.H.; Jeong, J.W.; Jung, Y.H.; Lee, W.K. Cholesterol and visceral fat owering effects of combined lactic acid bacteria (Lactobacillus casei WK3, Enterococus faecium WK5, Bifidobacterium longum WK9 and Lactobacillus plantarum K-1) in high fat diet-fed C57BL/6J Mice. J. Korean Soc. Food Sci. Nutr. 2018, 12, 1210–1216. [Google Scholar] [CrossRef]
- Go, G.Y.; Park, J.H.; Kwak, J.Y.; Ahn, T.W. Anti-obesity effect of yeoldahanso-tang on obesity mice. J. Sassng Const. Med. 2017, 29, 154–173. [Google Scholar]
- Ryu, H.J.; Um, M.Y.; Ahn, J.Y.; Jung, C.H.; Huh, D.; Kim, T.W.; Ha, T.Y. Anti-obesity effect of Hypsizigus marmoreus in high fat-fed mice. J. Korean Soc. Food Sci. Nutr. 2011, 40, 1708–1714. [Google Scholar] [CrossRef]
- Song, M.Y.; Jung, H.W.; Park, Y.K. Anti-obesity effect of water extract of Coix lacrymajobi var. mayuen in high fat fed C5BL/6 mice. J. Korean Med. Obes. Res. 2016, 16, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Bob, T.; Robert, R.M.; Takanori, H.; Abraham, N.; Colin, R.; Thomas, N.; David, E.M.; Wolfgang, K.; Karin, K.; Ulrich, D.; et al. Proliferative and nonproliferative lesions of the rat and mouse hepatobiliary system. J. Toxicol. Pathol. 2010, 38, 5S–81S. [Google Scholar]
- Yu, Z.; Mao, C.; Fu, X.; Ma, M. High density lipoprotein from egg yolk (EYHDL) improves dyslipidemia by mediating fatty acids metabolism in high fat diet-induced obese mice. Food Sci. Anim. Resour. 2019, 39, 179–196. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.M.; Oh, H.G.; Kang, Y.G.; Kim, Y.P.; Sin, H.S.; Jang, S.H.; Kim, H.J.; Lee, S.W.; Lee, H.Y. Anti-obesity effects of lactic acid bacteria-fermented Dioscoreae Rhizoma powder on high fat diet-fed animal model. J. Physiol. Pathol. Korean Med. 2015, 29, 267–272. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, H.S.; Seo, S.W. Anti-obesity effect of Aster yomena ethanol extract in high fat diet-induced obese mice. J. Physiol. Pathol. Korean Med. 2017, 31, 348–355. [Google Scholar] [CrossRef]
- Hur, H.J.; Hwang, J.T. Effect of silybin on body weight and glucose tolerance in high-fat-diet induced obese mice. KSBB J. 2011, 26, 78–82. [Google Scholar] [CrossRef]
- Park, S.J.; Choi, C.Y.; Jeong, J.C.; Lee, R.K.; Hwang, Y.P. Artemisia annua extract ameliorates high-fat diet-induced fatty liver by activating AMPK. J. Biomed. Transl. Res. 2020, 21, 59–71. [Google Scholar] [CrossRef]
- Ikarashi, N.; Toda, T.; Okaniwa, T.; Ito, K.; Ochiai, W.; Sugiyama, K. Anti-obesity and anti-diabetic effects of Acacia polyphenol in obese diabetic KKAy mice fed high-fat diet. Evid. Based Complement. Alternat. Med. 2011, 2011, 952031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.R.; Choi, B.K.; Jung, H.J. Anti-obesity effects of mixture of Atractylodes macrocephala and Amomum villosum extracts. J. Physiol. Pathol. Korean Med. 2019, 33, 282–287. [Google Scholar] [CrossRef]
- Cho, J.K.; Kang, H.S.; Yoon, J.H. Exercise strategies for the prevention and treatment of non-alcoholic fatty liver disease. Korean J. Obes. 2015, 24, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.B.; Lee, J.W.; Hong, Y.M. Serum fatty acid and carnitine levels in obese children with fatty livers. J. Korean Pediatr. Soc. 2002, 45, 1083–1089. [Google Scholar]
- Lee, J.W.; Hong, Y.M. Changes of serum fatty acid and carnitine levels after administration of L-carnitine in rats. J. Korean Pediatr. Soc. 2002, 45, 1075–1082. [Google Scholar]
- Kim, T.R.; Kim, Y.J.; Woo, C.H. Anti-obesity effects of Banggihwnggi-tang-hap-yeonggyechulgam-tang in high fat diet induced obese mice. J. Korean Med. Rehabil. 2019, 29, 29–45. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lactic Acid Bacteria Strains | Identification |
|---|---|
| 1, 44 | Lactobacillus pentosus |
| 3, 28, 29, 31, 32, 66, 70, 73, 52-1-2 | Lactobacillus fermentum |
| 6, 50, 76, 77, 78, 79, 38-Y | Lactobacillus plantarum |
| 7, 9, 11, 20 | Pediococcus pentosaceus |
| 12, 75 | Weissella cibaria |
| 26, 30 | Lactobacillus sakei |
| 45, 72 | Lactobacillus spp. |
| 71, 4Y | Lactobacillus brevis |
| 98-1-3 | Lactobacillus curvatus |
| Gene | Primer Sequence (5′→ 3′) | Reference | |
|---|---|---|---|
| β-actin | Forward | CCG TGA AAA GAT GAC CCA GAT C | Kim et al., 2012 [20] |
| Reverse | CAC AGC CTG GAT GGC TAC GT | ||
| Fas | Forward | TCT GGG CCA ACC TCA TTG GT | Ritze et al., 2014 [21] |
| Reverse | GAA GCT GGG GGT CCA TTG TG | ||
| Cpt-2 | Forward | GCC TGC TGT TGC GTG ACT G | Yoo et al., 2013 [22] |
| Reverse | TGG GTA CGA TGC TGT GC | ||
| Lactic Acid Bacteria Strains | Inhibition Rate | Lactic Acid Bacteria Strains | Inhibition Rate |
|---|---|---|---|
| 66 | 41.45 ± 6.02 a | 72 | 29.02 ± 2.22 ab |
| 29 | 40.64 ± 3.76 ab | 32 | 27.85 ± 7.93 b |
| 50 | 39.13 ± 6.81 ab | 1 | 27.60 ± 16.90 b |
| 70 | 39.09 ± 4.49 ab | 7 | 27.35 ± 4.65 b |
| 75 | 37.22 ± 4.43 ab | 79 | 25.78 ± 9.50 b |
| 45 | 37.04 ± 4.04 ab | 52-1-2 | 25.69 ± 13.28 b |
| 44 | 34.64 ± 1.15 ab | 78 | 23.96 ± 9.31 bc |
| 73 | 33.11 ± 3.93 ab | 77 | 23.35 ± 4.69 bc |
| 20 | 30.68 ± 2.00 ab | 26 | 22.66 ± 2.96 bc |
| 98-1-3 | 30.16 ± 3.73 ab | 9 | 19.48 ± 5.29 bc |
| 31 | 30.00 ± 9.45 ab | 12 | 17.17 ± 9.19 bc |
| 38-Y | 29.90 ± 10.90 ab | 3 | 15.23 ± 9.35 bc |
| 30 | 29.59 ± 9.94 ab | 11 | 12.27 ± 9.00 c |
| 76 | 29.52 ± 5.65 ab | 6 | 10.31 ± 12.71 c |
| 4Y | 29.48 ± 10.71 ab | 28 | 6.58 ± 5.50 c |
| 71 | 29.02 ± 9.22 ab |
| Lactic Acid Bacteria Strains | Inhibition Rate | Lactic Acid Bacteria Strains | Inhibition Rate |
|---|---|---|---|
| 20 | 18.92 ± 4.27 a | 70 | 4.61 ± 0.94 bc |
| 11 | 18.80 ± 7.36 a | 44 | 3.25 ± 4.90 bc |
| 26 | 18.10 ± 1.76 ab | 12 | 2.91 ± 3.52 bc |
| 30 | 15.22 ± 6.93 ab | 79 | 2.76 ± 1.55 bc |
| 29 | 14.43 ± 5.27 ab | 45 | 2.40 ± 0.84 bc |
| 73 | 13.74 ± 4.44 ab | 71 | 2.24 ± 0.93 bc |
| 72 | 10.96 ± 5.39 ab | 66 | 2.13 ± 1.38 bc |
| 50 | 10.95 ± 5.87 ab | 78 | 2.05 ± 1.54 bc |
| 38-Y | 10.39 ± 7.99 ab | 76 | 1.71 ± 2.83 bc |
| 31 | 10.15 ± 3.44 ab | 6 | 0.01 ± 6.87 bc |
| 4Y | 9.03 ± 5.98 b | 9 | −1.60 ± 3.89 c |
| 98-1-3 | 8.86 ± 8.91 bc | 75 | −1.63 ± 4.30 c |
| 32 | 7.22 ± 3.74 bc | 52-1-2 | −2.63 ± 3.54 c |
| 7 | 7.05 ± 0.84 bc | 3 | −7.78 ± 7.01 c |
| 77 | 6.91 ± 6.84 bc | 28 | −10.58 ± 16.38 c |
| 1 | 5.21 ± 4.52 bc |
| Lactic Acid Bacteria Strains | Inhibition Rate | |
|---|---|---|
| 73 | L. fermentum SMFM2017-NK4 | 21.44 ± 6.70 a |
| 29 | L. fermentum SMFM2017-NK3 | 20.99 ± 10.06 a |
| 20 | P. pentosaceus SMFM2017-GK1 | 20.45 ± 2.53 a |
| 50 | L. plantarum SMFM2017-NK2 | 15.57 ± 7.37 a |
| Groups | ||||||
|---|---|---|---|---|---|---|
| Normal | Control | PPGK1 | LFNK3 | LPNK2 | LFNK4 | |
| Steatosis | ± | +++ | +++ | ++ | +++ | ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Choi, Y.; Kim, S.; Ha, J.; Oh, H.; Lee, Y.; Kim, Y.; Seo, Y.; Park, E.; Kang, J.; et al. Lactobacillus fermentum SMFM2017-NK4 Isolated from Kimchi Can Prevent Obesity by Inhibiting Fat Accumulation. Foods 2021, 10, 772. https://doi.org/10.3390/foods10040772
Kim D, Choi Y, Kim S, Ha J, Oh H, Lee Y, Kim Y, Seo Y, Park E, Kang J, et al. Lactobacillus fermentum SMFM2017-NK4 Isolated from Kimchi Can Prevent Obesity by Inhibiting Fat Accumulation. Foods. 2021; 10(4):772. https://doi.org/10.3390/foods10040772
Chicago/Turabian StyleKim, Doyeon, Yukyung Choi, Sejeong Kim, Jimyeong Ha, Hyemin Oh, Yewon Lee, Yujin Kim, Yeongeun Seo, Eunyoung Park, Joohyun Kang, and et al. 2021. "Lactobacillus fermentum SMFM2017-NK4 Isolated from Kimchi Can Prevent Obesity by Inhibiting Fat Accumulation" Foods 10, no. 4: 772. https://doi.org/10.3390/foods10040772