
Adjustment of Whey:Casein Ratio from 20:80 to 60:40 in Milk Formulation Affects Food Intake and Brainstem and Hypothalamic Neuronal Activation and Gene Expression in Laboratory Mice

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Milk Formulations
2.3. Feeding Studies
2.3.1. Preference for the Simultaneously Presented Formulations
2.3.2. Energy Deprivation-Induced 2-h Intake of each Formulation Presented Individually Along with Standard Chow
2.3.3. Intake of the Formulations Presented Individually for 24 h
2.4. Neuronal Activation in Feeding-Related Hypothalamic and Brainstem Areas after Consumption of the Same Amount of the Control 60:40 versus 20:80 Whey:Casein Milk Formulation
2.5. Hypothalamic and Brainstem Gene Expression Following 24-h Exposure to the Control 20:80 versus 60:40 Whey:Casein Milk Formulation
2.6. Statistical Analyses
3. Results
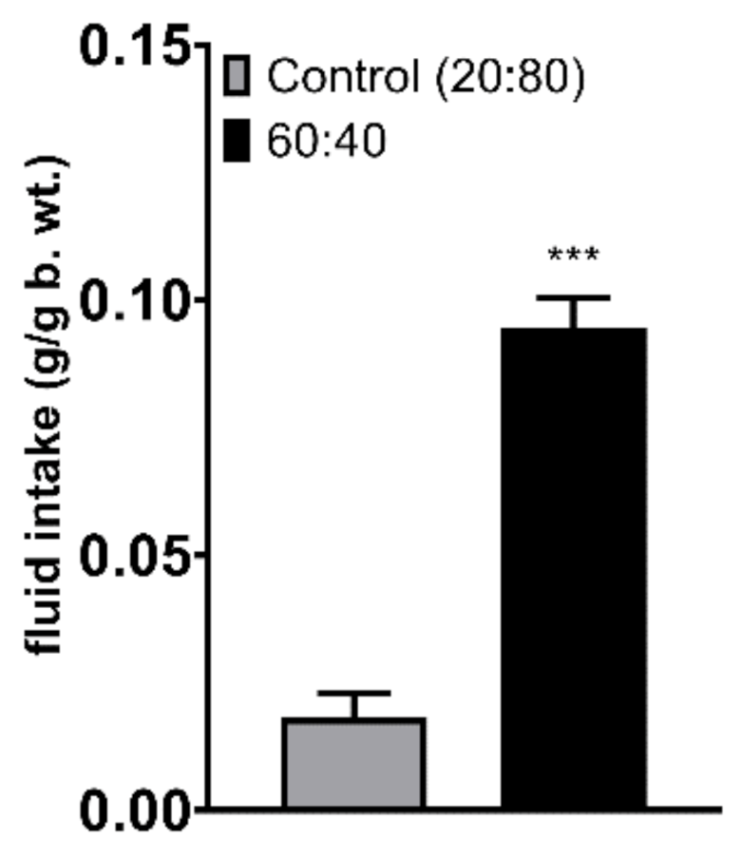
3.1. Feeding Studies
3.2. c-Fos Immunoreactivity
3.3. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, G.H.; Moore, S.E. Dietary Proteins in the Regulation of Food Intake and Body Weight in Humans. J. Nutr. 2004, 134, 974S–979S. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Sun, Q.; Giovannucci, E.; Mozaffarian, D.; Manson, J.E.; Willett, W.C.; Hu, F.B. Dairy consumption and risk of type 2 diabetes: 3 cohorts of US adults and an updated meta-analysis. BMC Med. 2014, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Hirahatake, K.M.; Slavin, J.L.; Maki, K.C.; Adams, S.H. Associations between dairy foods, diabetes, and metabolic health: Potential mechanisms and future directions. Metabolism 2014, 63, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Pasiakos, S.M. Metabolic Advantages of Higher Protein Diets and Benefits of Dairy Foods on Weight Management, Glycemic Regulation, and Bone. J. Food Sci. 2015, 80, A2–A7. [Google Scholar] [CrossRef]
- Jahan-Mihan, A.; Luhovyy, B.L.; El Khoury, D.; Anderson, G.H. Dietary Proteins as Determinants of Metabolic and Physiologic Functions of the Gastrointestinal Tract. Nutrients 2011, 3, 574–603. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.W.; Haenlein, G.F.W. (Eds.) Other minor species milk. In Milk and Dairy Products in Human Nutrition: Production, Composition and Health, 1st ed.; Wiley: Hoboken, NJ, USA, 2013; pp. 644–658. [Google Scholar]
- Goedhart, A.C.; Bindels, J.G. The Composition of Human Milk as a Model for the Design of Infant Formulas: Recent Findings and Possible Applications. Nutr. Res. Rev. 1994, 7, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Heird, W.C. Progress in Promoting Breast-Feeding, Combating Malnutrition, and Composition and Use of Infant Formula, 1981–2006. J. Nutr. 2007, 137, 499S–502S. [Google Scholar] [CrossRef] [Green Version]
- Lönnerdal, B. Nutritional and physiologic significance of human milk proteins. Am. J. Clin. Nutr. 2003, 77, 1537S–1543S. [Google Scholar] [CrossRef]
- Zemel, M.B. Role of calcium and dairy products in energy partitioning and weight management. Am. J. Clin. Nutr. 2004, 79, 907S–912S. [Google Scholar] [CrossRef] [Green Version]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.-P.; Maubois, J.-L.; Beaufrère, B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935. [Google Scholar] [CrossRef] [Green Version]
- Dangin, M.; Boirie, Y.; Garcia-Rodenas, C.; Gachon, P.; Fauquant, J.; Callier, P.; Ballèvre, O.; Beaufrère, B. The digestion rate of protein is an independent regulating factor of postprandial protein retention. Am. J. Physiol. Metab. 2001, 280, E340–E348. [Google Scholar] [CrossRef] [Green Version]
- Luiking, Y.C.; Abrahamse, E.; Ludwig, T.; Boirie, Y.; Verlaan, S. Protein type and caloric density of protein supplements modulate postprandial amino acid profile through changes in gastrointestinal behaviour: A randomized trial. Clin. Nutr. 2016, 35, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ye, A.; Lin, Q.; Han, J.; Singh, H. Gastric digestion of milk protein ingredients: Study using an in vitro dynamic model. J. Dairy Sci. 2018, 101, 6842–6852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalziel, J.E.; Young, W.; McKenzie, C.M.; Haggarty, N.W.; Roy, N.C. Gastric Emptying and Gastrointestinal Transit Compared among Native and Hydrolyzed Whey and Casein Milk Proteins in an Aged Rat Model. Nutrients 2017, 9, 1351. [Google Scholar] [CrossRef] [Green Version]
- Marsset-Baglieri, A.; Fromentin, G.; Airinei, G.; Pedersen, C.; Léonil, J.; Piedcoq, J.; Rémond, D.; Benamouzig, R.; Tomé, D.; Gaudichon, C. Milk protein fractions moderately extend the duration of satiety compared with carbohydrates independently of their digestive kinetics in overweight subjects. Br. J. Nutr. 2014, 112, 557–564. [Google Scholar] [CrossRef]
- Meyer, R.; Foong, R.-X.M.; Thapar, N.; Kritas, S.; Shah, N. Systematic review of the impact of feed protein type and degree of hydrolysis on gastric emptying in children. BMC Gastroenterol. 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calbet, J.A.L.; Holst, J.J. Gastric emptying, gastric secretion and enterogastrone response after administration of milk proteins or their peptide hydrolysates in humans. Eur. J. Nutr. 2004, 43, 127–139. [Google Scholar] [CrossRef]
- Hall, W.L.; Millward, D.J.; Long, S.J.; Morgan, L.M. Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite. Br. J. Nutr. 2003, 89, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Reidelberger, R.; Haver, A.; Chelikani, P.K. Role of peptide YY(3–36) in the satiety produced by gastric delivery of macronutrients in rats. Am. J. Physiol. Metab. 2013, 304, E944–E950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, R.C.; Singh, A.; Chelikani, P.K. Peptide YY mediates the satiety effects of diets enriched with whey protein fractions in male rats. FASEB J. 2018, 32, 850–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Khoury, D.; Vien, S.; Sanchez-Hernandez, D.; Kung, B.; Wright, A.; Goff, H.D.; Anderson, G.H. Increased milk protein content and whey-to-casein ratio in milk served with breakfast cereal reduce postprandial glycemia in healthy adults: An examination of mechanisms of action. J. Dairy Sci. 2019, 102, 6766–6780. [Google Scholar] [CrossRef]
- Hoefle, A.S.; Bangert, A.M.; Stamfort, A.; Gedrich, K.; Rist, M.J.; Lee, Y.-M.; Skurk, T.; Daniel, H. Metabolic Responses of Healthy or Prediabetic Adults to Bovine Whey Protein and Sodium Caseinate Do Not Differ. J. Nutr. 2015, 145, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Blouet, C.; Jo, Y.-H.; Li, X.; Schwartz, G.J. Mediobasal Hypothalamic Leucine Sensing Regulates Food Intake through Activation of a Hypothalamus-Brainstem Circuit. J. Neurosci. 2009, 29, 8302–8311. [Google Scholar] [CrossRef]
- Gartner, S.N.; Aidney, F.; Klockars, A.; Prosser, C.; Carpenter, E.A.; Isgrove, K.; Levine, A.S.; Olszewski, P.K. Intragastric preloads of l-tryptophan reduce ingestive behavior via oxytocinergic neural mechanisms in male mice. Appetite 2018, 125, 278–286. [Google Scholar] [CrossRef]
- Gietzen, D.; Leung, P.; Rogers, Q. Dietary amino acid imbalance and neurochemical changes in three hypothalamic areas. Physiol. Behav. 1989, 46, 503–511. [Google Scholar] [CrossRef]
- Heeley, N.; Kirwan, P.; Darwish, T.; Arnaud, M.; Evans, M.L.; Merkle, F.T.; Reimann, F.; Gribble, F.M.; Blouet, C. Rapid sensing of l -leucine by human and murine hypothalamic neurons: Neurochemical and mechanistic insights. Mol. Metab. 2018, 10, 14–27. [Google Scholar] [CrossRef]
- Choi, S.; DiSilvio, B.; Fernstrom, M.H.; Fernstrom, J.D. Meal ingestion, amino acids and brain neurotransmitters: Effects of dietary protein source on serotonin and catecholamine synthesis rates. Physiol. Behav. 2009, 98, 156–162. [Google Scholar] [CrossRef]
- Andreoli, M.F.; Stoker, C.; Lazzarino, G.P.; Canesini, G.; Luque, E.H.; Ramos, J.G. Dietary whey reduces energy intake and alters hypothalamic gene expression in obese phyto-oestrogen-deprived male rats. Br. J. Nutr. 2016, 116, 1125–1133. [Google Scholar] [CrossRef] [Green Version]
- Nilaweera, K.N.; Cabrera-Rubio, R.; Speakman, J.R.; O’Connor, P.M.; McAuliffe, A.; Guinane, C.M.; Lawton, E.M.; Crispie, F.; Aguilera, M.; Stanley, M.; et al. Whey protein effects on energy balance link the intestinal mechanisms of energy absorption with adiposity and hypothalamic neuropeptide gene expression. Am. J. Physiol. Metab. 2017, 313, E1–E11. [Google Scholar] [CrossRef] [PubMed]
- Potier, M.; Fromentin, G.; Calvez, J.; Benamouzig, R.; Martin-Rouas, C.; Pichon, L.; Tomé, D.; Marsset-Baglieri, A. A high-protein, moderate-energy, regular cheesy snack is energetically compensated in human subjects. Br. J. Nutr. 2009, 102, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.C.; Radavelli-Bagatini, S.; Hagger, M.S.; Ellis, V. Comparative effects of whey and casein proteins on satiety in overweight and obese individuals: A randomized controlled trial. Eur. J. Clin. Nutr. 2014, 68, 980–986. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Pezeshki, A.; Zapata, R.C.; Yee, N.J.; Knight, C.G.; Tuor, U.I.; Chelikani, P.K. Diets enriched in whey or casein improve energy balance and prevent morbidity and renal damage in salt-loaded and high-fat-fed spontaneously hypertensive stroke-prone rats. J. Nutr. Biochem. 2016, 37, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Holst, J.J.; Björck, I.M. Metabolic effects of amino acid mixtures and whey protein in healthy subjects: Studies using glucose-equivalent drinks. Am. J. Clin. Nutr. 2007, 85, 996–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezeshki, A.; Fahim, A.; Chelikani, P.K. Dietary Whey and Casein Differentially Affect Energy Balance, Gut Hormones, Glucose Metabolism, and Taste Preference in Diet-Induced Obese Rats. J. Nutr. 2015, 145, 2236–2244. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, P.K.; Billington, C.J.; Levine, A.S. Fos expression in feeding-related brain areas following intracerebroventricular administration of orphanin FQ in rats. Brain Res. 2000, 855, 171–175. [Google Scholar] [CrossRef]
- Beckman, T.R.; Shi, Q.; Levine, A.S.; Billington, C.J. Amygdalar opioids modulate hypothalamic melanocortin-induced anorexia. Physiol. Behav. 2009, 96, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, P.K.; Grace, M.K.; Fard, S.S.; Le Grevès, M.; Klockars, A.; Massi, M.; Schiöth, H.B.; Levine, A.S. Central nociceptin/orphanin FQ system elevates food consumption by both increasing energy intake and reducing aversive responsiveness. Am. J. Physiol. Integr. Comp. Physiol. 2010, 299, R655–R663. [Google Scholar] [CrossRef]
- Head, M.A.; Jewett, D.C.; Gartner, S.N.; Klockars, A.; Levine, A.S.; Olszewski, P.K. Effect of Oxytocin on Hunger Discrimination. Front. Endocrinol. 2019, 10, 297. [Google Scholar] [CrossRef]
- Roth, E.; Benoit, S.; Quentin, B.; Lam, B.; Will, S.; Ma, M.; Heeley, N.; Darwish, T.; Shrestha, Y.; Gribble, F.; et al. Behavioural and neurochemical mechanisms underpinning the feeding-suppressive effect of GLP-1/CCK combinatorial therapy. Mol. Metab. 2021, 43, 101118. [Google Scholar] [CrossRef]
- Yamamuro, K.; Bicks, L.K.; Leventhal, M.B.; Kato, D.; Im, S.; Flanigan, M.E.; Garkun, Y.; Norman, K.J.; Caro, K.; Sadahiro, M.; et al. A prefrontal–paraventricular thalamus circuit requires juvenile social experience to regulate adult sociability in mice. Nat. Neurosci. 2020, 23, 1240–1252. [Google Scholar] [CrossRef]
- James, J.C.; Funke, K. Repetitive transcranial magnetic stimulation reverses reduced excitability of rat visual cortex induced by dark rearing during early critical period. Dev. Neurobiol. 2020, 80, 399–410. [Google Scholar] [CrossRef]
- Delimaris, I. Adverse Effects Associated with Protein Intake above the Recommended Dietary Allowance for Adults. ISRN Nutr. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Soenen, S.; Westerterp-Plantenga, M.S. Proteins and satiety: Implications for weight management. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 747–751. [Google Scholar] [CrossRef] [Green Version]
- Fromentin, G.; Darcel, N.; Chaumontet, C.; Marsset-Baglieri, A.; Nadkarni, N.; Tomé, D. Peripheral and central mechanisms involved in the control of food intake by dietary amino acids and proteins. Nutr. Res. Rev. 2012, 25, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Meyer, B.; Al-Diab, D.; Vollmer, G.; Pischetsrieder, M. Mapping the glycoxidation product Nε-carboxymethyllysine in the milk proteome. Proteomics 2011, 11, 420–428. [Google Scholar] [CrossRef]
- Prosser, C.G.; Carpenter, E.A.; Hodgkinson, A.J. Nε-carboxymethyllysine in nutritional milk formulas for infants. Food Chem. 2019, 274, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Lönnerdal, B. Effects of Industrial Heating Processes of Milk-Based Enteral Formulas on Site-Specific Protein Modifications and Their Relationship to in Vitro and in Vivo Protein Digestibility. J. Agric. Food Chem. 2015, 63, 6787–6798. [Google Scholar] [CrossRef] [PubMed]
- Seiquer, I.; Rubio, L.A.; Peinado, M.J.; Delgado-Andrade, C.; Navarro, M.P. Maillard reaction products modulate gut microbiota composition in adolescents. Mol. Nutr. Food Res. 2014, 58, 1552–1560. [Google Scholar] [CrossRef]
- De Silva, A.; Bloom, S.R. Gut Hormones and Appetite Control: A Focus on PYY and GLP-1 as Therapeutic Targets in Obesity. Gut Liver 2012, 6, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Lam, D.D.; Garfield, A.S.; Marston, O.J.; Shaw, J.; Heisler, L.K. Brain serotonin system in the coordination of food intake and body weight. Pharmacol. Biochem. Behav. 2010, 97, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Jinno, S.; Yamazaki, K.; Nakamura, Y.; Kinouchi, T. Growth of term infants fed a commercial infant formula with a protein content of 2.2 g/100 kcal: An observational follow-up study. Biosci. Biotechnol. Biochem. 2020, 84, 633–639. [Google Scholar] [CrossRef]
- Semon, B.; Leung, P.; Rogers, Q.; Gietzen, D. Effect of type of protein on food intake of rats fed high protein diets. Physiol. Behav. 1987, 41, 451–458. [Google Scholar] [CrossRef]
- Stratford, J.M.; Thompson, J.A.; Finger, T.E. Immunocytochemical organization and sour taste activation in the rostral nucleus of the solitary tract of mice. J. Comp. Neurol. 2016, 525, 271–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travers, S.P. Quinine and citric acid elicit distinctive Fos-like immunoreactivity in the rat nucleus of the solitary tract. Am. J. Physiol. Integr. Comp. Physiol. 2002, 282, R1798–R1810. [Google Scholar] [CrossRef] [Green Version]
- Harrer, M.I.; Travers, S.P. Topographic organization of Fos-like immunoreactivity in the rostral nucleus of the solitary tract evoked by gustatory stimulation with sucrose and quinine. Brain Res. 1996, 711, 125–137. [Google Scholar] [CrossRef]
- King, C.T.; Travers, S.P.; Rowland, N.E.; Garcea, M.; Spector, A.C. Glossopharyngeal Nerve Transection Eliminates Quinine-Stimulated Fos-Like Immunoreactivity in the Nucleus of the Solitary Tract: Implications for a Functional Topography of Gustatory Nerve Input in Rats. J. Neurosci. 1999, 19, 3107–3121. [Google Scholar] [CrossRef]
- Ter Horst, G.; De Boer, P.; Luiten, P.; Van Willigen, J. Ascending projections from the solitary tract nucleus to the hypothalamus. A Phaseolus vulgaris lectin tracing study in the rat. Neuroscience 1989, 31, 785–797. [Google Scholar] [CrossRef]
- Rinaman, L. Ascending projections from the caudal visceral nucleus of the solitary tract to brain regions involved in food intake and energy expenditure. Brain Res. 2010, 1350, 18–34. [Google Scholar] [CrossRef] [Green Version]
- Stanstrup, J.; Schou, S.S.; Holmer-Jensen, J.; Hermansen, K.; Dragsted, L.O. Whey Protein Delays Gastric Emptying and Suppresses Plasma Fatty Acids and Their Metabolites Compared to Casein, Gluten, and Fish Protein. J. Proteome Res. 2014, 13, 2396–2408. [Google Scholar] [CrossRef] [PubMed]
- Hume, C.; Sabatier, N.; Menzies, J. High-Sugar, but Not High-Fat, Food Activates Supraoptic Nucleus Neurons in the Male Rat. Endocrinology 2017, 158, 2200–2211. [Google Scholar] [CrossRef]
- Kang, L.; Routh, V.H.; Kuzhikandathil, E.V.; Gaspers, L.D.; Levin, B.E. Physiological and molecular characteristics of rat hypothalamic ventromedial nucleus glucosensing neurons. Diabetes 2004, 53, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, B.; Prabhakar, E. Effects of body weight loss and taste on VMH-LH electrical activity of rats. Physiol. Behav. 1992, 52, 1187–1192. [Google Scholar] [CrossRef]
- Hodgkinson, A.J.; Wallace, O.A.; Smolenski, G.; Prosser, C.G. Gastric digestion of cow and goat milk: Peptides derived from simulated conditions of infant digestion. Food Chem. 2019, 276, 619–625. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| kJ | Protein (g) | Whey Protein (%) | Fat (g) | Carbohydrate (Lactose, g) | |
|---|---|---|---|---|---|
| Control (20:80) | 278.1 | 1.3 | 20.0 | 3.5 | 7.5 |
| 60:40 | 275.5 | 1.4 | 60.0 | 3.5 | 7.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wood, E.L.; Christian, D.G.; Arafat, M.; McColl, L.K.; Prosser, C.G.; Carpenter, E.A.; Levine, A.S.; Klockars, A.; Olszewski, P.K. Adjustment of Whey:Casein Ratio from 20:80 to 60:40 in Milk Formulation Affects Food Intake and Brainstem and Hypothalamic Neuronal Activation and Gene Expression in Laboratory Mice. Foods 2021, 10, 658. https://doi.org/10.3390/foods10030658
Wood EL, Christian DG, Arafat M, McColl LK, Prosser CG, Carpenter EA, Levine AS, Klockars A, Olszewski PK. Adjustment of Whey:Casein Ratio from 20:80 to 60:40 in Milk Formulation Affects Food Intake and Brainstem and Hypothalamic Neuronal Activation and Gene Expression in Laboratory Mice. Foods. 2021; 10(3):658. https://doi.org/10.3390/foods10030658
Chicago/Turabian StyleWood, Erin L., David G. Christian, Mohammed Arafat, Laura K. McColl, Colin G. Prosser, Elizabeth A. Carpenter, Allen S. Levine, Anica Klockars, and Pawel K. Olszewski. 2021. "Adjustment of Whey:Casein Ratio from 20:80 to 60:40 in Milk Formulation Affects Food Intake and Brainstem and Hypothalamic Neuronal Activation and Gene Expression in Laboratory Mice" Foods 10, no. 3: 658. https://doi.org/10.3390/foods10030658