
Antioxidant Activity and Bio-Accessibility of Polyphenols in Black Carrot (Daucus carota L. ssp. sativus var. atrorubens Alef.) and Two Derived Products during Simulated Gastrointestinal Digestion and Colonic Fermentation

 , , ,
, , ,  , ,
, ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Materials and Sample Preparation
2.3. In Vitro Gastrointestinal Digestion
2.4. In Vitro Colonic Fermentation
2.5. Extraction of Polyphenols from Digested Samples and from Faecal Incubates
2.6. Antioxidant Capacity Measurements
2.6.1. ABST Assay
2.6.2. DPPH Assay
2.6.3. Oxygen Radical Absorbance Capacity (ORAC) Assay
2.7. UHPLC-HRMS Polyphenol Analysis
2.7.1. Analysis of Flavonoids
2.7.2. Analysis of Anthocyanins
2.7.3. Identification and Quantification of Flavonoids and Anthocyanins
2.8. Statistical Analysis
3. Results and Discussion
3.1. Effect of In Vitro Digestion of Black Carrot and Its Derived Products on Polyphenol Stability and Antioxidant Capacity
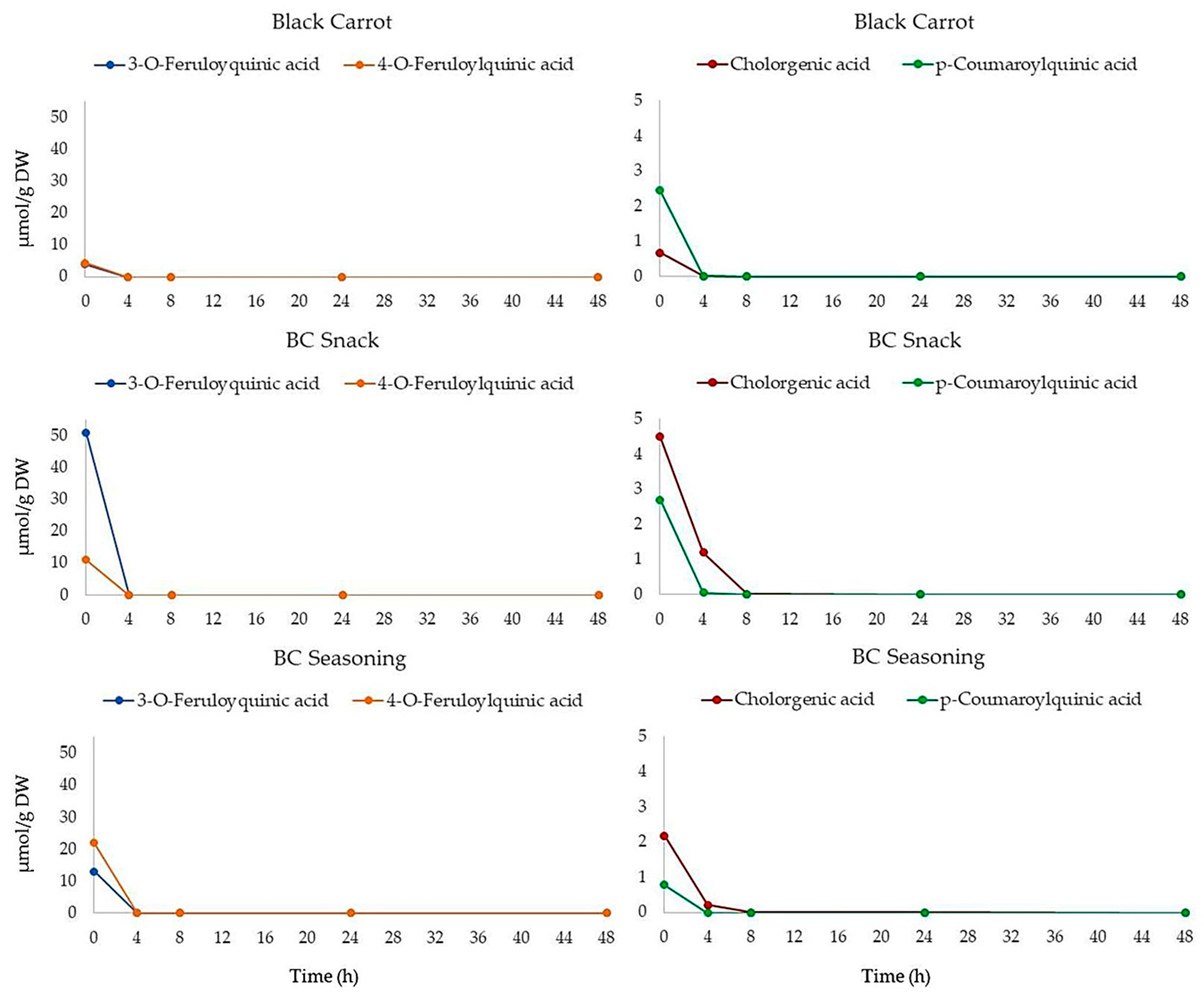
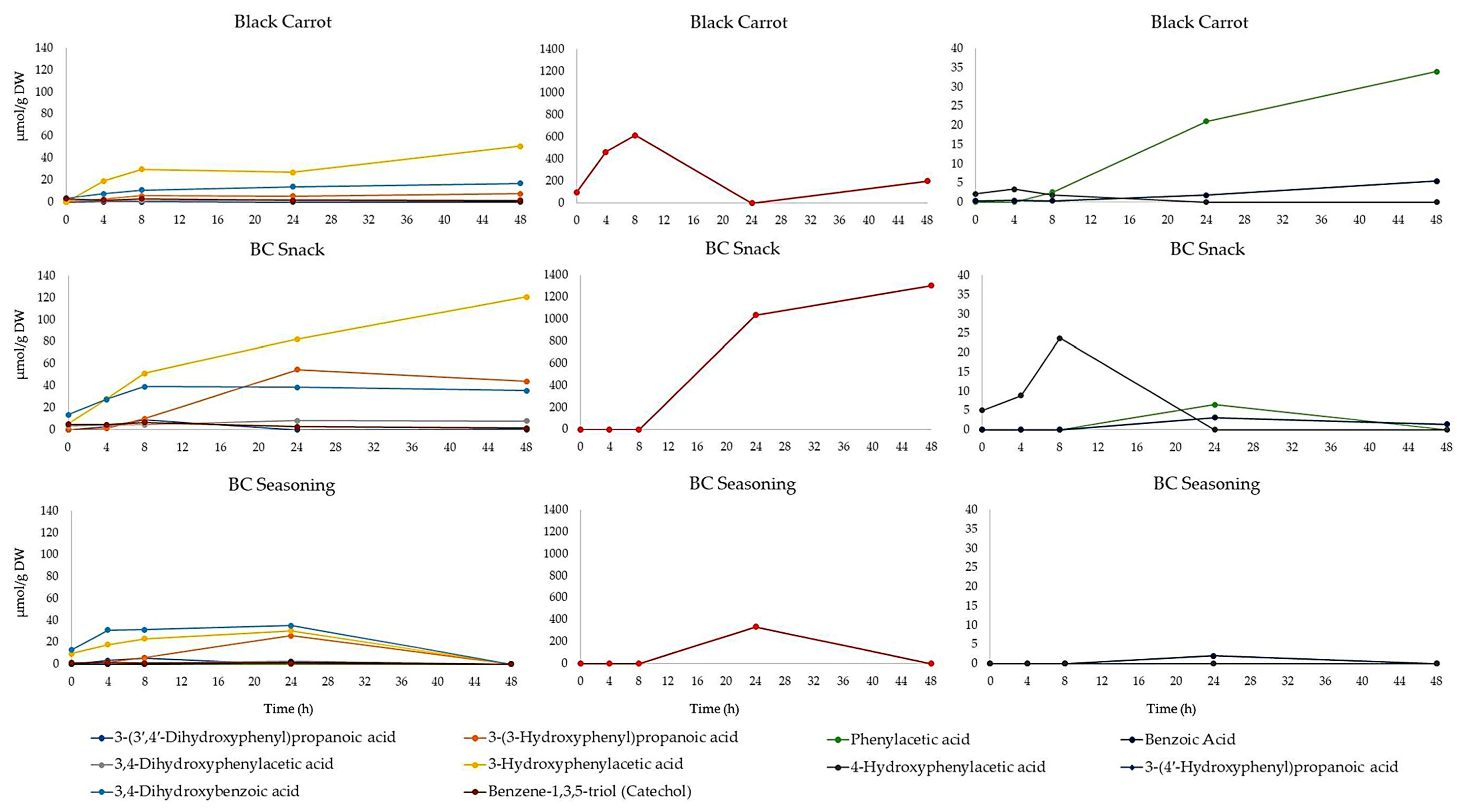
3.2. Effect of Colonic Fermentation on Polyphenol Stability in Black Carrot and Its Derived Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kamiloglu, S.; Van Camp, J.; Capanoglu, E. Black carrot polyphenols: Effect of processing. storage and digestion—An overview. Phytochem. Rev. 2018, 17, 379–395. [Google Scholar] [CrossRef]
- Ahmad, T.; Cawood, M.; Iqbal, Q.; Ariño, A.; Batool, A.; Sabir Tariq, R.M.; Azam, M.; Akhtar, S. Phytochemicals in daucus carota and their health benefits—Review article. Foods 2019, 8, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, S.; Rauf, A.; Imran, M.; Qamar, M.; Riaz, M.; Mubarak, M.S. Black carrot (Daucus carota L.). dietary and health promoting perspectives of its polyphenols: A review. Trends Food Sci. Technol. 2017, 66, 36–47. [Google Scholar] [CrossRef]
- Yoo, K.S.; Bang, H.; Pike, L.; Patil, B.S.; Lee, E.J. Comparing carotene. anthocyanins. and terpenoid concentrations in selected carrot lines of different colors. Hortic. Environ. Biotechnol. 2020, 61, 385–393. [Google Scholar] [CrossRef]
- Kumar, M.; Dahuja, A.; Sachdev, A.; Kaur, C.; Varghese, E.; Saha, S.; Sairam, K.V.S.S. Black Carrot (Daucus carota ssp.) and Black Soybean (Glycine max (L.) Merr.) Anthocyanin Extract: A Remedy to Enhance Stability and Functionality of Fruit Juices by Copigmentation. Waste Biomass Valorization 2020, 11, 99–108. [Google Scholar] [CrossRef]
- Gajewski, M.; Szymczak, P.; Dabrowska, A.; Kret, A.; Elkner, K.; Danilcenko, H. Some aspects of nutritive and biological value of carrot cultivars with orange. yellow and purple-coloured roots. Veg. Crop. Res. Bull. 2007, 67, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Kammerer, D.; Carle, R.; Schieber, A. Characterization of phenolic acids in black carrots (Daucus carota ssp. sativus var. atrorubens Alef.) by high-performance liquid chromatography/ electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 1331–1340. [Google Scholar] [CrossRef]
- Montilla, E.C.; Arzaba, M.R.; Hillebrand, S.; Winterhalter, P. Anthocyanin composition of black carrot (Daucus carota ssp. sativus var. atrorubens Alef.) Cultivars antonina. beta sweet. deep purple. and purple haze. J. Agric. Food Chem. 2011, 59, 3385–3390. [Google Scholar] [CrossRef]
- Schwarz, M.; Wray, V.; Winterhalter, P. Isolation and identification of novel pyranoanthocyanins from black carrot (Daucus carota L.) juice. J. Agric. Food Chem. 2004, 52, 5095–5101. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Acosta, G.; Ramos-Jacques, A.L.; Molina, G.A.; Maya-Cornejo, J.; Esparza, R.; Hernandez-Martinez, A.R.; Sánchez-González, I.; Estevez, M. Stability analysis of anthocyanins using alcoholic extracts from black carrot (Daucus carota ssp. Sativus var. Atrorubens alef.). Molecules 2018, 23, 2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizgier, P.; Kucharska, A.Z.; Sokół-Łetowska, A.; Kolniak-Ostek, J.; Kidoń, M.; Fecka, I. Characterization of phenolic compounds and antioxidant and anti-inflammatory properties of red cabbage and purple carrot extracts. J. Funct. Foods 2016, 21, 133–146. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Stintzing, A.C.; Carle, B.; Frei, B.; Wrolstad, R.E. Color and antioxidant properties of cyanidin-based anthocyanin pigments. J. Agric. Food Chem. 2002, 50, 6172–6181. [Google Scholar] [CrossRef]
- Algarra, M.; Fernandes, A.; Mateus, N.; de Freitas, V.; Esteves da Silva, J.C.G.; Casado, J. Anthocyanin profile and antioxidant capacity of black carrots (Daucus carota L. ssp. sativus var. atrorubens Alef.) from Cuevas Bajas. Spain. J. Food Compos. Anal. 2014, 33, 71–76. [Google Scholar] [CrossRef]
- Kammerer, D.; Carle, R.; Schieber, A. Detection of peonidin and pelargonidin glycosides in black carrots (Daucus carota ssp. sativus var. atrorubens Alef.) by high-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2407–2412. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Howell, K.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF/MS characterisation of phenolic acids and flavonoids in polyphenol-rich fruits and vegetables and their potential antioxidant activities. Antioxidants 2019, 8, 405. [Google Scholar] [CrossRef] [Green Version]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of in vitro digestion on composition. bioaccessibility and antioxidant activity of food polyphenols—A non-systematic review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry. bioavailability and effects on health. Nat. Prod. Rep. 2009. [Google Scholar] [CrossRef]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frígola, A. Analytical methods for determining bioavailability and bioaccessibility of bioactive compounds from fruits and vegetables: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Moreno-Rojas, J.M.; Brindani, N.; Del Rio, D.; Lean, M.E.J.; Hara, Y.; Crozier, A. Bioavailability of black tea theaflavins: Absorption. metabolism. and colonic catabolism. J. Agric. Food Chem. 2017, 65, 5365–5374. [Google Scholar] [CrossRef]
- De Santiago, E.; Pereira-Caro, G.; Moreno-Rojas, J.M.; Cid, C.; De Peña, M.P. Digestibility of (poly)phenols and antioxidant activity in raw and cooked cactus cladodes (Opuntia ficus-indica). J. Agric. Food Chem. 2018, 66, 5832–5844. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Pasli, A.A.; Ozcelik, B.; Van Camp, J.; Capanoglu, E. Colour retention. anthocyanin stability and antioxidant capacity in black carrot (Daucus carota) jams and marmalades: Effect of processing. storage conditions and in vitro gastrointestinal digestion. J. Funct. Foods 2015, 13, 1–10. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Capanoglu, E.; Bilen, F.D.; Gonzales, G.B.; Grootaert, C.; Van De Wiele, T.; Van Camp, J. Bioaccessibility of polyphenols from plant-processing byproducts of black carrot (Daucus carota L.). J. Agric. Food Chem. 2016, 64, 2450–2458. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, C.; Kamiloglu, S.; Grootaert, C.; Van Camp, J.; Hendrickx, M. Co-Ingestion of black carrot and strawberry. Effects on anthocyanin stability. bioaccessibility and u ptake. Foods 2020, 9, 1595. [Google Scholar] [CrossRef]
- Gu, C.; Suleria, H.A.R.; Dunshea, F.R.; Howell, K. Dietary lipids influence bioaccessibility of polyphenols from black carrots and affect microbial diversity under simulated gastrointestinal digestion. Antioxidants 2020, 9, 762. [Google Scholar] [CrossRef]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of many colors provide basic nutrition and bioavailable phytochemicals acting as a functional food. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Day, L.; Seymour, R.B.; Pitts, K.F.; Konczak, I.; Lundin, L. Incorporation of functional ingredients into foods. Trends Food Sci. Technol. 2009, 20, 388–395. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Pasli, A.A.; Ozcelik, B.; Van Camp, J.; Capanoglu, E. Influence of different processing and storage conditions on in vitro bioaccessibility of polyphenols in black carrot jams and marmalades. Food Chem. 2015, 186, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Değirmencioğlu, N.; Gurbuz, O.; Şahan, Y. The monitoring. via an in vitro digestion system. of the bioactive content of vegetable juice fermented with Saccharomyces cerevisiae and Saccharomyces boulardii. J. Food Process. Preserv. 2016, 40, 798–811. [Google Scholar] [CrossRef]
- Kay, C.D.; Clifford, M.N.; Mena, P.; McDougall, G.J.; Andres-Lacueva, C.; Cassidy, A.; Del Rio, D.; Kuhnert, N.; Manach, C.; Pereira-Caro, G.; et al. Recommendations for standardizing nomenclature for dietary (poly)phenol catabolites. Am. J. Clin. Nutr. 2020, 112, 1051–1068. [Google Scholar] [CrossRef]
- Moreno-Ortega, A.; Pereira-Caro, G.; Ordóñez, J.L.; Moreno-Rojas, R.; Ortíz-Somovilla, V.; Moreno-rojas, J.M. Bioaccessibility of bioactive compounds of ‘fresh garlic’ and ‘black garlic’ through in vitro gastrointestinal digestion. Foods 2020, 9, 1582. [Google Scholar] [CrossRef]
- Ordoñez-Díaz, J.L.; Moreno-Ortega, A.; Roldán-Guerra, F.J.; Ortíz-Somovilla, V.; Moreno-Rojas, J.M.; Pereira-Caro, G. In Vitro Gastrointestinal Digestion and Colonic Catabolism of Mango (Mangifera indica L.) Pulp Polyphenols. Foods 2020, 9, 1836. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Madrona, A.; Bravo, L.; Espartero, J.L.; Alcudia, F.; Cert, A.; Mateos, R. Antioxidant activity evaluation of alkyl hydroxytyrosyl ethers, a new class of hydroxytyrosol derivatives. Food Chem. 2009, 115, 86–91. [Google Scholar] [CrossRef]
- Sánchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Hornedo-Ortega, R.; Álvarez-Fernández, M.A.; Cerezo, A.B.; Garcia-Garcia, I.; Troncoso, A.M.; Garcia-Parrilla, M.C. Influence of fermentation process on the anthocyanin composition of wine and vinegar elaborated from strawberry. J. Food Sci. 2017, 82, 364–372. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goldagre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemicals analysis chemical analysis Chemical Analylsis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Mannino, G.; Di Stefano, V.; Laurita, A.; Pitonzo, R.; Gentile, C. Vaccinium macrocarpon (Cranberry)-based dietary supplements: Variation in mass uniformity, proanthocyanidin dosage and anthocyanidin profile demonstrates quality control standard needed. Nutrients 2020, 12, 992. [Google Scholar] [CrossRef] [Green Version]
- Witrowa-Rajchert, D.; Rząca, M. Effect of drying method on the microstructure and physical properties of dried apples. Dry. Technol. 2009, 27, 903–909. [Google Scholar] [CrossRef]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays. A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Craft, B.D.; Kerrihard, A.L.; Amarowicz, R.; Pegg, R.B. Phenol-based antioxidants and the in vitro methods used for ther assessment. Compr. Rev. Food Sci. Food Saf. 2012, 11, 148–173. [Google Scholar] [CrossRef]
- Correa-Betanzo, J.; Allen-Vercoe, E.; McDonald, J.; Schroeter, K.; Corredig, M.; Paliyath, G. Stability and biological activity of wild blueberry (Vaccinium angustifolium) polyphenols during simulated in vitro gastrointestinal digestion. Food Chem. 2014, 165, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Wu, X.; Zhao, T.; Zhao, J.; Li, F.; Zou, Y.; Yang, L. In vitro bioaccessibility and antioxidant activity of anthocyanins from mulberry (Morus atropurpurea Roxb.) following simulated gastro-intestinal digestion. Food Res. Int. 2012, 46, 76–82. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Capanoglu, E. Investigating the in vitro bioaccessibility of polyphenols in fresh and sun-dried figs (F icus carica L.). Int. J. Food Sci. Technol. 2013, 48, 2621–2629. [Google Scholar] [CrossRef]
- Toydemir, G.; Capanoglu, E.; Kamiloglu, S.; Boyacioglu, D.; De Vos, R.C.; Hall, R.D.; Beekwilder, J. Changes in sour cherry (Prunus cerasus L.) antioxidants during nectar processing and in vitro gastrointestinal digestion. J. Funct. Foods 2013, 5, 1402–1413. [Google Scholar] [CrossRef]
- Netzel, M.; Netzel, G.; Kammerer, D.R.; Schieber, A.; Carle, R.; Simons, L.; Konczak, I. Cancer cell antiproliferation activity and metabolism of black carrot anthocyanins. Innov. Food Sci. Emerg. Technol. 2017, 8, 365–372. [Google Scholar] [CrossRef]
- Harada, K.; Kano, M.; Takayanagi, T.; Yamakawa, O.; Ishikawa, F. Absorption of acylated anthocyanins in rats and humans after ingesting an extract of Ipomoea batatas purple sweet potato tuber. Biosci. Biotechnol. Biochem. 2004, 68, 1500–1507. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Mena, P.; Calani, L.; Borges, G.; Pereira-Caro, G.; Bresciani, L.; Del Rio, D.; Lean, M.E.; Crozier, A. New insights into the bioavailability of red raspberry anthocyanins and ellagitannins. Free Radic. Biol. Med. 2015, 89, 758–769. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, I.L.; Dragsted, L.O.; Ravn-Haren, G.; Freese, R.; Rasmussen, S.E. Absorption and secretion of blackcurrant anthocyanins in humans and Watanabe heritable hyperlipidemic rabbits. J. Agric. Food Chem. 2003, 51, 2813–2820. [Google Scholar] [CrossRef]
- Wu, X.L.; Cao, G.H.; Prior, R.L. Absorption and metabolism of anthocyanins in elderly women after consumption of elderberry or blueberry. J. Nutr. 2002, 132, 1865–1871. [Google Scholar] [CrossRef]
- Parada, J.; Aguilera, J.M. Food microstructure affects the bioavailability of several nutrients. J. Food Sci. 2007, 72, R21–R32. [Google Scholar] [CrossRef] [PubMed]
- Couteau, D.; McCartney, A.L.; Gibson, G.R.; Williamson, G.; Faulds, C.B. Isolation and characterization of human colonic bacteria able to hydrolyse chlorogenic acid. J. Appl. Microbiol. 2001, 90, 873–881. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 4-Hydroxybenzoic Acid | 3,4-Dihydroxybenzoic Acid | p-Coumaric Acid | Caffeic Acid | Chlorogenic Acid | 3-O-Feruloylquinic Acid | 4-O-Feruloylquinic Acid | Ferulic Acid | p-Coumaroyl Quinic Acid | Total | |
|---|---|---|---|---|---|---|---|---|---|---|
| Black Carrot | ||||||||||
| Non-digested | 0.13 ± 0.02 c | 0.22 ± 0.04 d | 0.01 ± 0.01 b | 0.02 ± 0.01 c | 2.52 ± 0.08 a | 4.01 ± 0.02 a | 0.06 ± 0.01 c | 0.04 ± 0.01 b | 4,41 ± 0.8 a | 11.4 ± 0.3 a |
| Oral Digestion | 0.37 ± 0.01 a | 0.44 ± 0.05 c | 0.03 ± 0.01 a | 0.09 ± 0.01 a | 0.39 ± 0.03 c | 3.4 ± 0.7 b | n.d. | 0.05 ± 0.01 a | 1.93 ± 0.02 c | 6.7 ± 0.8 b |
| % | 297 | 199 | 342 | 435 | 15.4 | 85 | 0 | 118 | 44 | 58.6 |
| Gastric Digestion | 0.27 ± 0.01 b | 0.65 ± 0.01 b | 0.02 ± 0.01 b | 0.03 ± 0.01 b | 0.55 ± 0.06 b | 2.5 ± 0.4 c | 0.25 ± 0.06 b | 0.03 ± 0.01 c | 1.9 ± 0.7 c | 6.3 ± 0.6 b |
| % | 212 | 297 | 190 | 135 | 21.8 | 63 | 405 | 65 | 44 | 54.8 |
| Intestinal Digestion | 0.33 ± 0.01 a | 0.73 ± 0.02 a | 0.02 ± 0.00 b | 0.04 ± 0.00 b | 0.68 ± 0.03 b | 4.08 ± 0.04 a | 4.5 ± 0.2 a | 0.05 ± 0.01 a | 2.44 ± 0.03 b | 12.9 ± 0.3 a |
| % | 259 | 333 | 285 | 218 | 26.9 | 102 | 7231 | 136 | 55 | 113 |
| Black Carrot Snack | ||||||||||
| Non-digested | 0.87 ± 0.05 c | 3.2 ± 0.1 a | 0.03 ± 0.01 a | 0.27 ± 0.04 a | 6.6 ± 0.2 a | 81.3 ± 0.7 a | 11.6 ± 0.8 b | 0.56 ± 0.04 a | 3.34 ± 0.03 a | 108 ± 2 a |
| Oral Digestion | 1.30 ± 0.09 b | 3.0 ± 0.2 a | 0.02 ± 0.00 a | 0.25 ± 0.02 b | 5.3 ± 0.4 b | 60 ± 11 b | 9 ± 1 c | 0.43 ± 0.09 c | 2.6 ± 0.6 b | 82 ± 13 b |
| % | 149 | 92 | 74 | 90 | 80 | 74 | 79 | 77 | 79 | 76 |
| Gastric Digestion | 2.0 ± 0.1 a | 2.8 ± 0.0 b | n.d. | 0.2 ± 0.0 c | 4.6 ± 0.1 c | 60 ± 4 b | 13 ± 1 a | 0.5 ± 0.1 b | 3.1 ± 0.1 a | 85 ± 5 b |
| % | 230 | 87.6 | 0 | 91 | 69 | 74 | 109 | 81 | 93 | 80 |
| Intestinal Digestion | 1.8 ± 0.0 a | 2.8 ± 0.1 b | n.d. | 0.2 ± 0.0 c | 4.5 ± 0.1 c | 51 ± 2 c | 11.1 ± 0.5 b | 0.5 ± 0.0 b | 2.7 ± 0.8 b | 74 ± 3 c |
| % | 206 | 86 | 0 | 83 | 68 | 62 | 95 | 92 | 82 | 69 |
| Black Carrot Seasoning | ||||||||||
| Non-digested | 2.14 ± 0.05 a | 2.8 ± 0.2 a | 0.01 ± 0.00 | 0.05 ± 0.01 | 3.80 ± 0.08 a | 28 ± 4 a | 13.4 ± 0.8 b | 0.34 ± 0.07 a | 1.98 ± 0.05 a | 53 ±2 a |
| Oral Digestion | 1.8 ± 0.1 b | 2.4 ± 0.1 b | 0.01 ± 0.00 | 0.05 ± 0.01 | 3.1 ± 0.8 b | 21 ± 7 b | 9.0 ± 0.5 c | 0.22 ± 0.01 b | 1.4 ± 0.5 b | 39 ± 9 b |
| % | 82 | 86 | 100 | 100 | 83 | 76 | 67 | 66 | 72 | 75 |
| Gastric Digestion | 1.9 ± 0.6 b | 2.7 ± 0.4 a b | n.d. | n.d. | 2.3 ± 0.4 c | 17 ± 6 c | 13 ± 1 b | 0.2 ± 0.0 b | 1.1 ± 0.4 c | 38 ± 9 b |
| % | 88.6 | 97 | 0 | 0 | 60 | 62 | 94 | 58 | 56 | 73 |
| Intestinal Digestion | 1.9 ± 0.3 b | 2.4 ± 0.5 b | 0.01 ± 0.00 | 0.05 ± 0.01 | 2.2 ± 0.1 c | 13 ± 1 d | 22 ± 8 a | 0.11 ± 0.01 c | 0.79 ± 0.05 d | 43 ±10 ab |
| % | 89 | 87 | 100 | 100 | 58 | 47 | 164 | 32 | 40 | 81 |
| Cyanidin-3-xyl-gal | Cyanidin-3-xyl-coumaroyl-glu-gal | Cyanidin-3-xyl-feruloyl-glu-gal | Cyanidin-3-xyl-sinapoyl-glu-gal | Pelargonidin-3-sambiburoside | Pelargonidin-3,5-diglu | Cyanidin-3-xyl-glu-gal | Delphinidin-3-glu | Total | |
|---|---|---|---|---|---|---|---|---|---|
| Black Carrot | |||||||||
| Non-digested | 7 ± 1 | 15 ± 1 a | 134 ± 11 a | 35.7 ± 0.9 a | 0.15 ± 0.01 | 0.14 ± 0.01 | 1.8 ± 0.5 | 0.05 ± 0.01 | 195 ± 15 a |
| Oral Digestion | n.d. | 0.05 ± 0.01 b | 0.62 ± 0.05 b | 0.06 ± 0.01 b | n.d. | n.d. | n.d. | n.d. | 0.7 ± 0.1 b |
| % | 0 | 0.3 | 0.5 | 0.2 | 0 | 0 | 0 | 0 | 0.4 |
| Gastric Digestion | n.d. | 0.009 ± 0.001 c | 0.15 ± 0.02 c | 0.009 ± 0.001 c | n.d. | n.d. | n.d. | n.d. | 0.2 ± 0.0 c |
| % | 0 | 0.1 | 0.1 | 0.05 | 0 | 0 | 0 | 0 | 0.1 |
| Intestinal Digestion | n.d. | 0.07 ± 0.02 b | 0.9 ± 0.1 b | 0.05 ± 0.02 b | n.d. | n.d. | n.d. | n.d. | 1.1 ± 0.2 b |
| % | 0 | 0.4 | 0.7 | 0.1 | 0 | 0 | 0 | 0 | 0.5 |
| Black Carrot Snack | |||||||||
| Non-digested | 0.28 ± 0.02 a | 0.78 ± 0.07 a | 4.76 ± 0.01 a | 1.94 ± 0.01 a | 0.009 ± 0.001 b | 0.023 ± 0.001 b | 0.376 ± 0.002 a | 0.02 ± 0.01 a | 8.2 ± 0.1 a |
| Oral Digestion | 0.16 ± 0.01 b | 0.54 ± 0.03 b | 3.6 ± 0.2 b | 1.5 ± 0.1 b | 0.012 ± 0.001 a | 0.01 ±0.01 a | 0.21 ± 0.02 b | 0.01 ± 0.01 b | 6.1 ± 0.5 b |
| % | 59 | 70 | 77 | 78 | 133 | 61 | 57 | 46 | 75 |
| Gastric Digestion | 0.07 ± 0.02 c | 0.24 ± 0.02 c | 1.7 ± 0.2 c | 0.71 ± 0.02 c | n.d. | n.d. | 0.17 ± 0.04 c | n.d. | 2.9 ± 0.3 c |
| % | 27 | 31 | 36 | 37 | 0 | 0 | 45 | 0 | 36 |
| Intestinal Digestion | 0.036 ± 0.001 d | 0.27 ± 0.04 c | 2.2 ± 0.2 c | 0.83 ± 0.05 c | n.d. | n.d. | 0.15 ± 0.02 c | n.d. | 3.5 ± 0.3 c |
| % | 13 | 35 | 47 | 43 | 0 | 0 | 41 | 0 | 43 |
| Black Carrot Seasoning | |||||||||
| Non-digested | 0.009 ± 0.010 b | 0.30 ± 0.02 a | 3.0 ± 0.5 b | 1.6 ± 0.4 a | n.d. | n.d. | 0.06 ± 0.02 | 0.09 ± 0.01 | 5 ± 1 a |
| Oral Digestion | 0.03 ± 0.02 a | 0.26 ± 0.01 b | 3.7 ± 0.3 a | 1.6 ± 0.2 a | n.d. | n.d. | n.d. | n.d. | 5.7 ± 0.5 a |
| % | 34 | 88 | 124 | 101 | 0 | 0 | 0 | 0 | 109 |
| Gastric Digestion | 0.02 ± 0.01 a | 0.14 ± 0.01 c | 1.86 ± 0.01 c | 0.80 ± 0.08 b | n.d. | n.d. | n.d. | n.d. | 2.8 ± 0.1 b |
| % | 21 | 48 | 62 | 48 | 0 | 0 | 0 | 0 | 54 |
| Intestinal Digestion | 0.009 ± 0.001 b | 0.14 ± 0.01 c | 1.9 ± 0.1 c | 0.84 ± 0.02 b | n.d. | n.d. | n.d. | n.d. | 2.9 ± 0.1 b |
| % | 10 | 46 | 65 | 51 | 0 | 0 | 0 | 0 | 57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira-Caro, G.; Ordóñez-Díaz, J.L.; de Santiago, E.; Moreno-Ortega, A.; Cáceres-Jiménez, S.; Sánchez-Parra, M.; Roldán-Guerra, F.J.; Ortiz-Somovilla, V.; Moreno-Rojas, J.M. Antioxidant Activity and Bio-Accessibility of Polyphenols in Black Carrot (Daucus carota L. ssp. sativus var. atrorubens Alef.) and Two Derived Products during Simulated Gastrointestinal Digestion and Colonic Fermentation. Foods 2021, 10, 457. https://doi.org/10.3390/foods10020457
Pereira-Caro G, Ordóñez-Díaz JL, de Santiago E, Moreno-Ortega A, Cáceres-Jiménez S, Sánchez-Parra M, Roldán-Guerra FJ, Ortiz-Somovilla V, Moreno-Rojas JM. Antioxidant Activity and Bio-Accessibility of Polyphenols in Black Carrot (Daucus carota L. ssp. sativus var. atrorubens Alef.) and Two Derived Products during Simulated Gastrointestinal Digestion and Colonic Fermentation. Foods. 2021; 10(2):457. https://doi.org/10.3390/foods10020457
Chicago/Turabian StylePereira-Caro, Gema, José Luis Ordóñez-Díaz, Elsy de Santiago, Alicia Moreno-Ortega, Salud Cáceres-Jiménez, Mónica Sánchez-Parra, Francisco Javier Roldán-Guerra, Víctor Ortiz-Somovilla, and José Manuel Moreno-Rojas. 2021. "Antioxidant Activity and Bio-Accessibility of Polyphenols in Black Carrot (Daucus carota L. ssp. sativus var. atrorubens Alef.) and Two Derived Products during Simulated Gastrointestinal Digestion and Colonic Fermentation" Foods 10, no. 2: 457. https://doi.org/10.3390/foods10020457