
Interaction between Flavonoids and Carotenoids on Ameliorating Oxidative Stress and Cellular Uptake in Different Cells


Abstract
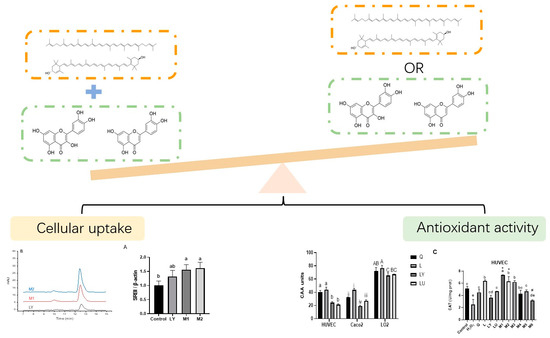
:
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Cell Culture and Treatments
2.3. Cell Viability Assay
2.4. Cellular Antioxidant Activity
2.5. Enzyme Activities of SOD, GSH-Px, and CAT
2.6. Measurement of Lycopene and Lutein Uptake in HUVEC, Caco-2, and L-02 Cells
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. Effects of Flavonoids and Carotenoids on the Oxidative-Induced Cell Damage
3.2. Interactions on Cellular Antioxidant Activity
3.3. Interactions of Phytochemicals on SOD, GSH-Px, and CAT Activity
3.4. Flavonoids Affect the Uptake of Carotenoids
3.5. The Presence of Flavonoids Influenced the Expression of Carotenoid Transporters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lampe, J.W. Health effects of vegetables and fruit: Assessing mechanisms of action in human experimental studies. Am. J. Clin. Nutr. 1999, 70, 475S–490S. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ouyang, Y.; Liu, J.; Zhu, M.; Zhao, G.; Bao, W.; Hu, F.B. Fruit and vegetable consumption and mortality from all causes, cardiovascular disease, and cancer: Systematic review and dose-response meta-analysis of prospective cohort studies. Bmj 2014, 349, g4490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517s–520s. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, M.A.T.; Paterson, J.; Bucknall, M.; Arcot, J. Interactions between phytochemicals from fruits and vegetables: Effects on bioactivities and bioavailability. Crit. Rev. Food Sci. Nutr. 2018, 58, 1310–1329. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, H.; Zhang, B.; Deng, Z. The synergistic and antagonistic antioxidant interactions of dietary phytochemical combinations. Crit. Rev. Food Sci. Nutr. 2021, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Durak, A.; Gawlik-Dziki, U.; Kowlska, I. Coffee with ginger—Interactions of biologically active phytochemicals in the model system. Food Chem. 2015, 166, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Enko, J.; Gliszczynska-Swiglo, A. Influence of the interactions between tea (Camellia sinensis) extracts and ascorbic acid on their antioxidant activity: Analysis with interaction indexes and isobolograms. Food Addit. Contam. Part A 2015, 32, 1234–1242. [Google Scholar] [CrossRef]
- Muhammad, D.R.A.; Praseptiangga, D.; Van de Walle, D.; Dewettinck, K. Interaction between natural antioxidants derived from cinnamon and cocoa in binary and complex mixtures. Food Chem. 2017, 231, 356–364. [Google Scholar] [CrossRef]
- Wang, S.A.; Zhu, F. Dietary antioxidant synergy in chemical and biological systems. Crit. Rev. Food Sci. Nutr. 2017, 57, 2343–2357. [Google Scholar] [CrossRef]
- Reboul, E.; Thap, S.; Tourniaire, F.; Andre, M.; Juhel, C.; Morange, S.; Amiot, M.J.; Lairon, D.; Borel, P. Differential effect of dietary antioxidant classes (carotenoids, polyphenols, vitamins C and E) on lutein absorption. Br. J. Nutr. 2007, 97, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Phan, M.A.T.; Bucknall, M.; Arcot, J. Interactive effects of β-carotene and anthocyanins on cellular uptake, antioxidant activity and anti-inflammatory activity in vitro and ex vivo. J. Funct. Foods 2018, 45, 129–137. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Current. Opinion in Food Science 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Olmedilla, B.; Granado, F.; Southon, S.; Wright, A.J.; Blanco, I.; Gil-Martinez, E.; Berg, H.; Corridan, B.; Roussel, A.M.; Chopra, M.; et al. Serum concentrations of carotenoids and vitamins A, E, and C in control subjects from five European countries. Br. J. Nutr. 2001, 85, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress-implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef]
- Borel, P.; Lietz, G.; Goncalves, A.; Szabo de Edelenyi, F.; Lecompte, S.; Curtis, P.; Goumidi, L.; Caslake, M.J.; Miles, E.A.; Packard, C.; et al. CD36 and SR-BI are involved in cellular uptake of provitamin A carotenoids by Caco-2 and HEK cells, and some of their genetic variants are associated with plasma concentrations of these micronutrients in humans. J. Nutr. 2013, 143, 448–456. [Google Scholar] [CrossRef] [Green Version]
- Biehler, E.; Hoffmann, L.; Krause, E.; Bohn, T. Divalent minerals decrease micellarization and uptake of carotenoids and digestion products into Caco-2 cells. J. Nutr. 2011, 141, 1769–1776. [Google Scholar] [CrossRef] [Green Version]
- Thompson, H.J.; Heimendinger, J.; Diker, A.; O’Neill, C.; Haegele, A.; Meinecke, B.; Wolfe, P.; Sedlacek, S.; Zhu, Z.J.; Jiang, W.Q. Dietary botanical diversity affects the reduction of oxidative biomarkers in women due to high vegetable and fruit intake. J. Nutr. 2006, 136, 2207–2212. [Google Scholar] [CrossRef] [Green Version]
- During, A.; Hussain, M.M.; Morel, D.W.; Harrison, E.H. Carotenoid uptake and secretion by CaCo-2 cells: Beta-carotene isomer selectivity and carotenoid interactions. J. Lipid Res. 2002, 43, 1086–1095. [Google Scholar] [CrossRef] [Green Version]
- Moussa, M.; Landrier, J.F.; Reboul, E.; Ghiringhelli, O.; Comera, C.; Collet, X.; Frohlich, K.; Bohm, V.; Borel, P. Lycopene absorption in human intestinal cells and in mice involves scavenger receptor class B type I but not Niemann-Pick C1-like 1. J. Nutr. 2008, 138, 1432–1436. [Google Scholar] [CrossRef] [Green Version]
- Liang, R.; Han, R.M.; Fu, L.M.; Ai, X.C.; Zhang, J.P.; Skibsted, L.H. Baicalin in radical scavenging and its synergistic effect with beta-carotene in antilipoxidation. J. Agric. Food Chem. 2009, 57, 7118–7124. [Google Scholar] [CrossRef]
- Li, H.; Deng, Z.; Liu, R.; Loewen, S.; Tsao, R. Bioaccessibility, in vitro antioxidant activities and in vivo anti-inflammatory activities of a purple tomato (Solanum lycopersicum L.). Food Chem. 2014, 159, 353–360. [Google Scholar] [CrossRef]
- Kaulmann, A.; Andre, C.M.; Schneider, Y.J.; Hoffmann, L.; Bohn, T. Carotenoid and polyphenol bioaccessibility and cellular uptake from plum and cabbage varieties. Food Chem. 2016, 197, 325–332. [Google Scholar] [CrossRef]
- Bando, N.; Muraki, N.; Murota, K.; Terao, J.; Yamanishi, R. Ingested quercetin but not rutin increases accumulation of hepatic beta-carotene in BALB/c mice. Mol. Nutr. Food Res. 2010, 54, S261–S267. [Google Scholar] [CrossRef]
- Reboul, E.; Thap, S.; Perrot, E.; Amiot, M.J.; Lairon, D.; Borel, P. Effect of the main dietary antioxidants (carotenoids, gamma-tocopherol, polyphenols, and vitamin C) on alpha-tocopherol absorption. Eur. J. Clin. Nutr. 2007, 61, 1167–1173. [Google Scholar] [CrossRef]
- Zhang, Z.; Nie, M.; Liu, C.; Jiang, N.; Liu, C.; Li, D. Citrus Flavanones Enhance beta-Carotene Uptake in Vitro Experiment Using Caco-2 Cell: Structure-Activity Relationship and Molecular Mechanisms. J. Agric. Food Chem. 2019, 67, 4280–4288. [Google Scholar] [CrossRef]
- Ferreira, A.L.A.; Yeum, K.-J.; Liu, C.; Smith, D.; Krinsky, N.I.; Wang, X.-D.; Russell, R.M. Tissue Distribution of Lycopene in Ferrets and Rats after Lycopene Supplementation. J. Nutr. 2000, 130, 1256–1260. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.; Li, Q.; Rubakhin, S.S.; Sweedler, J.V.; Smith, J.W.; Neuringer, M.; Kuchan, M.; Erdman, J.W., Jr. (13)C-lutein is differentially distributed in tissues of an adult female rhesus macaque following a single oral administration: A pilot study. Nutr. Res. 2019, 61, 102–108. [Google Scholar] [CrossRef]
- Phan, M.A.T.; Bucknall, M.P.; Arcot, J. Interferences of anthocyanins with the uptake of lycopene in Caco-2 cells, and their interactive effects on anti-oxidation and anti-inflammation in vitro and ex vivo. Food Chem. 2019, 276, 402–409. [Google Scholar] [CrossRef]
- Claudie, D.M.; Alexandrine, D.; Bertrand, C.; Franck, T.; Marie-Josephe, A. Citrus flavanones enhance carotenoid uptake by intestinal Caco-2 cells. Food Funct. 2013, 4, 1625–1631. [Google Scholar] [CrossRef]
- Crozier, A.; Del Rio, D.; Clifford, M.N. Bioavailability of dietary flavonoids and phenolic compounds. Mol. Aspects Med. 2010, 31, 446–467. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Deng, Z.-Y.; Chen, X.; Zhang, B.; Fan, Y.; Li, H. Synergistic antioxidant effects of phenolic acids and carotenes on H2O2-induced H9c2 cells: Role of cell membrane transporters. Food Chem. 2021, 341, 128000. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Tomas, M.; Ozdal, T.; Capanoglu, E. Effect of food matrix on the content and bioavailability of flavonoids. Trends Food Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Marques, M.C.; Hacke, A.; Neto, C.A.C.; Mariutti, L.R.B. Impact of phenolic compounds in the digestion and absorption of carotenoids. Curr. Opin. Food Sci. 2021, 39, 190–196. [Google Scholar] [CrossRef]
- Nie, M.; Zhang, Z.; Liu, C.; Li, D.; Huang, W.; Liu, C.; Jiang, N. Hesperetin and Hesperidin Improved β-Carotene Incorporation Efficiency, Intestinal Cell Uptake, and Retinoid Concentrations in Tissues. J. Agric. Food Chem. 2019, 67, 3363–3371. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combination | HUVEC Cells | Caco-2 Cells | L-02 Cells | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ratio | CAATheo (%) a | CAAExp (%) b | Effect c | CAATheo (%) | CAAExp (%) | Effect | CAATheo (%) | CAAExp (%) | Effect | |
| LUT-Q | 1:10 | 38.3 ± 2.71 | 43.1 ± 4.66 | Synergy d | 31.7 ± 0.97 | 41.8 ± 3.49 | Synergy | 71.8 ± 4.91 | 36.9 ± 2.83 | Antagonisme |
| 1:5 | 36.9 ± 2.60 | 37.1 ± 3.69 | Additivity f | 31.3 ± 1.02 | 40.4 ± 1.48 | Synergy | 68.1 ± 1.24 | 26.7 ± 2.46 | Antagonism | |
| 1:1 | 30.7 ± 2.12 | 36.3 ± 2.30 | Synergy | 29.6 ± 1.23 | 24.7 ± 2.60 | Antagonism | 69.7 ± 2.89 | 68.9 ± 4.15 | Additivity | |
| 5:1 | 24.5 ± 1.64 | 19.0 ± 1.30 | Antagonism | 28.0 ± 1.44 | 36.6 ± 3.88 | Synergy | 71.4 ± 4.54 | 59.1 ± 3.15 | Antagonism | |
| 10:1 | 23 ± 1.53 | 27.9 ± 1.97 | Synergy | 27.6 ± 1.49 | 34.1 ± 2.99 | Synergy | 71.8 ± 4.91 | 41.1 ± 3.31 | Antagonism | |
| LUT-L | 1:10 | 42.2 ± 3.48 | 53.7 ± 4.13 | Synergy | 42.3 ± 2.21 | 37.0 ± 1.75 | Antagonism | 75.8 ± 3.94 | 68.9 ± 1.30 | Antagonism |
| 1:5 | 40.4 ± 3.30 | 54.8 ± 3.65 | Synergy | 41.1 ± 2.15 | 50.7 ± 1.62 | Synergy | 68.8 ± 1.06 | 82.5 ± 6.61 | Synergy | |
| 1:1 | 32.8 ± 2.54 | 42.6 ± 3.80 | Synergy | 35.5 ± 1.91 | 34.8 ± 3.82 | Additivity | 71.9 ± 2.35 | 73.1 ± 50 | Additivity | |
| 5:1 | 25.2 ± 1.78 | 44.3 ± 4.66 | Synergy | 29.9 ± 1.67 | 18.2 ± 0.73 | Antagonism | 75.0 ± 3.64 | 63.7 ± 4.86 | Antagonism | |
| 10:1 | 23.4 ± 1.61 | 39.8 ± 3.46 | Synergy | 28.7 ± 1.61 | 34.8 ± 3.46 | Synergy | 75.78 ± 3.94 | 33.9 ± 2.15 | Antagonism | |
| LYP-Q | 1:10 | 38.6 ± 2.73 | 46.4 ± 3.65 | Synergy | 31.0 ± 0.91 | 41.2 ± 2.63 | Synergy | 71.6 ± 5.27 | 80.5 ± 3.46 | Synergy |
| 1:5 | 37.4 ± 2.64 | 55.3 ± 2.24 | Synergy | 30.0 ± 0.92 | 42.0 ± 3.82 | Synergy | 66.8 ± 4.47 | 78.6 ± 2.96 | Synergy | |
| 1:1 | 32.2 ± 2.25 | 51.8 ± 5.70 | Synergy | 25.8 ± 0.93 | 35.4 ± 2.55 | Synergy | 68.9 ± 4.83 | 69.4 ± 4.46 | Additivity | |
| 5:1 | 27.0 ± 1.85 | 54.5 ± 5.89 | Synergy | 21.5 ± 0.94 | 41.1 ± 3.76 | Synergy | 71.1 ± 5.18 | 56.7 ± 2.35 | Antagonism | |
| 10:1 | 25.8 ± 1.76 | 42.5 ± 7.73 | Synergy | 20.6 ± 0.94 | 31.1 ± 3.30 | Synergy | 71.6 ± 5.27 | 66.7 ± 3.82 | Additivity | |
| LYP-L | 1:10 | 42.4 ± 3.50 | 47.0 ± 2.59 | Synergy | 41.6 ± 2.15 | 38.2 ± 1.71 | Antagonism | 75.6 ± 4.28 | 77.9 ± 1.79 | Additivity |
| 1:5 | 40.9 ± 3.34 | 44.7 ± 7.20 | Synergy | 39.8 ± 2.05 | 25.3 ± 1.82 | Antagonism | 67.5 ± 4.16 | 84.4 ± 5.54 | Synergy | |
| 1:1 | 34.3 ± 2.67 | 52.5 ± 2.58 | Synergy | 31.6 ± 1.61 | 26.3 ± 1.68 | Antagonism | 71.1 ± 4.21 | 71.1 ± 4.65 | Additivity | |
| 5:1 | 27.7 ± 1.99 | 52.1 ± 9.59 | Synergy | 23.5 ± 1.16 | 35.7 ± 2.46 | Synergy | 74.8 ± 4.26 | 59.2 ± 3.62 | Antagonism | |
| 10:1 | 26.2 ± 1.84 | 49.3 ± 9.78 | Synergy | 21.6 ± 1.06 | 27.6 ± 2.51 | Synergy | 75.6 ± 4.28 | 52.7 ± 2.62 | Antagonism | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Deng, Z.; Zheng, L.; Zhang, B.; Luo, T.; Li, H. Interaction between Flavonoids and Carotenoids on Ameliorating Oxidative Stress and Cellular Uptake in Different Cells. Foods 2021, 10, 3096. https://doi.org/10.3390/foods10123096
Chen X, Deng Z, Zheng L, Zhang B, Luo T, Li H. Interaction between Flavonoids and Carotenoids on Ameliorating Oxidative Stress and Cellular Uptake in Different Cells. Foods. 2021; 10(12):3096. https://doi.org/10.3390/foods10123096
Chicago/Turabian StyleChen, Xuan, Zeyuan Deng, Liufeng Zheng, Bing Zhang, Ting Luo, and Hongyan Li. 2021. "Interaction between Flavonoids and Carotenoids on Ameliorating Oxidative Stress and Cellular Uptake in Different Cells" Foods 10, no. 12: 3096. https://doi.org/10.3390/foods10123096