Potential Biological Activities of Peptides Generated during Casein Proteolysis by Curly Kale (Brassica oleracea L. var. sabellica L.) Leaf Extract: An In Silico Preliminary Study

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Curly Kale Leaf Extract
2.3. Hydrolysis of Casein by Curly Kale Extract
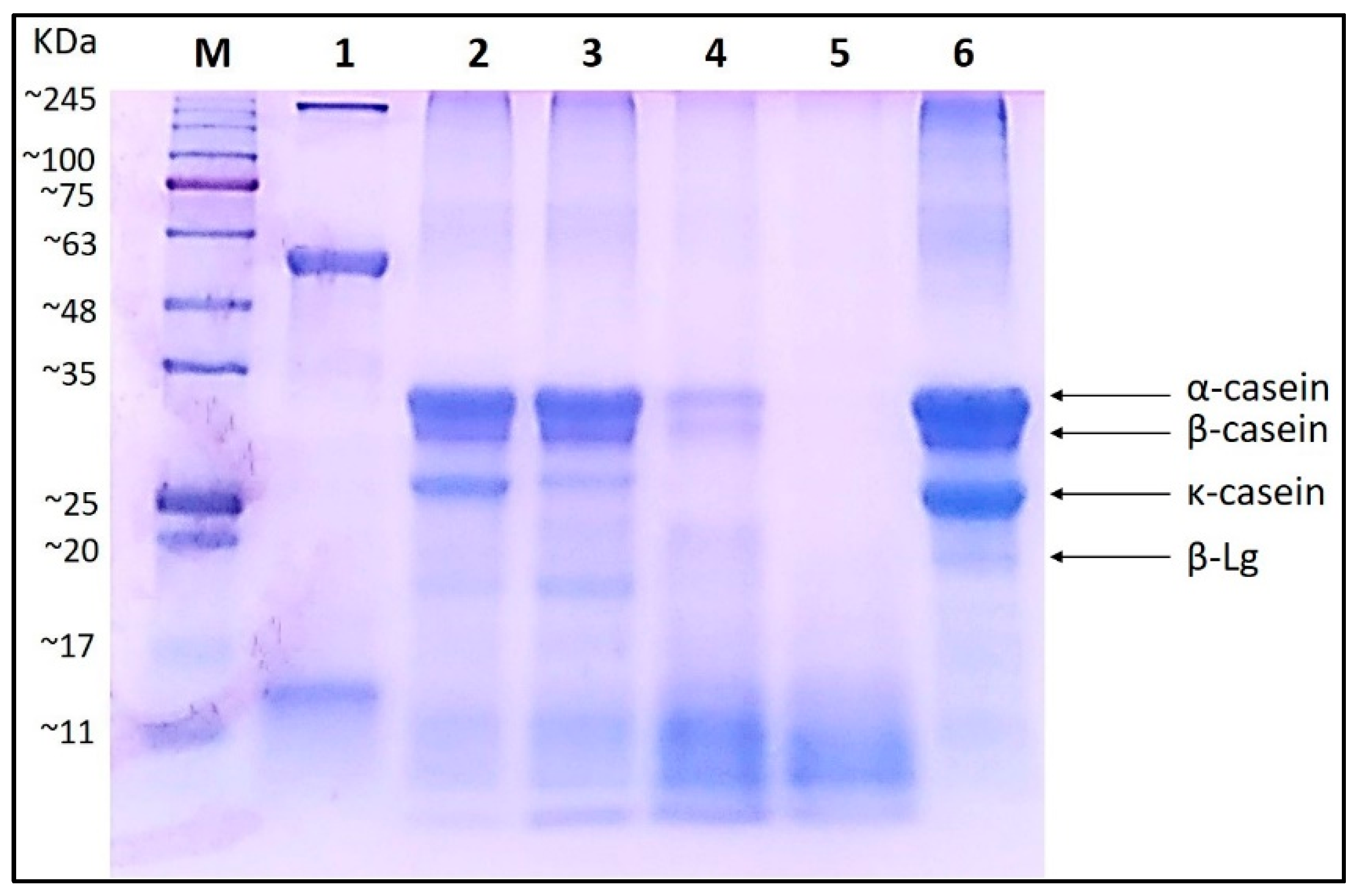
2.4. Sodium Dodecyl Sulfate–Polyacrylamide gel Electrophoresis (SDS–PAGE)
2.5. Determination of Peptide Concentration
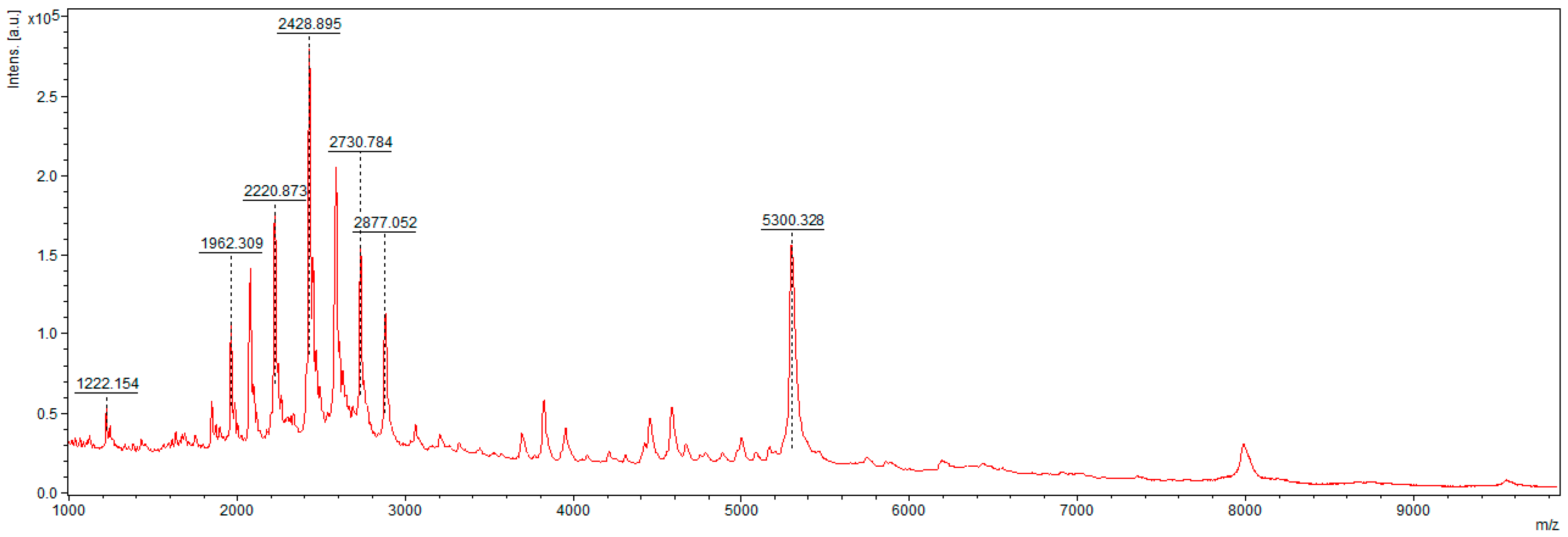
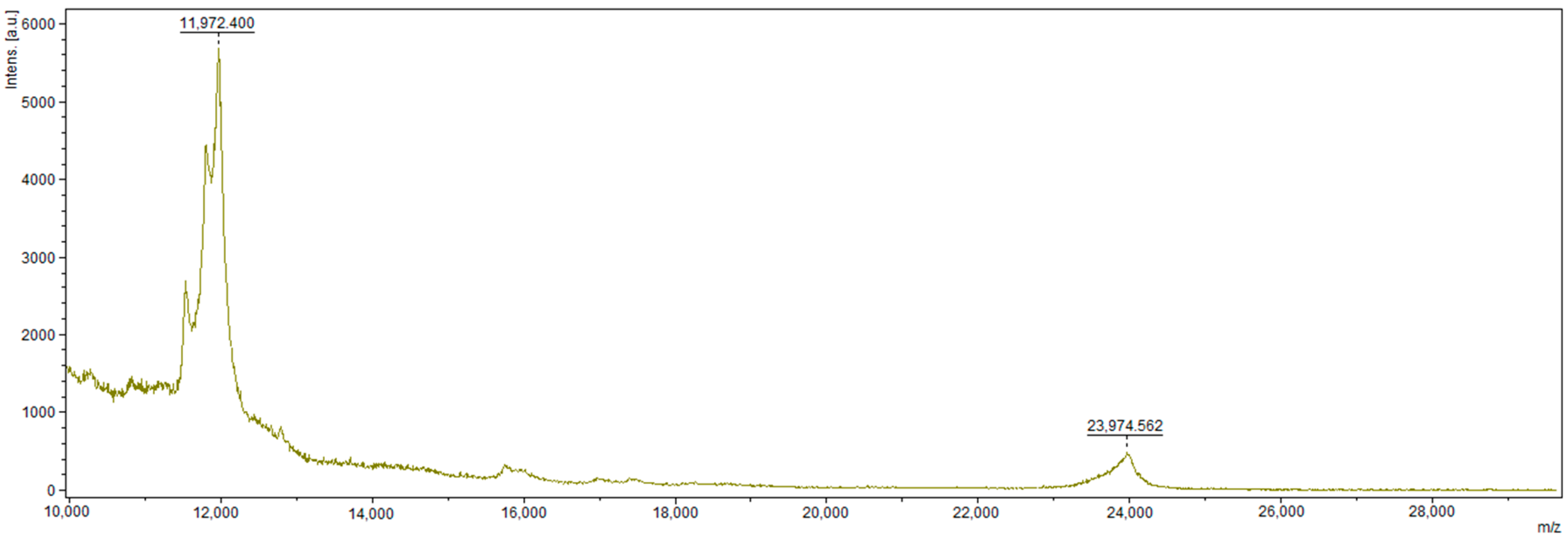
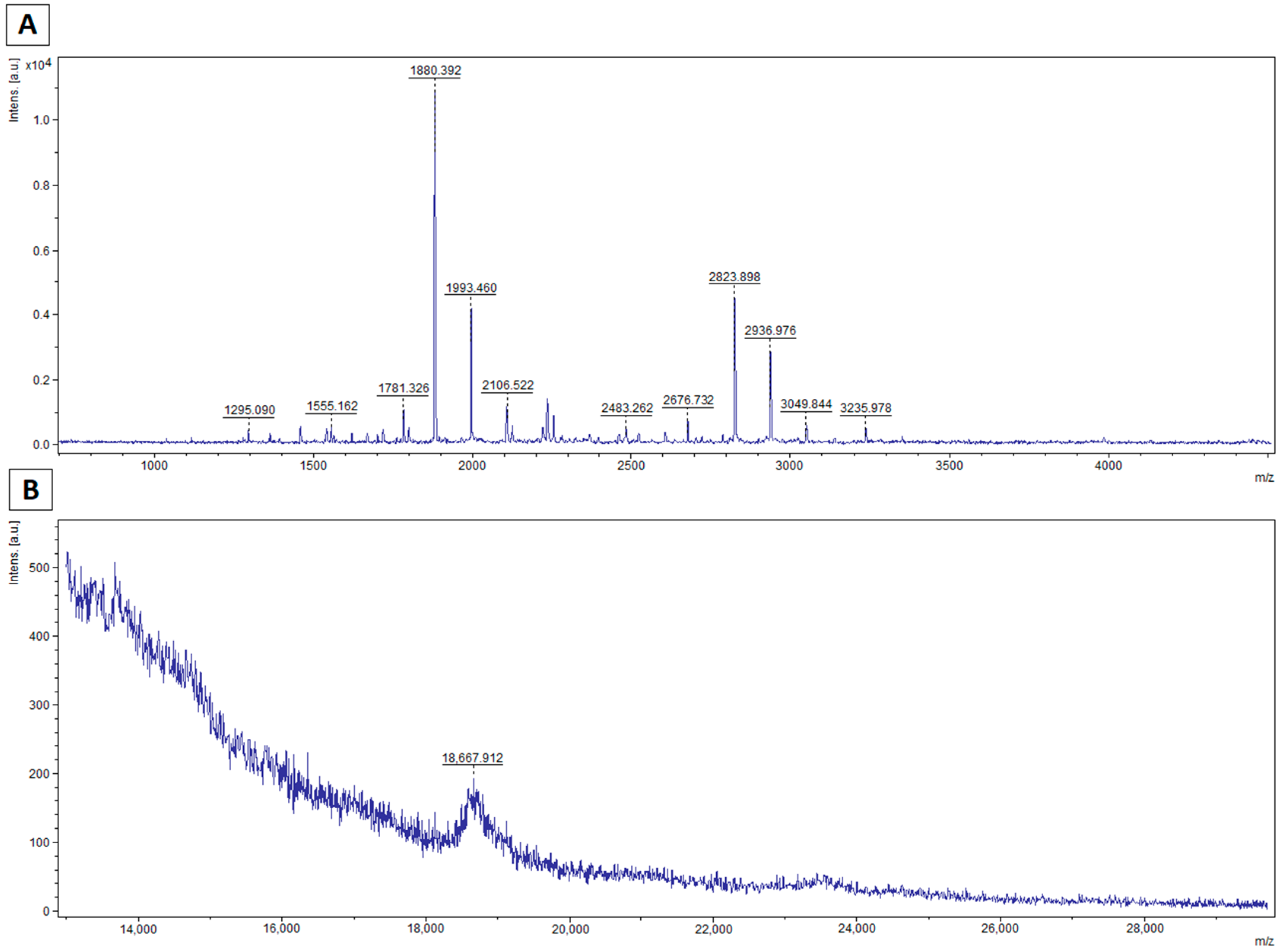
2.6. Mass Spectrometry Analysis
2.7. In Silico Analysis
3. Results and Discussion
3.1. Electrophoretic Pattern of Bovine Milk Casein Treated with Curly Kale Extract
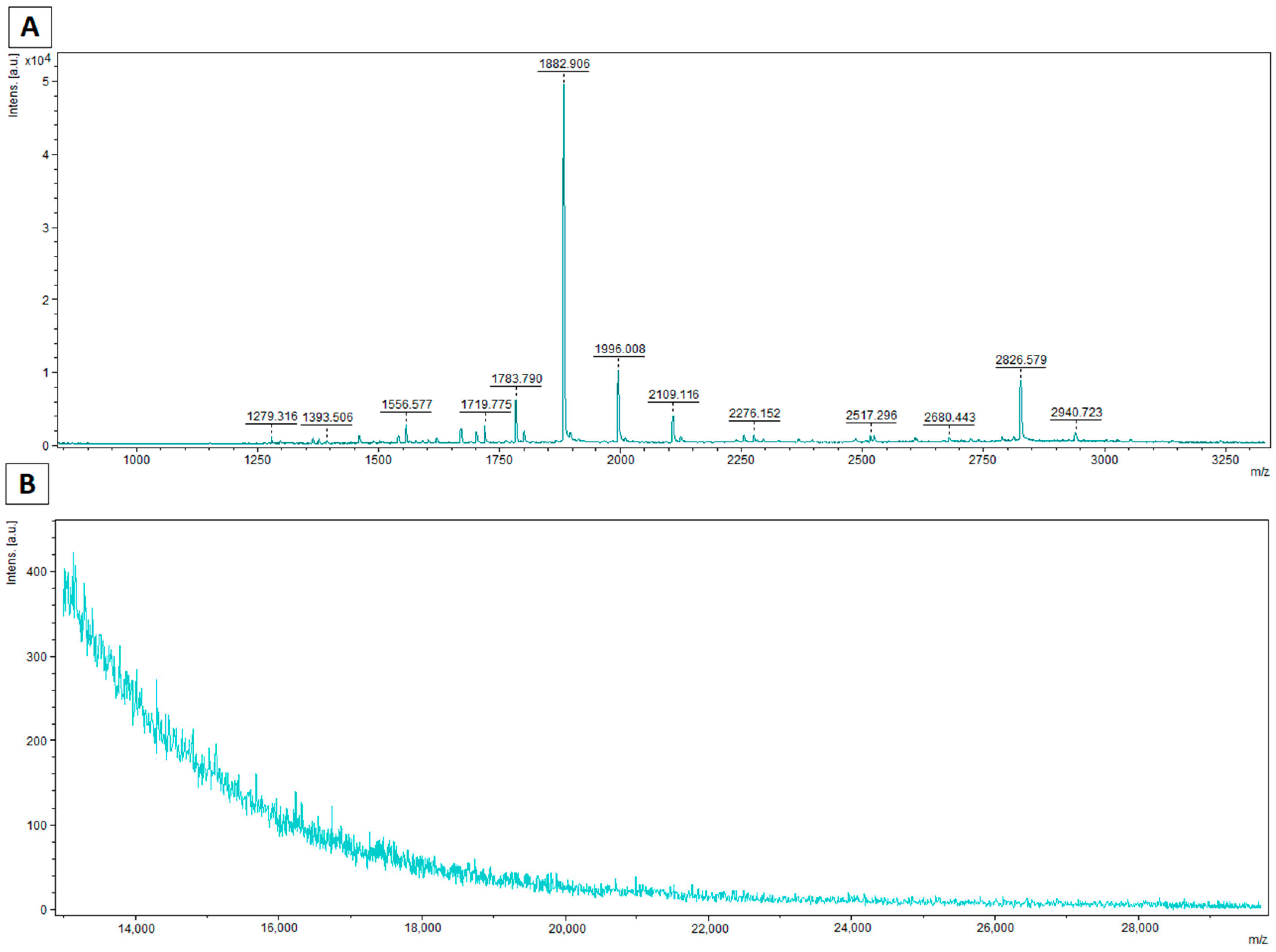
3.2. MALDI–TOF MS Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial Proteases Applications. Front. Bioeng. Biotechnol. 2019, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Scott, K.; Hemara, Y.; Zhang, H.; Otter, D. Purification and characterisation of a protease (tamarillin) from tamarillo fruit. Food Chem. 2018, 256, 228–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.A.; Mir, S.A.; Paray, M.A. Plant proteases as milk-clotting enzymes in cheesemaking: A review. Dairy Sci. Technol. 2013, 94, 5–16. [Google Scholar] [CrossRef]
- Ben Amira, A.; Besbes, S.; Attia, H.; Blecker, C. Milk-clotting properties of plant rennets and their enzymatic, rheological, and sensory role in cheese making: A review. Int. J. Food Prop. 2017, 20, S76–S93. [Google Scholar] [CrossRef] [Green Version]
- Galán, E.; Prados, F.; Pino, A.; Tejada, L.; Fernández-Salguero, J. Influence of different amounts of vegetable coagulant from cardoon Cynara cardunculus and calf rennet on the proteolysis and sensory characteristics of cheeses made with sheep milk. Int. Dairy J. 2008, 18, 93–98. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Ramírez-Suarez, J.C.; Yada, R.Y. Plant proteases for bioactive peptides release: A review. Crit. Rev. Food Sci. Nutr. 2017, 58, 2147–2163. [Google Scholar] [CrossRef]
- Tripathi, P.; Tomar, R.; Jagannadham, M.V. Purification and biochemical characterisation of a novel protease streblin. Food Chem. 2011, 125, 1005–1012. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, B.; Yan, Q.-J.; Jiang, Z.-Q. Comparative analysis on the distribution of protease activities among fruits and vegetable resources. Food Chem. 2016, 213, 708–713. [Google Scholar] [CrossRef]
- Michalak, M.; Skrzypczak, K.; Nastaj, M.; Terpiłowski, K.; Skrzypek, T.; Waśko, A.; Polak-Berecka, M. Possibility of Using Fermented Curly Kale Juice to Manufacture Feta-Type Cheese. Appl. Sci. 2020, 10, 4020. [Google Scholar] [CrossRef]
- Phelan, M.; Aherne, A.; FitzGerald, R.J.; O’Brien, N.M. Casein-derived bioactive peptides: Biological effects, industrial uses, safety aspects and regulatory status. Int. Dairy J. 2009, 19, 643–654. [Google Scholar] [CrossRef]
- Meisel, H.; FitzGerald, R.J. Biofunctional peptides from milk proteins: Mineral binding and cytomodulatory effects. Curr. Pharm. Des. 2003, 9, 1289–1295. [Google Scholar] [PubMed]
- Cavalli, S.V.; Lufrano, D.; Colombo, M.L.; Priolo, N. Properties and applications of phytepsins from thistle flowers. Phytochemistry 2013, 92, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Sandré, C.; Gleizes, A.; Forestier, F.; Gorges-Kergot, R.; Chilmonczyk, S.; Léonil, J.; Moreau, M.; Labarre, C. A Peptide Derived from Bovine b-Casein Modulates Functional Properties of Bone Marrow-Derived Macrophages from Germfree and Human Flora-Associated Mice. Nutr. Immunol. 2001, 131, 2936–2942. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.C.; Razaname, A.; Mutter, M.; Juillerat, M.A. Identification of Angiotensin-I-Converting Enzyme Inhibitory Pep-tides Derived from Sodium Caseinate Hydrolysates Produced by Lactobacillus helveticus NCC 2765. J. Agric. Food Chem. 2004, 52, 6923–6931. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Cheng, S.; Lu, W.; Du, M. Advancement and prospects of bioinformatics analysis for studying bioactive peptides from food-derived protein: Sequence, structure, and functions. TrAC Trends Anal. Chem. 2018, 105, 7–17. [Google Scholar] [CrossRef]
- Kęska, P.; Stadnik, J. Porcine myofibrillar proteins as potential precursors of bioactive peptides—An in silico study. Food Funct. 2016, 7, 2878–2885. [Google Scholar] [CrossRef] [PubMed]
- Bechaux, J.; Gatellier, P.; Le Page, J.-F.; Drillet, Y.; Sante-Lhoutellier, V. A comprehensive review of bioactive peptides obtained from animal byproducts and their applications. Food Funct. 2019, 10, 6244–6266. [Google Scholar] [CrossRef]
- FitzGerald, R.J.; Cermeno, M.; Khalesi, M.; Kleekayai, T.; Amigo-Benavent, M. Application of in silico approaches for the generation of milk protein-derived bioactive peptides. J. Funct. Foods 2020, 64, 103636. [Google Scholar] [CrossRef]
- Egito, A.S.; Miclo, L.; López, C.; Adam, A.; Girardet, J.M.; Gaillard, J.L. Separation and characterization of mares’ milk αs1-, β-, κ-caseins, gamma-casein-like, and proteose peptone component 5-like peptides. J. Dairy Sci. 2002, 85, 697–706. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Signor, L.; Erba, E.B. Matrix-assisted Laser Desorption/Ionization Time of Flight (MALDI-TOF) Mass Spectrometric Analysis of Intact Proteins Larger than 100 kDa. J. Vis. Exp. 2013, 79, e50635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bio-active peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef] [Green Version]
- Milk Bioactive Peptides Data Base. Available online: https://mbpdb.nws.oregonstate.edu/ (accessed on 20 May 2021).
- Panyayai, T.; Ngamphiw, C.; Tongsima, S.; Mhuantong, W.; Limsripraphan, W.; Choowongkomon, K.; Sawatdichaikul, O. FeptideDB: A web application for new bioactive peptides from food protein. Heliyon 2019, 5, e02076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Scott, K.; Hemar, Y.; Otter, D. Protease activity of enzyme extracts from tamarillo fruit and their specific hydrolysis of bovine caseins. Food Res. Int. 2018, 109, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Méndez, N.; Balderrama-Carmona, A.; García-Sandoval, S.E.; Ramírez-Vigil, P.; Leal-Ramos, M.Y.; García-Triana, A. Proteolysis and Rheological Properties of Cream Cheese Made with a Plant-Derived Coagulant from Solanum elaeagnifolium. Foods 2019, 8, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Xiao, C.; Zhang, H.; Ren, F.; Lei, X.; Yang, Z.; Yu, Z. Characterization and milk coagulating properties of Cynanchum otophyllum Schneid. proteases. J. Dairy Sci. 2018, 101, 2842–2850. [Google Scholar] [CrossRef] [PubMed]
- Anusha, R.; Singh, M.K.; Bindhu, O.S. Characterisation of potential milk coagulants from Calotropis gigantea plant parts and their hydrolytic pattern of bovine casein. Eur. Food Res. Technol. 2014, 238, 997–1006. [Google Scholar] [CrossRef]
- Di Luccia, A.; Picariello, G.; Trani, A.; Alviti, G.; Loizzo, P.; Faccia, M.; Addeo, F. Occurrence of β-casein fragments in cold-stored and curdled river buffalo (Bubalus bubalis L.) milk. J. Dairy Sci. 2009, 92, 1319–1329. [Google Scholar] [CrossRef] [Green Version]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Takano, T. Antihypertensive Effect of Sour Milk and Peptides Isolated from It That are Inhibitors to Angiotensin I-Converting Enzyme. J. Dairy Sci. 1995, 78, 1253–1257. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, A.; Yue, T.; Wang, J.; Wang, Y.; Su, J. Purification and identification of five novel antioxidant peptides from goat milk casein hydrolysates. J. Dairy Sci. 2013, 96, 4242–4251. [Google Scholar] [CrossRef] [PubMed]
- Corrons, M.A.; Liggieri, C.S.; Trejo, S.A.; Bruno, M.A. ACE-inhibitory peptides from bovine caseins released with peptidases from Maclura pomifera latex. Food Res. Int. 2017, 93, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Bueno-Gavilá, E.; Abellán, A.; Girón-Rodríguez, F.; Cayuela, J.M.; Salazar, E.; Gómez, R.; Tejada, L. Bioactivity of hydrolysates obtained from bovine casein using artichoke (Cynara scolymus L.) proteases. J. Dairy Sci. 2019, 102, 10711–10723. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mass (Da) | Peptide Sequence | Source of Peptide | ID of the Bioactive Peptide in the Database | Activity |
|---|---|---|---|---|
| 1151 | LGPVRGPFPIIV | β-casein | (3169, 8594, 8882, 8857, 8801, 8506) c | dipeptidyl peptidase IV inhibitor |
| (7512, 7619, 8160, 7508, 3502, 7545) c | ACE inhibitor | |||
| 8286 c | antioxidative | |||
| 2753 c | regulating the stomach mucosal membrane activity | |||
| 3283 c | antithrombotic | |||
| 3461 c | antiamnesic | |||
| 8325 c | glucose uptake stimulating | |||
| 1459 | PGPIPNSLPQNIPPLT | β-casein | (3169, 8824, 3180, 8875, 3170, 8638) c | dipeptidyl peptidase IV inhibitor |
| (7512, 7836, 7625, 7513)c | ACE inhibitor | |||
| (2753, 2756, 2754)c | regulating the stomach mucosal membrane activity | |||
| (3283, 3284, 3285) c | antithrombotic | |||
| (3461, 3459, 3460) c | antiamnesic | |||
| 1718 | YQEPVLGPVRGPFPIIV | β-casein | biopep01621 a | antihypertensive |
| biopep04091 a; P02666 b | antimicrobial | |||
| biopep04801 a; P02666 b | immunomodulatory and cytomodulatory | |||
| (3169, 8869, 8922, 8594, 8593, 8882) c | dipeptidyl peptidase IV inhibitor | |||
| P02666 b(7512, 7619, 8160, 7508, 3502, 7545) c | ACE inhibitor | |||
| (8286, 7878, 7879) c | antioxidative | |||
| 2753 c | regulating the stomach mucosal membrane activity | |||
| P02666 b; 3283 c | antithrombotic | |||
| 3461 c | antiamnesic | |||
| 8325 c | glucose uptake stimulating | |||
| 1881 | LYQEPVLGPVRGPFPIIV | β-casein | P02666 b | immunomodulatory |
| (3169, 8869, 8922, 8594, 8593 8857) c | dipeptidyl peptidase IV inhibitor | |||
| (7512, 7619, 3381, 8160, 7508, 3502) c | ACE inhibitor | |||
| (7872, 7879, 7878, 8286) c | antioxidative | |||
| 2753 c | regulating the stomach mucosal membrane activity | |||
| 8325 c | glucose uptake stimulating | |||
| 3283 c | antithrombotic | |||
| 3461 c | antiamnesic | |||
| 2281 | WMHQPHQPLPPTVMFPPQSVLS | β-casein | (3180, 8923, 8877, 8532, 8856, 8638) c | dipeptidyl peptidase IV inhibitor |
| (3385, 7836, 3502, 3391, 2664, 7513) c | ACE inhibitor | |||
| 2824 | LSQSKVLPVPQKAVPYPQRDMPIQAFL | β-casein | (3180, 8877, 8922, 3181, 3171, 8857) c | dipeptidyl peptidase IV inhibitor |
| (3373, 2653, 3370, 8951, 7583, 3666) c | ACE inhibitor | |||
| (8479, 7877, 7896, 7875, 7876, 8278) c | antioxidative | |||
| 3024 | LSLSQSKVLPVPQKAVPYPQRDMPIQAFLL | β-casein | (3182, 3180, 8877, 8867, 8922, 3181) c | dipeptidyl peptidase IV inhibitor |
| (3373, 2653, 3370, 8951, 7583, 3666) c | ACE inhibitor | |||
| (8479, 7877, 7896, 7875, 7876, 8278) c | antioxidative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polak-Berecka, M.; Michalak-Tomczyk, M.; Skrzypczak, K.; Michalak, K.; Rachwał, K.; Waśko, A. Potential Biological Activities of Peptides Generated during Casein Proteolysis by Curly Kale (Brassica oleracea L. var. sabellica L.) Leaf Extract: An In Silico Preliminary Study. Foods 2021, 10, 2877. https://doi.org/10.3390/foods10112877
Polak-Berecka M, Michalak-Tomczyk M, Skrzypczak K, Michalak K, Rachwał K, Waśko A. Potential Biological Activities of Peptides Generated during Casein Proteolysis by Curly Kale (Brassica oleracea L. var. sabellica L.) Leaf Extract: An In Silico Preliminary Study. Foods. 2021; 10(11):2877. https://doi.org/10.3390/foods10112877
Chicago/Turabian StylePolak-Berecka, Magdalena, Magdalena Michalak-Tomczyk, Katarzyna Skrzypczak, Katarzyna Michalak, Kamila Rachwał, and Adam Waśko. 2021. "Potential Biological Activities of Peptides Generated during Casein Proteolysis by Curly Kale (Brassica oleracea L. var. sabellica L.) Leaf Extract: An In Silico Preliminary Study" Foods 10, no. 11: 2877. https://doi.org/10.3390/foods10112877