
Influence of Plant Origins and Seasonal Variations on Nutritive Values, Phenolics and Antioxidant Activities of Adenia viridiflora Craib., an Endangered Species from Thailand

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Extraction
2.2. Determination of Nutritive Values
2.2.1. Moisture Content
2.2.2. Protein
2.2.3. Fat
2.2.4. Dietary Fiber
2.2.5. Sugars
2.2.6. Ash
2.2.7. Carbohydrate
2.2.8. Energy
2.2.9. Vitamin C
2.2.10. Carotenoids
2.2.11. Minerals
2.3. Determination of Total Phenolic Contents
2.4. Determination of Antioxidant Activities
2.5. Statistical Analysis
3. Results
3.1. Nutritive Values
3.2. Total Phenolic Contents and Total Flavonoid Contents
3.3. Antioxidant Activities
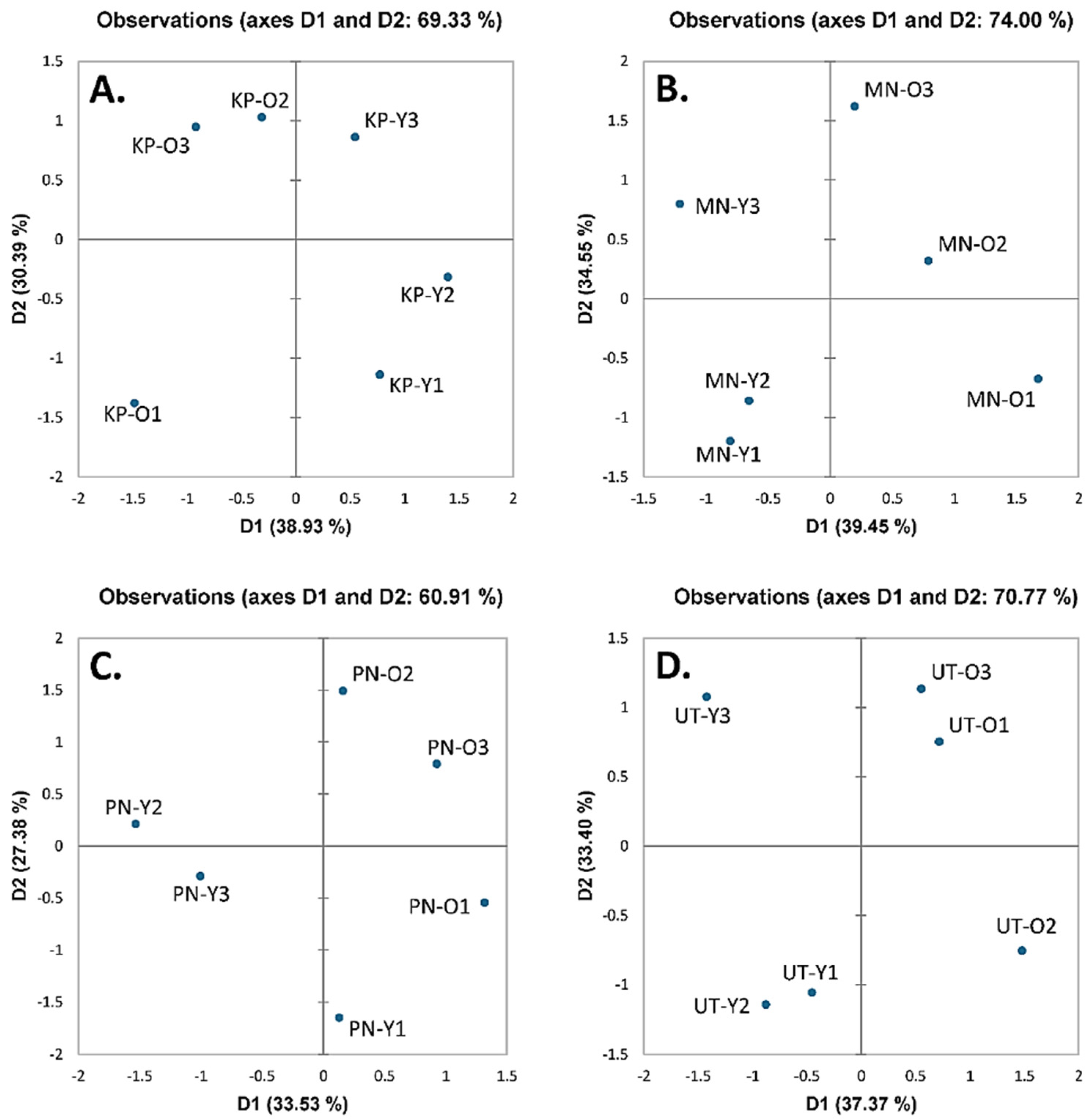
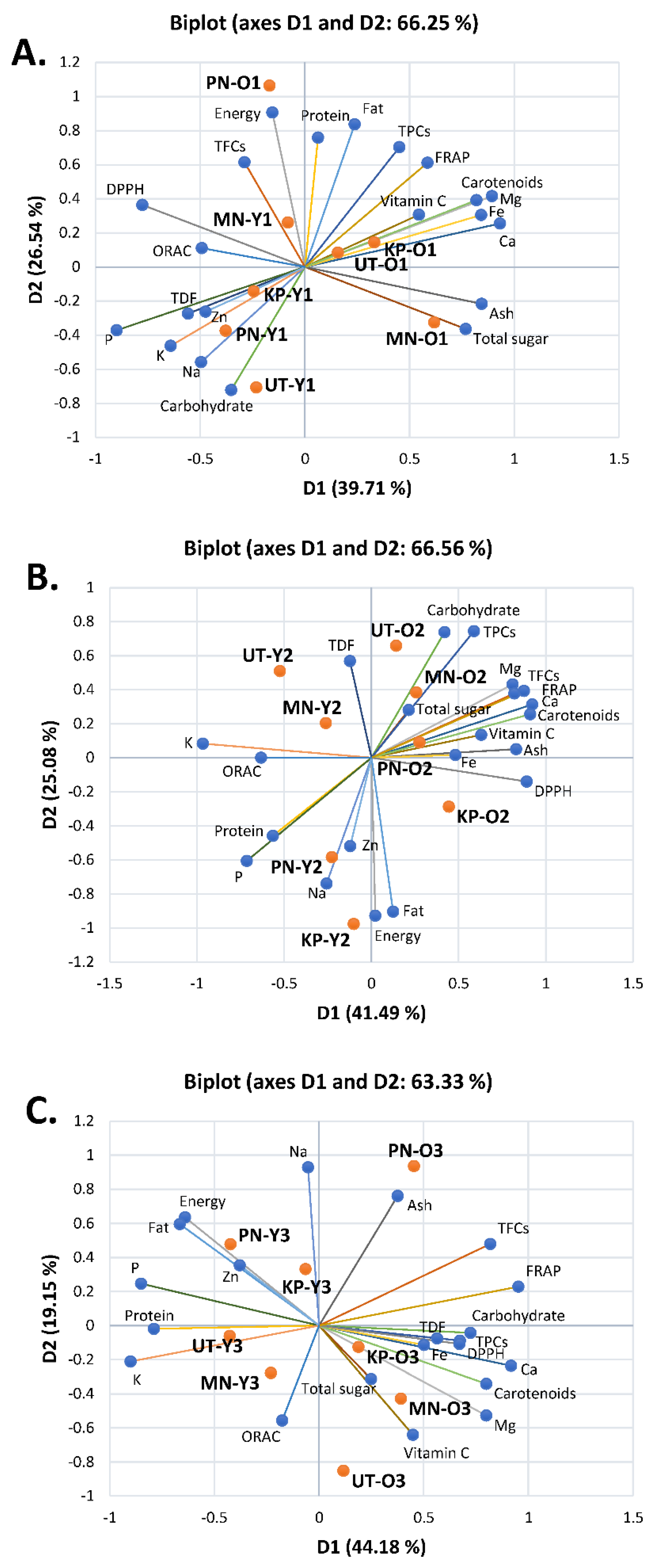
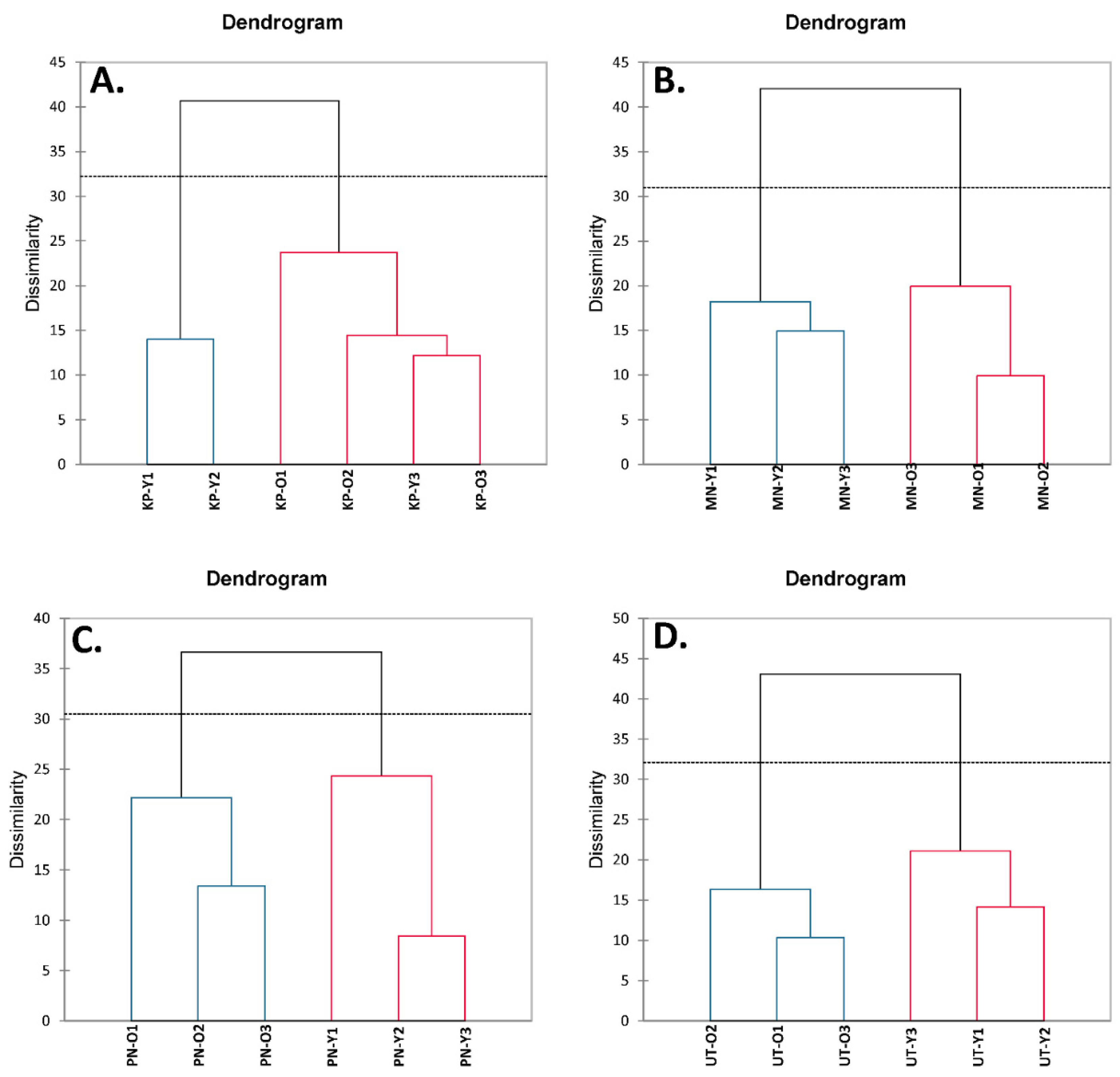
3.4. Correlation by Principal Component Analysis (PCA) and Hierarchical Cluster Analysis (HCA)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polito, L.; Bortolotti, M.; Maiello, S.; Battelli, M.G.; Bolognesi, A. Plants producing robosome-inactivateing proteins in traditional medicine. Molecules 2016, 21, 1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wannasaksri, W.; On-Nom, N.; Chupeerach, C.; Temviriyanukul, P.; Charoenkiatkul, S.; Suttisansanee, U. In Vitro Phytotherapeutic Properties of Aqueous Extracted Adenia viridiflora Craib. towards Civilization Diseases. Molecules 2021, 26, 1082. [Google Scholar] [CrossRef]
- Latimer, G.W. Official Method of Analysis of AOAC International, 21th ed.; AOAC International: Rockville, ML, USA, 2019. [Google Scholar]
- Chupeerach, C.; Aursalung, A.; Watcharachaisoponsiri, T.; Whanmek, K.; Thiyajai, P.; Yosphan, K.; Sritalahareuthai, V.; Sahasakul, Y.; Santivarangkna, C.; Suttisansanee, U. The Effect of Steaming and Fermentation on Nutritive Values, Antioxidant Activities, and Inhibitory Properties of Tea Leaves. Foods 2021, 10, 117. [Google Scholar] [CrossRef] [PubMed]
- Hinkaew, J.; Aursalung, A.; Sahasakul, Y.; Tangsuphoom, N.; Suttisansanee, U. A Comparison of Nutritional and Biochemical Quality of Date Palm Fruits Obtained from Different Planting Techniques. Molecules 2021, 26, 2245. [Google Scholar] [CrossRef]
- Sritalahareuthai, V.; Aursalung, A.; On-Nom, N.; Temviriyanukul, P.; Charoenkiatkul, S.; Suttisansanee, U. Nutritional composition of conserved Kadsura spp. plants in Northern Thailand. Heliyon 2020, 6, e04451. [Google Scholar] [CrossRef] [PubMed]
- Thuphairo, K.; Sornchan, P.; Suttisansanee, U. Bioactive Compounds, Antioxidant Activity and Inhibition of Key Enzymes Relevant to Alzheimer’s Disease from Sweet Pepper (Capsicum annuum) Extracts. Prev. Nutr. Food Sci. 2019, 24, 327–337. [Google Scholar] [CrossRef]
- Suttisansanee, U.; Thiyajai, P.; Chalermchaiwat, P.; Wongwathanarat, K.; Pruesapan, K.; Charoenkiatkul, S.; Temviriyanukul, P. Phytochemicals and In Vitro Bioactivities of Aqueous Ethanolic Extracts from Common Vegetables in Thai Food. Plants 2021, 10, 1563. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Sripum, C.; Kukreja, R.K.; Charoenkiatkul, S.; Kriengsinyos, W.; Suttisansanee, U. The effect of extraction conditions on antioxidant activities and total phenolic contents of different processed Thai Jasmine rice. Int. Food Res. J. 2017, 24, 1644–1650. [Google Scholar]
- Magdalene, O.A.; Okpashi, V.E.; Bayim, B.P. Comparative Evaluation of Proximate Composition of Selected Wild-edible Plants in Central Cross River State. J. Sci. Eng. Technol. 2019, 6, 1–8. [Google Scholar]
- Nnamani, C.V.; Oselebe, H.O.; Agbatutu, A. Assessment of nutritional values of three underutilized indigenous leafy vegetables of Ebonyi State, Nigeria. Afr. J. Biotechnol. 2009, 8, 2321–2324. [Google Scholar]
- Maroyi, A. Evaluation of Medicinal uses, Phytochemistry and Biological Activities of Adenia gummifera (Harv.) Harms. J. Pharm. Nutr. Sci. 2020, 10, 280–286. [Google Scholar] [CrossRef]
- Salim, A.; Hasyim, M.; Adam, A. Nutrient Contents of Moringa Leaves based on Leaf Age. Indian J. Public Health Res. Dev. 2018, 6, 397–401. [Google Scholar] [CrossRef]
- Maillard, A.; Diquélou, S.; Billard, V.; Laîné, P.; Garnica, M.; Prudent, M.; Garcia-Mina, J.-M.; Yvin, J.-C.; Ourry, A. Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Front. Plant Sci. 2015, 6, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flyman, M.V.; Afolayan, A.J. Effect of plant maturity on the mineral content of the leaves of Monordica balsamina L. and Vigna unguiculata subsp. sesquipedalis (L.) Verdc. J. Food Qual. 2008, 31, 661–671. [Google Scholar] [CrossRef]
- Ayua, E.; Mugalavai, V.; Simon, J.; Weller, S.; Obura, P.; Nyabinda, N. Ascorbic acid content in leaves of Nightshade (Solanum spp.) and spider plant (Cleome gynandra) varieties grown under different fertilizer regimes in Western Kenya. Afr. J. Biotechnol. 2016, 15, 199–206. [Google Scholar]
- Nobossé, P.; Fombang, E.N.; Mbofung, C.M.F. Effects of age and extraction solvent on phytochemical content and antioxidant activity of fresh Moringa oleifera L. leaves. Food Sci. Nutr. 2018, 6, 2188–2198. [Google Scholar] [CrossRef] [PubMed]
- Wingler, A.; Stangberg, E.J.; Saxena, T.; Mistry, R. Interactions Between Temperature and Sugars in the Regulation of Leaf Senescence in the Perennial Herb Arabis alpina L. J. Integr. Plant Biol. 2012, 54, 595–605. [Google Scholar] [CrossRef]
- Chang, X.; Lu, Y.; Lin, Z.; Qiu, I.; Guo, X.; Pan, J.; Abbasi, A.M. Impact of Leaf Development Stages on Polyphenolics Profile and Antioxidant Activity in Clausena lansium (Lour.) Skeels. BioMed Res. Int. 2018, 2018, 7093691. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, S.B.; Rabhi, M.; Harbaoui, F.; Zar-kalai, F.; Lachâal, M.; Karray-Bouraoui, N. Distribution of phenolic compounds and antioxidant activity between young and old leaves of Carthamus tinctorius L. and their induction by salt stress. Acta Physiol. Plant 2013, 35, 1161–1169. [Google Scholar] [CrossRef]
- Liu, Z.; Bruins, M.E.; de Bruijn, W.J.C.; Vincken, J. A comparison of the phenolic composition of old and young tea leaves reveals a decrease in flavanols and phenolic acids and an increase in flavonols upon tea leaf maturation. J. Food Compost. Anal. 2020, 86, 103385. [Google Scholar] [CrossRef]
- Zhang, T.; Zheng, J.; Yu, Z.; Huang, X.; Zhang, Q.; Tian, X.; Peng, C. Functional characteristics of phenolic compounds accumulated in young leaves of two subtropical forest tree species of different successional stages. Tree Physiol. 2018, 38, 1486–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum-Silva, C.H.; Chaves, V.C.; Schenkel, E.P.; Coelho, G.C.; Reginatto, F.H. The influence of leaf age on methylxanthines, total phenolic content, and free radical scavenging capacity of Ilex paraguariensis aqueous extracts. Rev. Bras Farmacogn. 2015, 25, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Soufan, W.; Okla, M.K.; Salamatullah, A.; Hayat, K.; Abdel-Maksoud, M.A.; Al-Amri, S.S. Seasonal variation in yield, nutritive value, and antioxidant capacity of leaves of alfalfa plants grown in arid climate of Saudi Arabia. Chil. J. Agric. Res. 2021, 81, 182–190. [Google Scholar] [CrossRef]
- Abebe, A.; Tolera, A.; Holand, Ø.; Ådnøy, T.; Eik, L.O. Seasonal variation in nutritive value of some browse and grass species in Borane rangeland, southern Ethiopia. Trop. Subtrop. Agroecosyst. 2012, 15, 261–271. [Google Scholar]
- Oliveira, M.; João Rodrigues, M.; Neng, N.R.; Nogueira, J.M.F.; Bessa, R.J.B.; Custódio, L. Seasonal Variations of the Nutritive Value and Phytotherapeutic Potential of Cladium mariscus L. (Pohl.) Targeting Ruminant’s Production. Plants 2021, 10, 556. [Google Scholar] [CrossRef]
- Ribeiro, D.A.; Camilo, C.J.; de Fátima Alves Nonato, C.; Rodrigues, F.F.G.; Menezes, I.R.A.; Ribeiro-Filho, J.; Xiao, J.; de Almeida Souza, M.M.; da Costa, J.G.M. Influence of seasonal variation on phenolic content and in vitro antioxidant activity of Secondatia floribunda A. DC. (Apocynaceae). Food Chem. 2020, 315, 126277. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Plant Secondary Metabolism: Diversity, Function and its Evolution. Nat. Prod. Commun. 2008, 3, 1205–1216. [Google Scholar] [CrossRef] [Green Version]
- Demain, A.L.; Fang, A. The natural functions of secondary metabolites. Adv. Biochem. Eng. Biotechnol. 2000, 69, 1–39. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Harvesting Times (Month/2018) | Rainfall (mm) |
|---|---|
| March | 7.8 |
| April | 289.0 |
| May | 214.8 |
| June | 41.3 |
| July | 67.7 |
| August | 94.9 |
| Nutrients | Young Shoots | Old Leaves | ||||
|---|---|---|---|---|---|---|
| March–April | May–June | July–August | March–April | May–June | July–August | |
| Energy (kcal) | ||||||
| KP | 381.52 ± 4.36 aB | 387.69 ± 0.08 aA * | 381.06 ± 0.28 cB * | 380.80 ± 2.27 bA | 379.72 ± 0.40 aA | 380.17 ± 4.06 abA |
| MN | 382.50 ± 1.47 aA * | 376.99 ± 0.93 cB | 381.92 ± 1.41 bcA | 373.22 ± 0.68 cB | 376.08 ± 1.67 aAB | 376.79 ± 1.90 bA |
| PN | 376.42 ± 0.24 bC * | 381.52 ± 0.79 bB * | 386.55 ± 0.51 aA | 386.97 ± 0.73 aA | 376.31 ± 1.16 aC | 382.95 ± 2.80 aB |
| UT | 371.45 ± 0.11 cC * | 373.95 ± 0.97 dB | 382.76 ± 0.56 bA * | 382.44 ± 1.24 bA | 376.66 ± 3.32 aB | 379.72 ± 0.04 abAB |
| Protein (g) | ||||||
| KP | 20.82 ± 0.36 aA | 19.33 ± 0.06 bB * | 18.74 ± 0.48 bB | 21.22 ± 0.13 bA | 17.19 ± 0.06 cC | 19.48 ± 0.29 aB |
| MN | 20.73 ± 0.72 aA | 20.04 ± 0.29 bA * | 18.43 ± 0.21 bB | 19.85 ± 0.28 cA | 18.19 ± 0.19 bB | 18.02 ± 0.63 bB |
| PN | 18.15 ± 0.18 cC * | 21.94 ± 0.22 aA * | 20.87 ± 0.24 aB * | 22.68 ± 0.29 aA | 19.67 ± 0.26 aB | 17.80 ± 0.25 bC |
| UT | 19.66 ± 0.00 bAB * | 19.15 ± 1.20 bC * | 20.76 ± 0.48 aA * | 18.98 ± 0.28 dA | 16.53 ± 0.45 dB | 19.21 ± 0.40 aA |
| Fat (g) | ||||||
| KP | 2.43 ± 0.71 aB | 4.10 ± 0.14 aA * | 3.14 ± 0.14 aB | 2.29 ± 0.48 bA | 3.28 ± 0.19 aA | 2.91 ± 0.73 aA |
| MN | 2.35 ± 0.15 aA * | 2.01 ± 0.08 cB | 2.61 ± 0.24 bA | 1.66 ± 0.19 cA | 2.31 ± 0.29 bA | 2.24 ± 0.64 aA |
| PN | 0.62 ± 0.07 bC * | 2.77 ± 0.05 bB * | 3.27 ± 0.00 aA | 3.09 ± 0.11 aA | 2.13 ± 0.20 bB | 2.66 ± 0.43 aAB |
| UT | 0.64 ± 0.04 bC * | 2.08 ± 0.28 cB | 3.14 ± 0.23 aA * | 2.69 ± 0.18 abA | 1.82 ± 0.72 bA | 2.23 ± 0.06 aA |
| Carbohydrate (g) | ||||||
| KP | 69.09 ± 0.13 cA | 68.36 ± 0.36 abA * | 69.47 ± 0.85 bA | 68.82 ± 0.64 bB | 70.35 ± 0.28 bA | 69.02 ± 0.35 bB |
| MN | 69.60 ± 0.68 cB | 69.68 ± 0.34 aB * | 71.18 ± 0.02 aA | 69.72 ± 0.54 aA | 70.62 ± 0.43 bA | 71.13 ± 1.60 aA |
| PN | 74.56 ± 0.28 aA * | 67.21 ± 0.54 cC * | 68.42 ± 0.12 bcB * | 67.12 ± 0.35 cC | 69.63 ± 0.10 bB | 71.94 ± 0.03 aA |
| UT | 71.76 ± 0.06 bA * | 69.65 ± 1.59 aB * | 67.87 ± 0.85 cB * | 70.57 ± 0.18 aB | 73.53 ± 1.24 aA | 70.69 ± 0.55 aB |
| Ash (g) | ||||||
| KP | 7.66 ± 0.21 bC | 8.21 ± 0.16 bB * | 8.66 ± 0.24 aA | 7.67 ± 0.03 bC | 9.17 ± 0.14 aA | 8.59 ± 0.10 aB |
| MN | 7.31 ± 0.18 cC * | 8.27 ± 0.13 bA * | 7.79 ± 0.05 cB * | 8.77 ± 0.06 aA | 8.87 ± 0.05 bA | 8.61 ± 0.33 aA |
| PN | 6.67 ± 0.03 dC * | 8.08 ± 0.27 bA * | 7.44 ± 0.12 dB | 7.11 ± 0.05 cC | 8.58 ± 0.04 cA | 7.59 ± 0.16 bB |
| UT | 7.94 ± 0.02 aC * | 9.12 ± 0.11 aA * | 8.23 ± 0.15 bB * | 7.75 ± 0.08 bB | 8.12 ± 0.07 dA | 7.86 ± 0.09 bB |
| Nutrients | Young Shoots | Old Leaves | ||||
|---|---|---|---|---|---|---|
| March–April | May–June | July–August | March–April | May–June | July–August | |
| Total dietary fiber (g) | ||||||
| KP | 36.16 ± 0.90 cB * | 36.54 ± 0.97 dB * | 58.16 ± 2.06 aA * | 29.15 ± 0.38 cC | 42.89 ± 0.40 bB | 51.46 ± 0.99 aA |
| MN | 32.89 ± 0.41 dB * | 49.97 ± 0.59 bA * | 48.57 ± 2.50 bA | 34.39 ± 0.09 bB | 44.60 ± 0.47 abA | 48.10 ± 3.85 abA |
| PN | 43.98 ± 1.39 bB * | 40.98 ± 0.68 cB * | 50.75 ± 2.63 bA | 39.32 ± 1.29 aC | 42.93 ± 0.48 bB | 46.42 ± 1.19 bA |
| UT | 60.46 ± 1.28 aA * | 65.26 ± 4.06 aA * | 52.11 ± 0.55 bB * | 28.37 ± 0.12 cB | 46.43 ± 1.80 aA | 46.49 ± 0.98 bA |
| Soluble dietary fiber (g) | ||||||
| KP | 5.91 ± 1.22 cB | 11.84 ± 0.16 bA * | 10.39 ± 0.86 abA | 4.32 ± 0.13 dC | 10.64 ± 0.66 cB | 12.86 ± 1.45 aA |
| MN | 3.43 ± 0.01 dB * | 9.52 ± 0.12 cA * | 8.85 ± 1.53 bA * | 6.25 ± 0.09 bC | 12.50 ± 0.52 bB | 14.58 ± 1.70 aA |
| PN | 12.44 ± 0.32 bA * | 10.83 ± 1.42 bcA * | 12.10 ± 1.19 aA | 11.20 ± 0.16 aB | 18.96 ± 0.67 aA | 12.58 ± 1.38 aB |
| UT | 15.19 ± 1.45 aB * | 20.47 ± 1.69 aA | 11.88 ± 1.74 aC | 4.91 ± 0.09 cC | 17.93 ± 1.26 aA | 12.27 ± 0.31 aB |
| Insoluble dietary fiber (g) | ||||||
| KP | 30.25 ± 0.32 cB * | 24.70 ± 1.13 dC * | 47.77 ± 1.20 aA * | 24.82 ± 0.25 bC | 32.25 ± 0.26 aB | 38.60 ± 0.46 aA |
| MN | 29.46 ± 0.42 cB * | 40.45 ± 0.47 bA * | 39.72 ± 0.97 bA * | 28.14 ± 0.00 aB | 32.11 ± 0.06 aA | 33.52 ± 2.15 bA |
| PN | 31.53 ± 1.07 bB * | 30.14 ± 0.74 cB * | 38.65 ± 1.43 bA * | 28.12 ± 1.13 aB | 23.97 ± 0.19 cC | 33.84 ± 0.18 bA |
| UT | 45.27 ± 0.17 aA * | 44.79 ± 2.37 aA * | 40.23 ± 1.19 bB * | 23.46 ± 0.21 cC | 28.50 ± 0.53 bB | 34.22 ± 1.29 bA |
| Total sugar (g) | ||||||
| KP | 11.49 ± 0.25 aB * | 12.11 ± 0.35 bB * | 15.00 ± 0.45 bA * | 15.15 ± 0.07 aB | 13.44 ± 0.70 bC | 18.04 ± 0.06 aA |
| MN | 8.98 ± 0.23 cB * | 8.45 ± 0.05 dC * | 16.02 ± 0.31 aA * | 13.96 ± 0.20 bB | 14.66 ± 0.27 aB | 18.19 ± 0.69 aA |
| PN | 8.51 ± 0.19 dC * | 10.04 ± 0.49 cB * | 14.89 ± 0.03 bA * | 11.83 ± 0.11 dC | 13.64 ± 0.05 abB | 18.22 ± 0.57 aA |
| UT | 10.81 ± 0.14 bC * | 14.60 ± 0.16 aB | 16.16 ± 0.08 aA | 13.57 ± 0.16 cB | 13.53 ± 0.85 bB | 16.60 ± 0.54 bA |
| Fructose (g) | ||||||
| KP | 2.02 ± 0.12 aC * | 4.04 ± 0.04 bA * | 3.37 ± 0.11 bB * | 5.16 ± 0.28 aB | 4.89 ± 0.09 cB | 6.21 ± 0.08 aA |
| MN | 2.09 ± 0.17 aC * | 4.03 ± 0.14 bB * | 4.48 ± 0.13 aA * | 4.32 ± 0.01 bC | 5.49 ± 0.12 aB | 6.44 ± 0.23 aA |
| PN | 1.50 ± 0.00 bC * | 3.83 ± 0.19 bA * | 3.06 ± 0.00 cB * | 4.58 ± 0.18 bB | 4.81 ± 0.01 cAB | 5.03 ± 0.12 bA |
| UT | 0.88 ± 0.07 cC * | 4.62 ± 0.11 aA * | 3.24 ± 0.05 bB * | 4.50 ± 0.10 bB | 5.15 ± 0.00 bA | 5.34 ± 0.20 bA |
| Glucose (g) | ||||||
| KP | 9.47 ± 0.12 bA | 8.06 ± 0.31 bB | 9.71 ± 0.25 bA | 9.44 ± 0.18 aA | 7.48 ± 0.49 aB | 9.89 ± 0.13 aA |
| MN | 6.88 ± 0.06 cB * | 4.42 ± 0.19 dC * | 9.92 ± 0.16 bA | 7.66 ± 0.20 cC | 8.30 ± 0.12 aB | 10.15 ± 0.42 aA |
| PN | 7.01 ± 0.19 cB | 6.21 ± 0.68 cB * | 9.98 ± 0.12 bA | 7.25 ± 0.08 dC | 8.05 ± 0.09 aB | 10.11 ± 0.45 aA |
| UT | 9.93 ± 0.07 aB * | 9.98 ± 0.28 aB * | 10.88 ± 0.11 aA * | 9.07 ± 0.26 bAB | 8.38 ± 0.86 aB | 9.52 ± 0.26 aA |
| Sucrose (g) | ||||||
| KP | ND | ND | 1.91 ± 0.08 b | 0.55 ± 0.02 bC | 1.08 ± 0.13 aB | 1.94 ± 0.01 bA |
| MN | <LOD | ND | 1.62 ± 0.02 c | 1.98 ± 0.01 aA | 0.87 ± 0.03 bC | 1.60 ± 0.04 dB |
| PN | <LOD | ND | 1.86 ± 0.08 b * | ND | 0.78 ± 0.03 bB | 3.09 ± 0.01 aA |
| UT | <LOD | ND | 2.04 ± 0.02 a * | <LOD | ND | 1.74 ± 0.08 cA |
| Nutrients | Young Shoots | Old Leaves | ||||
|---|---|---|---|---|---|---|
| March–April | May–June | July–August | March–April | May–June | July–August | |
| Vitamin C (mg) | ||||||
| KP | 754.27 ± 28.67 bC * | 1034.83 ± 26.26 bB * | 1292.44 ± 99.87 bA * | 1058.37 ± 37.71 bC | 1349.45 ± 0.35 bB | 1841.90 ± 28.65 bA |
| MN | 776.90 ± 51.55 bC * | 1019.73 ± 41.34 bB * | 1311.54 ± 2.98 bA * | 862.64 ± 1.27 dC | 1105.31 ± 12.07 dB | 1598.29 ± 53.21 cA |
| PN | 573.93 ± 23.69 cC * | 1231.19 ± 79.40 aA | 800.81 ± 72.52 cB * | 928.17 ± 8.76 cC | 1200.02 ± 32.83 cB | 1501.99 ± 20.89 dA |
| UT | 879.40 ± 10.81 aC * | 1007.39 ± 56.25 bB * | 1696.46 ± 18.22 aA * | 1181.70 ± 6.03 aC | 1439.03 ± 42.55 aB | 2240.65 ± 39.79 aA |
| Lutein (mg) | ||||||
| KP | 65.20 ± 0.21 aA * | 55.29 ± 0.39 aB * | 48.74 ± 3.24 aC * | 93.49 ± 3.22 aA | 80.00 ± 3.22 aB | 79.74 ± 3.89 aB |
| MN | 49.75 ± 0.07 bA * | 47.94 ± 3.57 bA * | 31.00 ± 3.12 bB * | 84.61 ± 2.94 bA | 82.82 ± 3.13 aA | 64.10 ± 4.42 bB |
| PN | 33.25 ± 1.55 dA * | 26.47 ± 1.67 dC * | 30.44 ± 0.73 bB * | 62.43 ± 4.25 dB | 70.12 ± 0.17 bA | 50.26 ± 2.62 cC |
| UT | 38.10 ± 0.47 cB * | 41.32 ± 1.00 cA * | 33.92 ± 2.50 bC * | 69.30 ± 3.83 cAB | 66.04 ± 1.28 bB | 75.09 ± 3.91 aA |
| β-carotene(mg) | ||||||
| KP | 27.24 ± 0.91 aA * | 20.14 ± 1.32 aB * | 18.65 ± 1.19 aB * | 47.04 ± 2.67 aA | 41.71 ± 1.57 bAB | 37.19 ± 3.47 aB |
| MN | 20.78 ± 0.09 bA * | 18.50 ± 1.22 aA * | 12.89 ± 0.84 bC * | 45.77 ± 1.28 aA | 44.46 ± 0.91 bA | 33.70 ± 2.52 aB |
| PN | 19.36 ± 1.58 bA * | 13.94 ± 0.90 bB * | 18.08 ± 0.86 aA * | 44.74 ± 3.07 aB | 49.84 ± 0.53 aA | 35.28 ± 1.55 aC |
| UT | 14.51 ± 0.01 cA * | 14.49 ± 0.25 bA * | 12.08 ± 1.00 bB * | 34.39 ± 2.18 bA | 31.52 ± 2.59 cA | 34.99 ± 0.91 aA |
| Nutrients | Young Shoots | Old Leaves | ||||
|---|---|---|---|---|---|---|
| March–April | May–June | July–August | March–April | May–June | July–August | |
| Calcium (mg) | ||||||
| KP | 653.93 ± 29.03 aC * | 920.61 ± 4.37 aA * | 854.27 ± 29.36 aB * | 1137.28 ± 4.22 bC | 1645.10 ± 63.78 aA | 1277.89 ± 17.38 bB |
| MN | 669.84 ± 1.44 aB * | 895.96 ± 40.42 aA * | 907.11 ± 19.55 aA * | 1610.80 ± 23.08 aA | 1642.38 ± 105.65 aA | 1438.75 ± 83.01 aB |
| PN | 397.97 ± 0.22 bC * | 631.55 ± 37.25 cA * | 529.35 ± 25.74 cB * | 968.40 ± 11.87 cC | 1554.59 ± 14.57 aA | 1165.90 ± 19.29 cB |
| UT | 418.83 ± 7.81 bC * | 786.85 ± 70.01 bA * | 621.94 ± 36.62 bB * | 977.39 ± 15.34 cB | 1371.91 ± 26.54 bA | 1000.70 ± 33.63 dB |
| Phosphorus (mg) | ||||||
| KP | 534.08 ± 71.72 aA | 601.59 ± 82.19 aA | 561.18 ± 36.40 aA * | 450.88 ± 10.80 bB | 477.53 ± 6.97 aA | 433.28 ± 1.29 bC |
| MN | 477.13 ± 19.34 aA * | 512.81 ± 6.65 aA * | 467.56 ± 34.41 bA | 418.16 ± 11.73 cA | 401.93 ± 11.60 bA | 428.15 ± 19.11 bA |
| PN | 531.45 ± 4.85 aA * | 535.66 ± 59.77 aA * | 579.87 ± 0.55 aA * | 472.77 ± 3.33 aA | 418.90 ± 18.29 bB | 421.54 ± 17.10 bB |
| UT | 537.39 ± 57.99 aA | 555.34 ± 48.21 aA * | 557.51 ± 9.87 aA * | 477.52 ± 11.11 aA | 432.89 ± 28.95 bB | 459.07 ± 3.83 aAB |
| Sodium (mg) | ||||||
| KP | 135.42 ± 50.46 aA | 132.56 ± 26.68 aA * | 102.97 ± 55.61 aA | 62.39 ± 12.25 abB | 86.01 ± 1.16 bA | 59.71 ± 0.20 bB |
| MN | 55.11 ± 3.16 bA | 86.23 ± 45.18 aA | 51.74 ± 7.64 aA | 74.86 ± 37.24 aA | 61.59 ± 16.87 bA | 48.68 ± 5.15 cA |
| PN | 120.22 ± 9.21 aAB * | 152.02 ± 43.54 aA | 87.49 ± 11.24 aA | 52.32 ± 12.15 abC | 131.97 ± 2.97 aA | 103.75 ± 0.60 aB |
| UT | 95.99 ± 38.10 abA * | 93.04 ± 33.50 aA | 69.43 ± 1.85 aA * | 30.65 ± 4.48 bA | 66.94 ± 32.50 bA | 44.30 ± 7.59 cA |
| Potassium (mg) | ||||||
| KP | 2077.47 ± 93.61 aB * | 1949.93 ± 26.25 bB * | 2356.72 ± 79.16 bA * | 1543.54 ± 7.56 bB | 1543.54 ± 8.89 cB | 2001.22 ± 71.09 bA |
| MN | 1810.88 ± 73.40 bB * | 2186.31 ± 71.34 bA * | 2177.13 ± 39.18 cA * | 1357.16 ± 81.06 cB | 1877.61 ± 162.93 aA | 1983.55 ± 51.67 bA |
| PN | 1740.09 ± 66.25 bC * | 2197.43 ± 8.66 bB * | 2443.19 ± 28.34 bA * | 1497.68 ± 33.61 bcB | 1709.12 ± 34.03 bA | 1587.42 ± 107.86 cAB |
| UT | 2177.00 ± 79.72 aB * | 2475.16 ± 267.19 aAB * | 2549.33 ± 50.89 aA * | 1852.70 ± 133.06 aB | 1741.04 ± 23.68 abB | 2144.11 ± 8.72 aA |
| Magnesium (mg) | ||||||
| KP | 307.49 ± 21.15 bB * | 217.54 ± 1.97 bC * | 473.54 ± 31.55 aA * | 463.15 ± 6.33 bB | 341.69 ± 9.35 aC | 762.89 ± 15.67 bA |
| MN | 357.48 ± 16.83 aB * | 298.00 ± 22.65 aC * | 450.54 ± 10.53 abA * | 539.60 ± 6.68 aB | 344.00 ± 17.27 aC | 951.84 ± 31.55 aA |
| PN | 242.92 ± 4.00 cB * | 247.40 ± 2.70 bB * | 378.99 ± 17.32 cA * | 427.70 ± 3.43 cB | 355.33 ± 1.72 aC | 601.00 ± 7.97 cA |
| UT | 245.26 ± 11.72 cB * | 236.51 ± 21.54 bB * | 428.42 ± 7.40 bA * | 394.97 ± 4.37 dB | 305.46 ± 2.69 bC | 798.77 ± 20.61 bA |
| Iron (mg) | ||||||
| KP | 4.84 ± 0.27 aB * | 4.28 ± 0.32 cC * | 6.06 ± 0.06 aA * | 6.24 ± 0.01 bB | 6.49 ± 0.12 bAB | 7.51 ± 0.89 aA |
| MN | 4.66 ± 0.03 abB * | 5.19 ± 0.04 bA * | 4.93 ± 0.29 cAB * | 8.57 ± 1.24 aA | 7.08 ± 0.12 aAB | 6.43 ± 0.63 abB |
| PN | 4.27 ± 0.11 bC * | 7.08 ± 0.39 aA | 5.63 ± 0.14 bB | 6.83 ± 0.08 bA | 7.29 ± 0.44 aA | 5.71 ± 0.18 bB |
| UT | 4.39 ± 0.31 bB * | 5.08 ± 0.36 bA | 5.49 ± 0.25 bA | 6.35 ± 0.26 bA | 4.90 ± 0.20 cB | 5.84 ± 0.46 bA |
| Zinc (mg) | ||||||
| KP | 3.80 ± 0.27 aA * | 3.76 ± 0.06 aA | 3.89 ± 0.17 aA * | 2.99 ± 0.12 aC | 3.90 ± 0.15 aA | 3.46 ± 0.01 aB |
| MN | 2.25 ± 0.01 cC | 2.63 ± 0.17 bB | 3.19 ± 0.03 cA * | 2.60 ± 0.27 bB | 2.59 ± 0.17 dB | 3.54 ± 0.07 aA |
| PN | 3.13 ± 0.12 bB | 3.68 ± 0.10 aA | 3.79 ± 0.04 aA * | 2.93 ± 0.11 aB | 3.44 ± 0.25 bA | 3.14 ± 0.20 bAB |
| UT | 3.17 ± 0.45 bB | 3.85 ± 0.17 aA * | 3.50 ± 0.09 bAB * | 2.59 ± 0.07 bB | 3.07 ± 0.02 cA | 3.00 ± 0.11 bA |
| Activities | Young Shoots | Old Leaves | ||||
|---|---|---|---|---|---|---|
| March–April # | May–June | July–August | March–April # | May–June | July–August | |
| Total phenolic contents (mg GAE/g DW) | ||||||
| KP | 20.57 ± 0.72 cB * | 18.94 ± 1.20 cC * | 26.95 ± 1.81 aA | 28.70 ± 1.79 aA | 23.71 ± 1.05 cB | 23.84 ± 1.99 bB |
| MN | 26.35 ± 1.84 aA | 22.47 ± 1.25 aB * | 20.79 ± 1.41 bcC * | 25.37 ± 1.16 bA | 24.86 ± 0.93 bA | 24.38 ± 1.43 bA |
| PN | 23.57 ± 0.92 bA * | 20.59 ± 1.92 bB * | 21.39 ± 0.97 bB * | 27.89 ± 1.61 aA | 24.80 ± 0.64 bB | 25.70 ± 2.02 bB |
| UT | 20.38 ± 0.79 cB * | 21.50 ± 0.48 abA * | 19.69 ± 1.21 cB * | 23.56 ± 1.34 cB | 28.05 ± 1.44 aA | 27.97 ± 2.44 aA |
| Total flavonoid contents (mg QE/g DW) | ||||||
| KP | 7.35 ± 0.42 bB * | 5.42 ± 0.54 bC * | 10.91 ± 0.92 aA | 5.96 ± 0.35 cC | 8.42 ± 0.72 bB | 10.84 ± 0.88 bA |
| MN | 7.15 ± 0.50 bB * | 5.37 ± 0.52 bC * | 8.78 ± 0.83 cA * | 8.35 ± 0.61 bC | 9.32 ± 0.46 aB | 11.92 ± 0.64 aA |
| PN | 9.65 ± 0.72 aA * | 6.24 ± 0.37 aB * | 9.98 ± 0.87 bA * | 14.08 ± 1.41 aA | 8.44 ± 0.31 bC | 12.61 ± 0.52 aB |
| UT | 7.50 ± 0.69 bB * | 6.24 ± 0.59 aC * | 8.53 ± 0.64 cA * | 8.38 ± 0.33 bB | 7.60 ± 0.63 cC | 9.38 ± 0.77 cA |
| 2,2-Diphenyl-1-picrylhydrazyl (DPPH)radical scavenging activities (µmol TE/100 g DW) | ||||||
| KP | 1.25 ± 0.06 bA * | 0.98 ± 0.08 aC | 1.15 ± 0.02 aB * | 0.96 ± 0.07 bA | 1.02 ± 0.09 aA | 1.03 ± 0.05 bA |
| MN | 1.37 ± 0.04 aA * | 0.86 ± 0.07 bC * | 1.02 ± 0.07 bB * | 0.99 ± 0.06 bB | 1.02 ± 0.10 aB | 1.16 ± 0.02 aA |
| PN | 1.28 ± 0.02 bA * | 0.95 ± 0.03 aC * | 1.02 ± 0.04 bB * | 1.44 ± 0.10 aA | 1.04 ± 0.08 aC | 1.12 ± 0.06 aB |
| UT | 1.23 ± 0.10 bA * | 0.78 ± 0.06 cC * | 1.03 ± 0.08 bB * | 0.97 ± 0.05 bC | 1.05 ± 0.09 aB | 1.14 ± 0.09 aA |
| Ferric reducing antioxidant power (FRAP)activities (µmol TE/g DW) | ||||||
| KP | 16.52 ± 0.97 bC * | 20.20 ± 1.73 bB * | 25.63 ± 1.64 aA * | 34.08 ± 3.12 bB | 39.83 ± 3.99 aA | 30.72 ± 0.88 cC |
| MN | 29.24 ± 2.88 aA * | 22.76 ± 2.22 aB * | 19.47 ± 1.68 cC * | 35.74 ± 3.27 abA | 34.91 ± 3.19 bA | 34.20 ± 2.10 bA |
| PN | 28.03 ± 0.98 aA * | 22.73 ± 1.43 aB * | 22.42 ± 1.59 bB * | 37.00 ± 1.60 aB | 40.31 ± 3.41 aA | 37.71 ± 3.33 aAB |
| UT | 14.36 ± 0.85 cC * | 19.82 ± 1.87 bA * | 17.06 ± 1.50 dB * | 35.17 ± 2.36 abB | 42.90 ± 1.68 aA | 27.08 ± 2.18 dC |
| Oxygen radical absorbance capacity (ORAC)activities (µmol TE/g DW) | ||||||
| KP | 1148.47 ± 74.70 bA * | 945.45 ± 80.39 bB | 843.16 ± 26.88 bC * | 753.77 ± 74.81 cB | 878.30 ± 71.10 bA | 557.60 ± 12.22 bC |
| MN | 842.36 ± 69.59 cA * | 819.31 ± 74.57 cA * | 879.40 ± 73.51 abA | 1094.66 ± 119.33 bA | 979.12 ± 88.65 aB | 817.34 ± 70.76 bC |
| PN | 1438.85 ± 145.27 aA | 1264.75 ± 83.41 aB * | 913.14 ± 58.62 aC * | 1403.53 ± 122.44 aA | 933.96 ± 66.54 abB | 592.08 ± 20.22 cC |
| UT | 1066.29 ± 100.98 bB | 1338.09 ± 81.11 aA * | 649.06 ± 58.15 cC * | 1106.77 ± 102.36 bB | 916.29 ± 73.94 abC | 1209.10 ± 75.07 aA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wannasaksri, W.; Temviriyanukul, P.; Aursalung, A.; Sahasakul, Y.; Thangsiri, S.; Inthachat, W.; On-Nom, N.; Chupeerach, C.; Pruesapan, K.; Charoenkiatkul, S.; et al. Influence of Plant Origins and Seasonal Variations on Nutritive Values, Phenolics and Antioxidant Activities of Adenia viridiflora Craib., an Endangered Species from Thailand. Foods 2021, 10, 2799. https://doi.org/10.3390/foods10112799
Wannasaksri W, Temviriyanukul P, Aursalung A, Sahasakul Y, Thangsiri S, Inthachat W, On-Nom N, Chupeerach C, Pruesapan K, Charoenkiatkul S, et al. Influence of Plant Origins and Seasonal Variations on Nutritive Values, Phenolics and Antioxidant Activities of Adenia viridiflora Craib., an Endangered Species from Thailand. Foods. 2021; 10(11):2799. https://doi.org/10.3390/foods10112799
Chicago/Turabian StyleWannasaksri, Werawat, Piya Temviriyanukul, Amornrat Aursalung, Yuraporn Sahasakul, Sirinapa Thangsiri, Woorawee Inthachat, Nattira On-Nom, Chaowanee Chupeerach, Kanchana Pruesapan, Somsri Charoenkiatkul, and et al. 2021. "Influence of Plant Origins and Seasonal Variations on Nutritive Values, Phenolics and Antioxidant Activities of Adenia viridiflora Craib., an Endangered Species from Thailand" Foods 10, no. 11: 2799. https://doi.org/10.3390/foods10112799