
Chemical Characterization, Nutritional and Bioactive Properties of Physalis peruviana Fruit from High Areas of the Atacama Desert

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit and Sample Preparation
2.2. Chemicals
2.3. Determination of Proximate Composition
2.4. Determination of Mineral Content
2.5. Instrument Conditions for Phenolic Compounds
2.6. Phytochemical Analysis
2.6.1. Phenolic and Flavonoid Contents
2.6.2. Total Tannin Content
2.6.3. Total Anthocyanin Content
2.7. Antioxidant Activity Assays
2.7.1. Ferric Reducing Antioxidant Power (FRAP)
2.7.2. ABTS Method
2.7.3. Oxygen Radical Absorbance Capacity
2.8. Antibacterial Activity
2.8.1. Strain and Growth Conditions
2.8.2. Minimum Inhibitory Concentration (MIC)
2.8.3. Minimum Bactericidal Concentration (MBC)
2.9. Inhibitory Activity Assays
2.9.1. α-Amylase Inhibition Method
2.9.2. α-Glucosidase Inhibition Method
2.10. Statistical Analysis
3. Results
3.1. Proximate Composition and Mineral Composition
3.2. Chemical Composition
3.3. Bioactive Compounds and Antioxidant Capacity
3.4. Antibacterial Activity
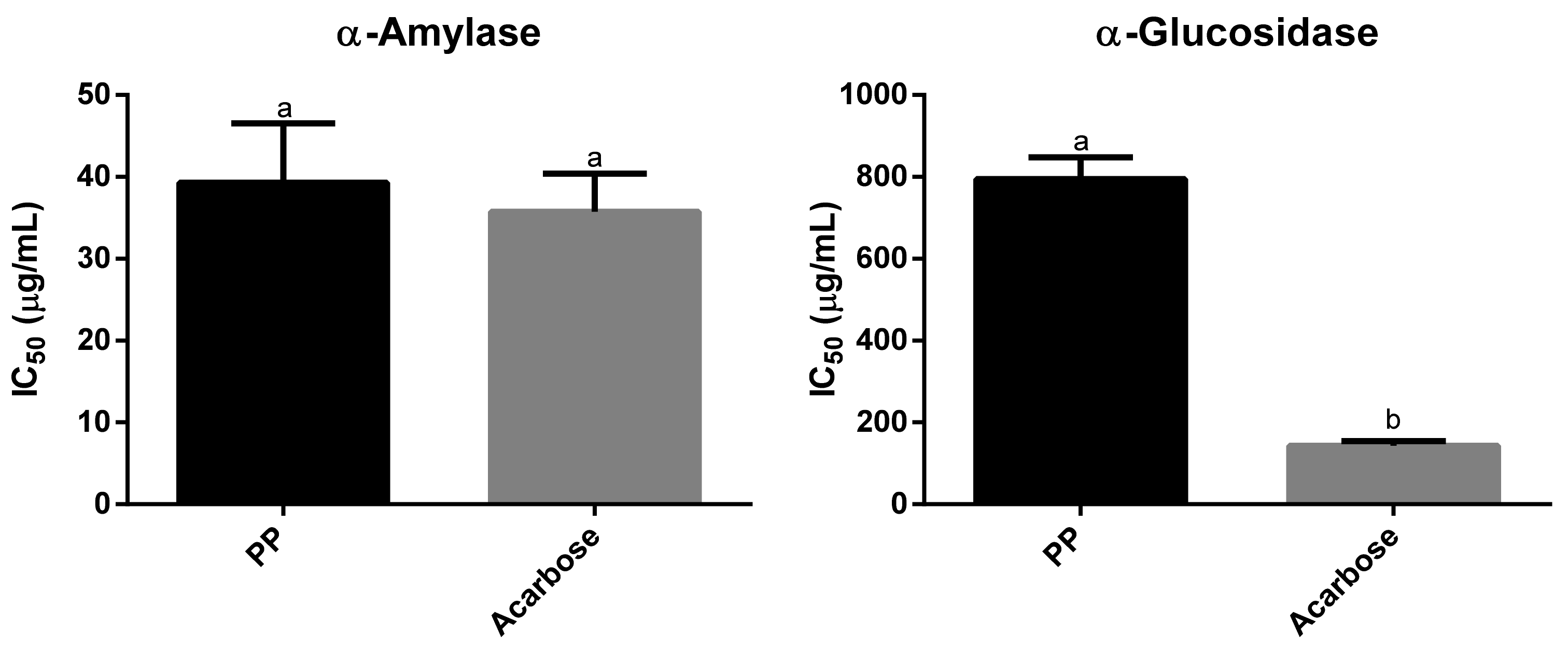
3.5. Enzyme Inhibitory Activity of Physalis peruviana
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, D.; Zhang, R.; Hou, F.; Zhang, M.; Guo, J.; Huang, F.; Deng, Y.; Wei, Z. Comparison of the free and bound phenolic profiles and cellular antioxidant activities of litchi pulp extracts from different solvents. BMC Complement. Altern. Med. 2014, 14, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega-Gálvez, A.; López, J.; Torres-Ossandón, M.J.; Galotto, M.J.; Puente-Díaz, L.; Quispe-Fuentes, I.; Di Scala, K. High hydrostatic pressure effect on chemical composition, color, phenolic acids and antioxidant capacity of Cape gooseberry pulp (Physalis peruviana L.). LWT-Food Sci. Technol. 2014, 58, 519–526. [Google Scholar] [CrossRef]
- Ramadan, M.F. Bioactive phytochemicals, nutritional value, and functional properties of cape gooseberry (Physalis peruviana): An overview. Food Res. Int. 2011, 44, 1830–1836. [Google Scholar] [CrossRef]
- Pereda, M.S.B.; Nazareno, M.A.; Viturro, C.I. Nutritional and antioxidant properties of Physalis peruviana L. fruits from the Argentinean northern Andean region. Plant Foods Hum. Nutr. 2019, 74, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Pereda, M.S.B.; Nazareno, M.A.; Viturro, C.I. Optimized formulation of a Physalis peruviana L. fruit nectar: Physicochemical characterization, sensorial traits and antioxidant properties. J. Food Sci. Technol. 2020, 57, 3267–3277. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Tenorio, M.-L.; Dekker, M.; Verkerk, R.; van Boekel, M.A. Health-promoting compounds in cape gooseberry (Physalis peruviana L.): Review from a supply chain perspective. Trends Food Sci. Technol. 2016, 57, 83–92. [Google Scholar] [CrossRef]
- Bravo, K.; Sepulveda-Ortega, S.; Lara-Guzman, O.; Navas-Arboleda, A.A.; Osorio, E. Influence of cultivar and ripening time on bioactive compounds and antioxidant properties in Cape gooseberry (Physalis peruviana L.). J. Sci. Food Agric. 2015, 95, 1562–1569. [Google Scholar] [CrossRef]
- Stein, A.; Compera, D.; Karge, B.; Brönstrup, M.; Franke, J. Isolation and characterisation of irinans, androstane-type withanolides from Physalis peruviana L. Beilstein J. Org. Chem. 2019, 15, 2003–2012. [Google Scholar] [CrossRef] [Green Version]
- Bernal, C.-A.; Castellanos, L.; Aragón, D.M.; Martínez-Matamoros, D.; Jiménez, C.; Baena, Y.; Ramos, F.A. Peruvioses A to F, sucrose esters from the exudate of Physalis peruviana fruit as α-amylase inhibitors. Carbohydr. Res. 2018, 461, 4–10. [Google Scholar] [CrossRef]
- Dong, B.; An, L.; Yang, X.; Zhang, X.; Zhang, J.; Tuerhong, M.; Jin, D.-Q.; Ohizumi, Y.; Lee, D.; Xu, J. Withanolides from Physalis peruviana showing nitric oxide inhibitory effects and affinities with iNOS. Bioorganic Chem. 2019, 87, 585–593. [Google Scholar] [CrossRef]
- Puente, L.A.; Pinto-Muñoz, C.A.; Castro, E.S.; Cortés, M. Physalis peruviana Linnaeus, the multiple properties of a highly functional fruit: A review. Food Res. Int. 2011, 44, 1733–1740. [Google Scholar] [CrossRef]
- Rey, D.P.; Ospina, L.F.; Aragón, D.M. Inhibitory effects of an extract of fruits of Physalis peruviana on some intestinal carbohydrases. Revista Colombiana de Ciencias Químico-Farmacéuticas 2015, 44, 72–89. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I.; Safwat, G.; Gamal, M.; Megahed, B.M. Chemical Composition and Biological Activity of Physalis peruviana L. Gesunde Pflanzen 2019, 71, 113–122. [Google Scholar] [CrossRef]
- Xu, Y.-M.; Wijeratne, E.K.; Babyak, A.L.; Marks, H.R.; Brooks, A.D.; Tewary, P.; Xuan, L.-J.; Wang, W.-Q.; Sayers, T.J.; Gunatilaka, A.L. Withanolides from aeroponically grown Physalis peruviana and their selective cytotoxicity to prostate cancer and renal carcinoma cells. J. Nat. Prod. 2017, 80, 1981–1991. [Google Scholar] [CrossRef]
- Embaby, H.E.-S.; Mokhtar, S.M. Impact of adding goldenberry (Physalis peruviana L.) on some quality characteristics and bio-functional properties of pasteurized carrot (Daucus carota L.) nectar. J. Food Sci. Technol. 2019, 56, 966–975. [Google Scholar] [CrossRef]
- Legge, A. Notes on the history, cultivation and uses of Physalis peruviana L. J. R. Hortic. Soc. 1974, 99, 310–314. [Google Scholar]
- Fischer, G.; Almanza-Merchán, P.J.; Miranda, D. Importancia y cultivo de la uchuva (Physalis peruviana L.). Revista Brasileira de Fruticultura 2014, 36, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Crosley, L.; Henríquez, J.M.; Parra, F.; Pacheco, P.; Escobar, H.; Parra, C. Rescate del cultivo de Goldenberry (Physalis peruviana) en los Andes del norte de Chile. Idesia 2019, 37, 115–118. [Google Scholar] [CrossRef]
- AOAC—Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International; AOAC: Gaithersburg, MD, USA, 2016. [Google Scholar]
- Soto, E.; Bustos, L.; Simirgiotis, M.J.; Parra, C. Potential of Baccharis alnifolia Meyen & Walpan (Chilka) from northern Chile used as a medicinal infusion. Ciência Rural 2019, 49, 10. [Google Scholar]
- Parra, C.; Soto, E.; León, G.; Salas, C.O.; Heinrich, M.; Echiburú-Chau, C. Nutritional composition, antioxidant activity and isolation of scopoletin from Senecio nutans: Support of ancestral and new uses. Nat. Prod. Res. 2018, 32, 719–722. [Google Scholar] [CrossRef]
- Echiburu-Chau, C.; Pastén, L.; Parra, C.; Bórquez, J.; Mocan, A.; Simirgiotis, M.J. High resolution UHPLC-MS characterization and isolation of main compounds from the antioxidant medicinal plant Parastrephia lucida (Meyen). Saudi Pharm. J. 2017, 25, 1032–1039. [Google Scholar] [CrossRef]
- Belwal, T.; Dhyani, P.; Bhatt, I.D.; Rawal, R.S.; Pande, V. Optimization extraction conditions for improving phenolic content and antioxidant activity in Berberis asiatica fruits using response surface methodology (RSM). Food Chem. 2016, 207, 115–124. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Burton, D.; Parra, F.; López, J.; Muñoz, P.; Escobar, H.; Parra, C. Antioxidant and Antibacterial Capacities of Origanum vulgare L. Essential Oil from the Arid Andean Region of Chile and its Chemical Characterization by GC-MS. Metabolites 2020, 10, 414. [Google Scholar] [CrossRef]
- Parra, C.; Muñoz, P.; Bustos, L.; Parra, F.; Simirgiotis, M.J.; Escobar, H. UHPLC-DAD Characterization of Origanum vulgare L. from Atacama Desert Andean Region and Antioxidant, Antibacterial and Enzyme Inhibition Activities. Molecules 2021, 26, 2100. [Google Scholar] [CrossRef]
- Ngemakwe, P.N.; Remize, F.; Thaoge, M.; Sivakumar, D. Phytochemical and nutritional properties of underutilised fruits in the southern African region. S. Afr. J. Bot. 2017, 113, 137–149. [Google Scholar] [CrossRef]
- Yıldız, G.; İzli, N.; Ünal, H.; Uylaşer, V. Physical and chemical characteristics of goldenberry fruit (Physalis peruviana L.). J. Food Sci. Technol. 2015, 52, 2320–2327. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Vitamin and Mineral Requirements in Human Nutrition; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Rondanelli, M.; Faliva, M.A.; Infantino, V.; Gasparri, C.; Iannello, G.; Perna, S.; Riva, A.; Petrangolini, G.; Tartara, A.; Peroni, G. Copper as Dietary Supplement for Bone Metabolism: A Review. Nutrients 2021, 13, 2246. [Google Scholar] [CrossRef]
- Vincente, A.R.; Manganaris, G.A.; Ortiz, C.M.; Sozzi, G.O.; Crisosto, C.H. Nutritional quality of fruits and vegetables. In Postharvest Handling; Elsevier: Amsterdam, The Netherlands, 2014; pp. 69–122. [Google Scholar]
- Mahmoud, E.A.; Abdelmohsen, S.A.; Elansary, H.O. Husk Cherry: Nutritional Attributes, Bioactive Compounds and Technological Applications. Arab. J. Chem. 2021, 14, 103402. [Google Scholar] [CrossRef]
- Punithavathi, V.R.; Prince, P.S.M.; Kumar, R.; Selvakumari, J. Antihyperglycaemic, antilipid peroxidative and antioxidant effects of gallic acid on streptozotocin induced diabetic Wistar rats. Eur. J. Pharmacol. 2011, 650, 465–471. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.; Yousef, A.I.; Abd El-Twab, S.M.; Reheim, E.S.A.; Ashour, M.B. Gallic acid and p-coumaric acid attenuate type 2 diabetes-induced neurodegeneration in rats. Metab. Brain Dis. 2017, 32, 1279–1286. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.Y.; Sim, G.Y.; Ahn, J.-H. Synthesis of 4-hydroxybenzoic acid derivatives in Escherichia coli. J. Agric. Food Chem. 2020, 68, 9743–9749. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, F.A.; Bertoft, E. Sugar and phenolic acid composition of stored commercial oleaster fruits. J. Food Compos. Anal. 2001, 14, 505–511. [Google Scholar] [CrossRef]
- Alkhalidy, H.; Moore, W.; Wang, Y.; Luo, J.; McMillan, R.P.; Zhen, W.; Zhou, K.; Liu, D. The flavonoid kaempferol ameliorates streptozotocin-induced diabetes by suppressing hepatic glucose production. Molecules 2018, 23, 2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, L.B.; Simirgiotis, M.J.; Parra, C.; Alfaro-Lira, S.; Soto, E.; Echiburú-Chau, C. In Vitro screening for growth inhibition activity on cancer cell lines of northern Chile highlands shrubs. Ciência Rural 2021, 51. [Google Scholar] [CrossRef]
- Bustos, L.; Soto, E.; Parra, F.; Echiburu-Chau, C.; Parra, C. Brewing of a porter craft beer enriched with the plant parastrephia lucida: A promising source of antioxidant compounds. J. Am. Soc. Brew. Chem. 2019, 77, 261–266. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Cimpeanu, C.; Predoi, G. Antioxidant capacity determination in plants and plant-derived products: A review. Oxidative Med. Cell. Longev. 2016, 2016, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.-L.; Chen, S.-G.; Zhao, Y.-Y.; Luo, C.-X.; Li, J.; Gao, Y.-Q. Total phenolic contents of 33 fruits and their antioxidant capacities before and after in vitro digestion. Ind. Crop. Prod. 2014, 57, 150–157. [Google Scholar] [CrossRef]
- Speisky, H.; López-Alarcón, C.; Gómez, M.; Fuentes, J.; Sandoval-Acuña, C. First web-based database on total phenolics and oxygen radical absorbance capacity (ORAC) of fruits produced and consumed within the south Andes region of South America. J. Agric. Food Chem. 2012, 60, 8851–8859. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Jaca, T.; Kambizi, L. Antibacterial properties of some wild leafy vegetables of the Eastern Cape Province, South Africa. J. Med. Plants Res. 2011, 5, 2624–2628. [Google Scholar]
- Çakir, Ö.; Pekmez, M.; Çepni, E.; Candar, B.; Fidan, K. Evaluation of biological activities of Physalis peruviana ethanol extracts and expression of Bcl-2 genes in HeLa cells. Food Sci. Technol. 2014, 34, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Mwambete, K. The In Vitro antimicrobial activity of fruit and leaf crude extracts of Momordica charantia: A Tanzania medicinal plant. Afr. Health Sci. 2009, 9, 34–39. [Google Scholar]
- Sah, A.N.; Juyal, V.; Melkani, A.B. Antimicrobial activity of six different parts of the plant Citrus medica Linn. Pharmacogn. J. 2011, 3, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Felhi, S.; Daoud, A.; Hajlaoui, H.; Mnafgui, K.; Gharsallah, N.; Kadri, A. Solvent extraction effects on phytochemical constituents profiles, antioxidant and antimicrobial activities and functional group analysis of Ecballium elaterium seeds and peels fruits. Food Sci. Technol. 2017, 37, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Ghani, U. Re-exploring promising α-glucosidase inhibitors for potential development into oral anti-diabetic drugs: Finding needle in the haystack. Eur. J. Med. Chem. 2015, 103, 133–162. [Google Scholar] [CrossRef]
- Dong, H.-Q.; Li, M.; Zhu, F.; Liu, F.-L.; Huang, J.-B. Inhibitory potential of trilobatin from Lithocarpus polystachyus Rehd against α-glucosidase and α-amylase linked to type 2 diabetes. Food Chem. 2012, 130, 261–266. [Google Scholar] [CrossRef]
- Picot, M.C.N.; Mahomoodally, M.F. Effects of Aphloia theiformis on key enzymes related to diabetes mellitus. Pharm. Biol. 2017, 55, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Hartini, Y.S.; Setyaningsih, D.; Chang, M.J.V.; Nugrahanti, M. Sambiloto (Andrographis paniculata Nees.) leaf extract activity as an a-Amylase enzyme inhibitor. Pharm. Educ. 2021, 21, 305–308. [Google Scholar] [CrossRef]
- Pinto, M.D.S.; Ranilla, L.G.; Apostolidis, E.; Lajolo, F.M.; Genovese, M.I.; Shetty, K. Evaluation of antihyperglycemia and antihypertension potential of native Peruvian fruits using in vitro models. J. Med. Food 2009, 12, 278–291. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Proximate Composition | PP | Literature [4,26,27] |
|---|---|---|
| Water (g/100 g FW) | 78.95 ± 2.01 | 73.77–78.9 |
| Carbohydrates (g/100 g FW) | 12.66 ± 0.04 | 12.91–19.66 |
| Crude Protein (g/100 g FW) | 1.43 ± 0.36 | 0.05–1.58 |
| Crude Lipid (g/100 g FW) | 0.20 ± 0.07 | 0.15–0.50 |
| Crude Fiber (g/100 g FW | 4.69 ± 0.10 | 4.12–4.90 |
| Ash (g/100 g FW) | 2.09 ± 0.68 | 0.77–2.95 |
| Mineral content | ||
| K (mg/100 g FW) | 256.32 ± 2.20 | 362.03–373.25 |
| Mg (mg/100 g FW) | 20.04 ± 0.08 | 35.96–48.70 |
| Ca (mg/100 g FW) | 17.80 ± 0.12 | 8.00–11.17 |
| Na (mg/100 g FW) | 16.87 ± 0.22 | 8.41–8.78 |
| P (mg/100 g FW) | 94.75 ± 3.65 | 55.3 |
| Mn (mg/100 g FW) | 0.17 ± 0.01 | -- |
| Zn (mg/100 g FW) | 0.15 ± 0.05 | -- |
| Cu (mg/100 g FW) | 0.09 ± 0.01 | 0.26–0.35 |
| Fe (mg/100 g FW) | 0.54 ± 0.05 | 1.2 |

| Peak | Phenolic Compounds | RT | PP * |
|---|---|---|---|
| 5 | Gallic acid | 9.46 | 303.63 ± 35.85 |
| 6 | 4-hydroxybenzoic acid | 9.73 | 43.93 ± 3.45 |
| 7 | Caffeic acid | 9.97 | 15.16 ± 2.22 |
| 8 | Vanillic acid | 10.20 | 9.05 ± 1.11 |
| 10 | Chlorogenic acid | 10.74 | 1.25 ± 0.07 |
| 11 | Ferulic acid | 11.08 | 15.89 ± 0.75 |
| 12 | p-Coumaric acid | 11.42 | 4.29 ± 0.31 |
| 16 | trans-Cinnamic acid | 14.32 | 4.09 ± 0.14 |
| 18 | Luteolin | 18.85 | 15.17 ± 1.85 |
| 20 | Naringenin | 19.48 | 9.47 ± 0.49 |
| 21 | Apigenin | 20.34 | 1.85 ± 0.02 |
| 22 | Kaempferol | 21.34 | 19.57 ± 1.24 |
| Phytochemical Analysis | PP | Literature [4,13,27] |
|---|---|---|
| Total Polyphenol (mg GAE/100 g FW) | 26.24 ± 2.16 | 27.32–7.10 |
| Total Flavonoids (mg QE/100 g FW) | 1.48 ± 0.04 | 1.28 |
| Total tannins (mg Tannic acid/100 g FW) | 1.74 ± 0.33 | 1.10–0.60 |
| Total anthocyanins (µg/100 g FW) | 0.88 ± 0.02 | 1.34 |
| Antioxidant Activity | ||
| FRAP (µmol TE/100 g FW) | 69.58 ± 2.20 | 11.12–8.96 |
| ABTS (µmol TE/100 g FW) | 24.99 ± 1.15 | 3.76–2.60 |
| ORAC (µmol TE/100 g FW) | 3126.82 ± 30.68 | -- |
| Bacterium | MIC 1 (mg/mL) | MIC Kan * (µg/mL) | MBC 2 (mg/mL) | MBC Kan * (µg/mL) |
|---|---|---|---|---|
| Bacillus subtilis (ATCC 6051) | 2.50 | 1.25 | 5.00 | 20.00 |
| Escherichia coli (ATCC 23716) | 2.50 | 5.00 | 5.00 | 10.00 |
| Pseudomonas aeruginosa (ATCC 19429) | 2.50 | 5.00 | 5.00 | 10.00 |
| Salmonella enterica (ATCC 13311) | 1.25 | 2.50 | 5.00 | 5.00 |
| Staphylococcus aureus (ATCC 29737) | 1.25 | 2.50 | 5.00 | 10.00 |
| Agrobacterium tumefaciens (ATCC 19358) | 1.25 | 1.25 | 5.00 | 5.00 |
| Erwinia rhapontici (MK883065) | 5.00 | 1.25 | 5.00 | 2.50 |
| Pantoea agglomerans (MK883087) | 1.25 | 2.50 | 5.00 | 5.00 |
| Pseudomonas syringae (MF547632) | 0.63 | 1.25 | 5.00 | 2.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz, P.; Parra, F.; Simirgiotis, M.J.; Sepúlveda Chavera, G.F.; Parra, C. Chemical Characterization, Nutritional and Bioactive Properties of Physalis peruviana Fruit from High Areas of the Atacama Desert. Foods 2021, 10, 2699. https://doi.org/10.3390/foods10112699
Muñoz P, Parra F, Simirgiotis MJ, Sepúlveda Chavera GF, Parra C. Chemical Characterization, Nutritional and Bioactive Properties of Physalis peruviana Fruit from High Areas of the Atacama Desert. Foods. 2021; 10(11):2699. https://doi.org/10.3390/foods10112699
Chicago/Turabian StyleMuñoz, Patricio, Felipe Parra, Mario J. Simirgiotis, Germán F. Sepúlveda Chavera, and Claudio Parra. 2021. "Chemical Characterization, Nutritional and Bioactive Properties of Physalis peruviana Fruit from High Areas of the Atacama Desert" Foods 10, no. 11: 2699. https://doi.org/10.3390/foods10112699