
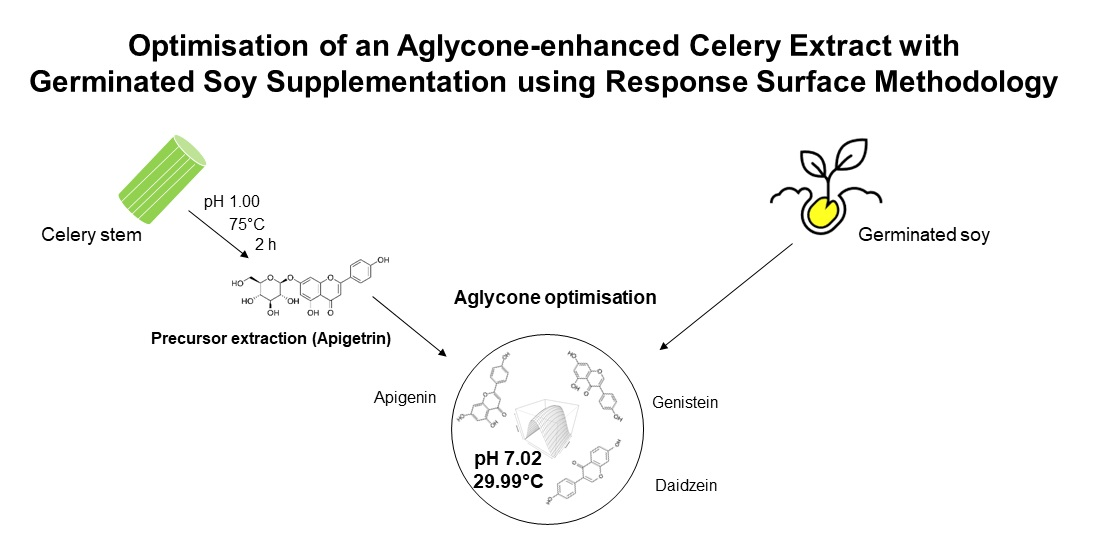
Optimisation of an Aglycone-Enhanced Celery Extract with Germinated Soy Supplementation Using Response Surface Methodology


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Plant Material
2.3. Extraction Procedure
2.4. Experimental Design and Model Verification
2.5. LC-MS/MS
2.6. Data Analysis
3. Results and Discussion
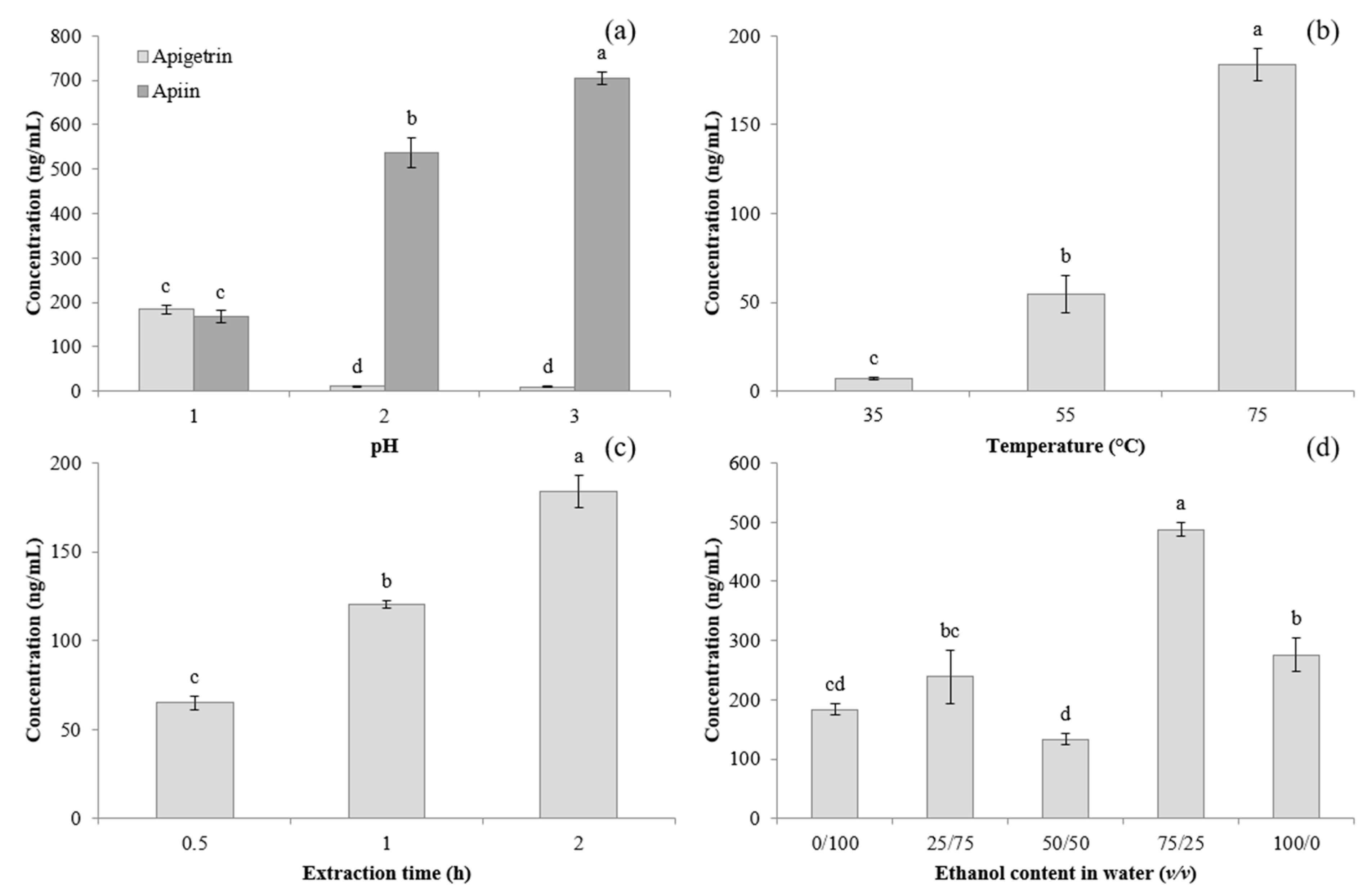
3.1. Extraction Optimisation of Apigetrin
3.2. Extraction Optimisation of Apigenin and Soy Aglycones
3.2.1. Validation of the Experimental Design
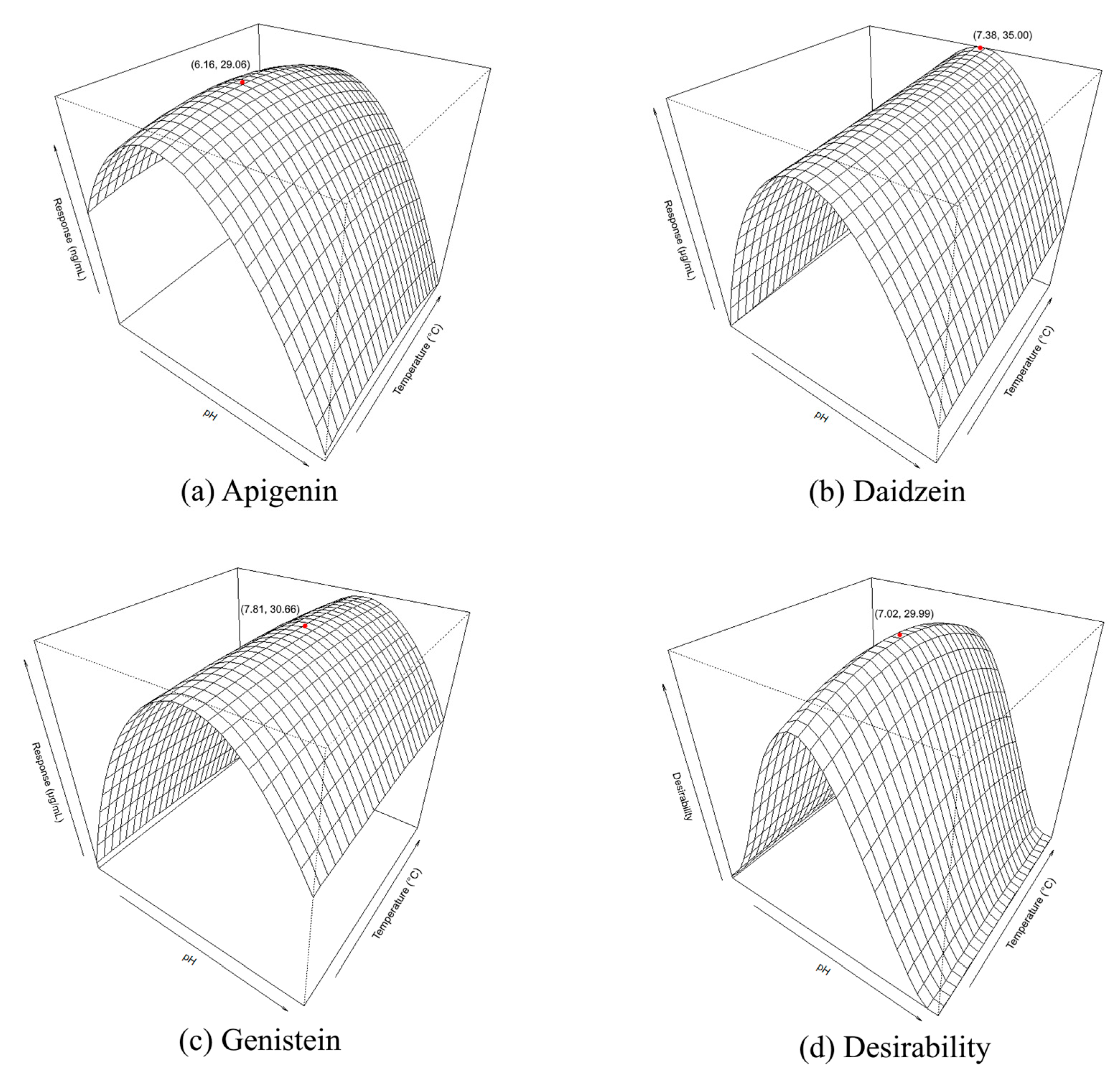
3.2.2. Analysis of the Response Surface
3.2.3. Optimisation Responses and Verification of the Model
3.2.4. Quantitation of Bioactive Compounds by LC-MS/MS Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, M.-Y.; Hou, X.-L.; Wang, F.; Tan, G.-F.; Xu, Z.-S.; Xiong, A.-S. Advances in the research of celery, an important Apiaceae vegetable crop. Crit. Rev. Biotechnol. 2018, 38, 172–183. [Google Scholar] [CrossRef]
- Che, D.N.; Cho, B.O.; Shin, J.Y.; Kang, H.J.; Kim, J.S.; Choi, J.; Jang, S.I. Anti-atopic dermatitis effects of hydrolyzed celery extract in mice. J. Food Biochem. 2020, 44, e13198. [Google Scholar] [CrossRef]
- Hu, J.; Ma, W.; Li, N.; Wang, K.-J. Antioxidant and anti-inflammatory flavonoids from the flowers of Chuju, a medical cultivar of Chrysanthemum morifolim Ramat. J. Mex. Chem. Soc. 2017, 61, 282–289. [Google Scholar] [CrossRef]
- Ali, F.; Rahul; Naz, F.; Jyoti, S.; Siddique, Y.H. Health functionality of apigenin: A review. Int. J. Food Prop. 2017, 20, 1197–1238. [Google Scholar] [CrossRef]
- Tan, G.-F.; Ma, J.; Zhang, X.-Y.; Xu, Z.-S.; Xiong, A.-S. AgFNS overexpression increase apigenin and decrease anthocyanins in petioles of transgenic celery. Plant Sci. 2017, 263, 31–38. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, M.M.; Chen, P.L.; Cao, Y.Y.; Tan, X.L. Optimization of ultrasonic-assisted enzymatic hydrolysis for the extraction of luteolin and apigenin from celery. J. Food Sci. 2011, 76, C680–C685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q. High-yield production of apigenin from celery with ultrasonic-assisted β-glucosidase hydrolysis. Asian J. Chem. 2015, 27, 1501–1505. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Riedl, K.M.; Schwartz, S.J. Effects of food formulation and thermal processing on flavones in celery and chamomile. Food Chem. 2013, 141, 1406–1411. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Yu, L.; Xu, S.; Gu, W.; Zhu, W. Apigenin accumulation and expression analysis of apigenin biosynthesis relative genes in celery. Sci. Hortic. 2014, 165, 218–224. [Google Scholar] [CrossRef]
- Cai, J.-S.; Feng, J.-Y.; Ni, Z.-J.; Ma, R.-H.; Thakur, K.; Wang, S.; Hu, F.; Zhang, J.-G.; Wei, Z.-J. An update on the nutritional, functional, sensory characteristics of soy products, and applications of new processing strategies. Trends Food Sci. Technol. 2021, 112, 676–689. [Google Scholar] [CrossRef]
- Garg, S.; Lule, V.K.; Malik, R.K.; Tomar, S.K. Soy bioactive components in functional perspective: A review. Int. J. Food Prop. 2016, 19, 2550–2574. [Google Scholar] [CrossRef] [Green Version]
- Izumi, T.; Piskula, M.K.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kubota, Y.; Kikuchi, M. Soy isoflavone aglycones are absorbed faster and in higher amounts than their glucosides in humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [Green Version]
- Rüfer, C.E.; Bub, A.; Möseneder, J.; Winterhalter, P.; Stürtz, M.; Kulling, S.E. Pharmacokinetics of the soybean isoflavone daidzein in its aglycone and glucoside form: A randomized, double-blind, crossover study. Am. J. Clin. Nutr. 2008, 87, 1314–1323. [Google Scholar] [CrossRef] [Green Version]
- Handa, C.; Couto, U.; Vicensoti, A.; Georgetti, S.; Ida, E. Optimisation of soy flour fermentation parameters to produce β-glucosidase for bioconversion into aglycones. Food Chem. 2014, 152, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Yoshiara, L.Y.; Madeira, T.B.; De Camargo, A.C.; Shahidi, F.; Ida, E.I. Multistep optimization of β-glucosidase extraction from germinated soybeans (Glycine max L. Merril) and recovery of isoflavone aglycones. Foods 2018, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, M.; Obata, A. β-Glucosidases from soybeans hydrolyze daidzin and genistin. J. Food Sci. 1993, 58, 144–147. [Google Scholar] [CrossRef]
- Hsieh, M.-C.; Graham, T.L. Partial purification and characterization of a soybean β-glucosidase with high specific activity towards isoflavone conjugates. Phytochemistry 2001, 58, 995–1005. [Google Scholar] [CrossRef]
- Taghinezhad, E.; Kaveh, M.; Szumny, A. Optimization and prediction of the drying and quality of turnip slices by convective-infrared dryer under various pretreatments by RSM and ANFIS methods. Foods 2021, 10, 284. [Google Scholar] [CrossRef] [PubMed]
- Yıkmış, S.; Aksu, F.; Altunatmaz, S.S.; Çöl, B.G. Ultrasound Processing of Vinegar: Modelling the Impact on Bioactives and Other Quality Factors. Foods 2021, 10, 1703. [Google Scholar] [CrossRef]
- Bezerra, M.A.; Santelli, R.E.; Oliveira, E.P.; Villar, L.S.; Escaleira, L.A. Response surface methodology (RSM) as a tool for optimization in analytical chemistry. Talanta 2008, 76, 965–977. [Google Scholar] [CrossRef]
- Ribeiro, M.; Mandarino, J.; CarrÃO-Panizzi, M.; Oliveira, M.; Campo, C.; Nepomuceno, A.; Ida, E. β-glucosidase activity and isoflavone content in germinated soybean radicles and cotyledons. J. Food Biochem. 2006, 30, 453–465. [Google Scholar] [CrossRef]
- Suzuki, H.; Takahashi, S.; Watanabe, R.; Fukushima, Y.; Fujita, N.; Noguchi, A.; Yokoyama, R.; Nishitani, K.; Nishino, T.; Nakayama, T. An isoflavone conjugate-hydrolyzing β-glucosidase from the roots of soybean (Glycine max) seedlings: Purification, gene cloning, phylogenetics, and cellular localization. J. Biol. Chem. 2006, 281, 30251–30259. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Gao, J.; Yang, A.; Chen, H. The ultrasound-treated soybean seeds improve edibility and nutritional quality of soybean sprouts. Food Res. Int. 2015, 77, 704–710. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Horikoshi, M.; Tang, Y. Ggfortify: Data Visualization Tools for Statistical Analysis Results; 2016. Available online: https://CRAN.R-project.org/package=ggfortify (accessed on 25 June 2021).
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Harbourne, N.; Jacquier, J.C.; O’Riordan, D. Optimisation of the extraction and processing conditions of chamomile (Matricaria chamomilla L.) for incorporation into a beverage. Food Chem. 2009, 115, 15–19. [Google Scholar] [CrossRef]
- Hossain, M.B.; Brunton, N.P.; Patras, A.; Tiwari, B.; O’donnell, C.; Martin-Diana, A.B.; Barry-Ryan, C. Optimization of ultrasound assisted extraction of antioxidant compounds from marjoram (Origanum majorana L.) using response surface methodology. Ultrason. Sonochem. 2012, 19, 582–590. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, Z.; Huang, Y.; Wen, X.; Wu, Y.; Zhao, Y.; Ni, Y. Extraction, purification, and hydrolysis behavior of apigenin-7-O-Glucoside from Chrysanthemum morifolium tea. Molecules 2018, 23, 2933. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Zhou, J.; Wu, Z.; Tan, G.; Li, H.; Mei, Q.; Qian, Z. Analysis of antioxidants in Chrysanthemum indici flos by online gradient extraction and HPLC-FRAP. Anal. Methods 2021, 13, 2283–2289. [Google Scholar] [CrossRef]
- Dent, M.; Dragović-Uzelac, V.; Penić, M.; Bosiljkov, T.; Levaj, B. The effect of extraction solvents, temperature and time on the composition and mass fraction of polyphenols in Dalmatian wild sage (Salvia officinalis L.) extracts. Food Technol. Biotechnol. 2013, 51, 84–91. [Google Scholar]
- Stanojević, L.; Stanković, M.; Nikolić, L.; Nikolić, V. The influence of the operation conditions and the extraction techniques on the yield, kinetics and the composition of ethanol extracts of Hieracium pilosella L. Chem. Ind. Chem. Eng. Q. 2007, 13, 199–204. [Google Scholar] [CrossRef]
- Batool, K.; Bibi, A. Microbial β-Glucosidases: Screening, Characterization, Cloning and Applications. J. Appl. Microbiol. 2017, 5, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Ungar, Y.; Osundahunsi, O.F.; Shimoni, E. Thermal stability of genistein and daidzein and its effect on their antioxidant activity. J. Agric. Food. Chem. 2003, 51, 4394–4399. [Google Scholar] [CrossRef]
- Da Silva, B.; Kupski, L.; Badiale-Furlong, E. Central composite design-desirability function approach for optimum ultrasound-assisted extraction of daidzein and genistein from soybean and their antimycotoxigenic potential. Food Anal. Methods 2019, 12, 258–270. [Google Scholar] [CrossRef]
- Derringer, G.; Suich, R. Simultaneous optimization of several response variables. J. Qual. Technol. 1980, 12, 214–219. [Google Scholar] [CrossRef]
- Islam, A.; Islam, M.S.; Uddin, M.N.; Hasan, M.M.I.; Akanda, M.R. The potential health benefits of the isoflavone glycoside genistin. Arch. Pharmacal Res. 2020, 43, 395–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Run | Parameters | Responses | ||||||
|---|---|---|---|---|---|---|---|---|
| X1 (pH) | X2 (T, °C) | Y1, Apigenin (ng/mL Extract) | Y2, Daidzein (µg/mL Extract) | Y3, Genistein (µg/mL Extract) | ||||
| Exp | Pred | Exp | Pred | Exp | Pred | |||
| 1 | 11.0 | 30.0 | 2.86 | −3.76 | 2.38 | 3.91 | 9.66 | 19.32 |
| 2 | 7.0 | 30.0 | 38.87 | 35.97 | 6.37 | 7.83 | 22.97 | 30.96 |
| 3 | 4.2 | 33.5 | 26.82 | 27.16 | 5.32 | 5.12 | 13.77 | 15.14 |
| 4 | 7.0 | 35.0 | 33.29 | 30.79 | 7.37 | 8.13 | 26.97 | 30.38 |
| 5 | 7.0 | 25.0 | 37.47 | 33.48 | 6.86 | 7.58 | 27.56 | 30.28 |
| 6 | 7.0 | 30.0 | 34.12 | 35.97 | 8.32 | 7.83 | 32.74 | 30.96 |
| 7 | 3.0 | 30.0 | 19.54 | 19.67 | 2.50 | 2.45 | 7.22 | 3.68 |
| 8 | 9.8 | 26.5 | 6.34 | 12.50 | 7.04 | 5.76 | 33.62 | 26.13 |
| 9 | 4.2 | 26.5 | 28.11 | 29.50 | 5.05 | 4.88 | 13.78 | 15.63 |
| 10 | 9.8 | 33.5 | 5.92 | 11.03 | 7.61 | 6.30 | 34.75 | 26.77 |
| 11 | 7.0 | 30.0 | 31.95 | 35.97 | 7.72 | 7.83 | 30.49 | 30.96 |
| 12 | 7.0 | 30.0 | 38.94 | 35.97 | 8.92 | 7.83 | 37.65 | 30.96 |
| Responses | Apigenin (Y1) | Daidzein (Y2) | Genistein (Y3) |
|---|---|---|---|
| Model Parameters | |||
| Second-order polynomial equation | −154.82 + 20.93 X1 + 8.79 X2 + 0.02 X1X2 − 1.75 X12 − 0.15 X22 | −6.92 + 4.02 X1 − 0.05 X2 + 0.01 X1X2 − 0.29 X12 + 0.0009 X22 | −59.46 + 18.13 X1 + 1.33 X2 + 0.03 X1X2 − 1.22 X12 − 0.03 X22 |
| R2 | 0.915 | 0.788 | 0.706 |
| R2 (adjusted) | 0.844 | 0.611 | 0.461 |
| Lack of fit | 0.161 | 0.304 | 0.263 |
| p | |||
| Model | 0.004 | 0.048 | 0.115 |
| X1 (pH) | 0.055 | 0.115 | 0.209 |
| X2 (T, °C) | 0.424 | 0.984 | 0.932 |
| X1X2 | 0.937 | 0.910 | 0.944 |
| X12 | 0.001 | 0.004 | 0.019 |
| X22 | 0.396 | 0.984 | 0.921 |
| Responses | p | Predicted Value | Experimental Value |
|---|---|---|---|
| Apigenin (Y1) | 0.467 | 35.97 | 38.97 ± 5.83 |
| Daidzein (Y2) | 0.201 | 7.83 | 6.90 ± 0.86 |
| Genistein (Y3) | 0.499 | 30.96 | 27.90 ± 6.49 |
| Compound | Concentration | ||
|---|---|---|---|
| Celery Extract (75% ETOH) | Celery Extract (0% ETOH) | Germinated Soy-Supplemented Celery Extract | |
| Apigenin (ng/mL) | 3.17 ± 0.16 | 0.95 ± 0.05 | 38.97 ± 5.83 |
| Apigetrin (ng/mL) | 487.78 ± 11.58 | 183.88 ± 8.94 | 7.85 ± 1.25 |
| Apiin (ng/mL) | 222.89 ± 11.78 | 167.36 ± 13.58 | 6.83 ± 0.75 |
| Genistein (µg/mL) | - | - | 27.90 ± 6.49 |
| Daidzein (µg/mL) | - | - | 6.90 ± 0.86 |
| Genistin (µg/mL) | - | - | 3.34 ± 0.37 |
| Daidzin (µg/mL) | - | - | 20.18 ± 1.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lau, H.; Koh, H.M.; Dayal, H.; Ren, Y.; Li, S.F.Y. Optimisation of an Aglycone-Enhanced Celery Extract with Germinated Soy Supplementation Using Response Surface Methodology. Foods 2021, 10, 2505. https://doi.org/10.3390/foods10102505
Lau H, Koh HM, Dayal H, Ren Y, Li SFY. Optimisation of an Aglycone-Enhanced Celery Extract with Germinated Soy Supplementation Using Response Surface Methodology. Foods. 2021; 10(10):2505. https://doi.org/10.3390/foods10102505
Chicago/Turabian StyleLau, Hazel, Hui Min Koh, Hiranya Dayal, Yi Ren, and Sam Fong Yau Li. 2021. "Optimisation of an Aglycone-Enhanced Celery Extract with Germinated Soy Supplementation Using Response Surface Methodology" Foods 10, no. 10: 2505. https://doi.org/10.3390/foods10102505