Review of Virus Inactivation by Visible Light


Abstract
:1. Introduction
2. Materials and Methods
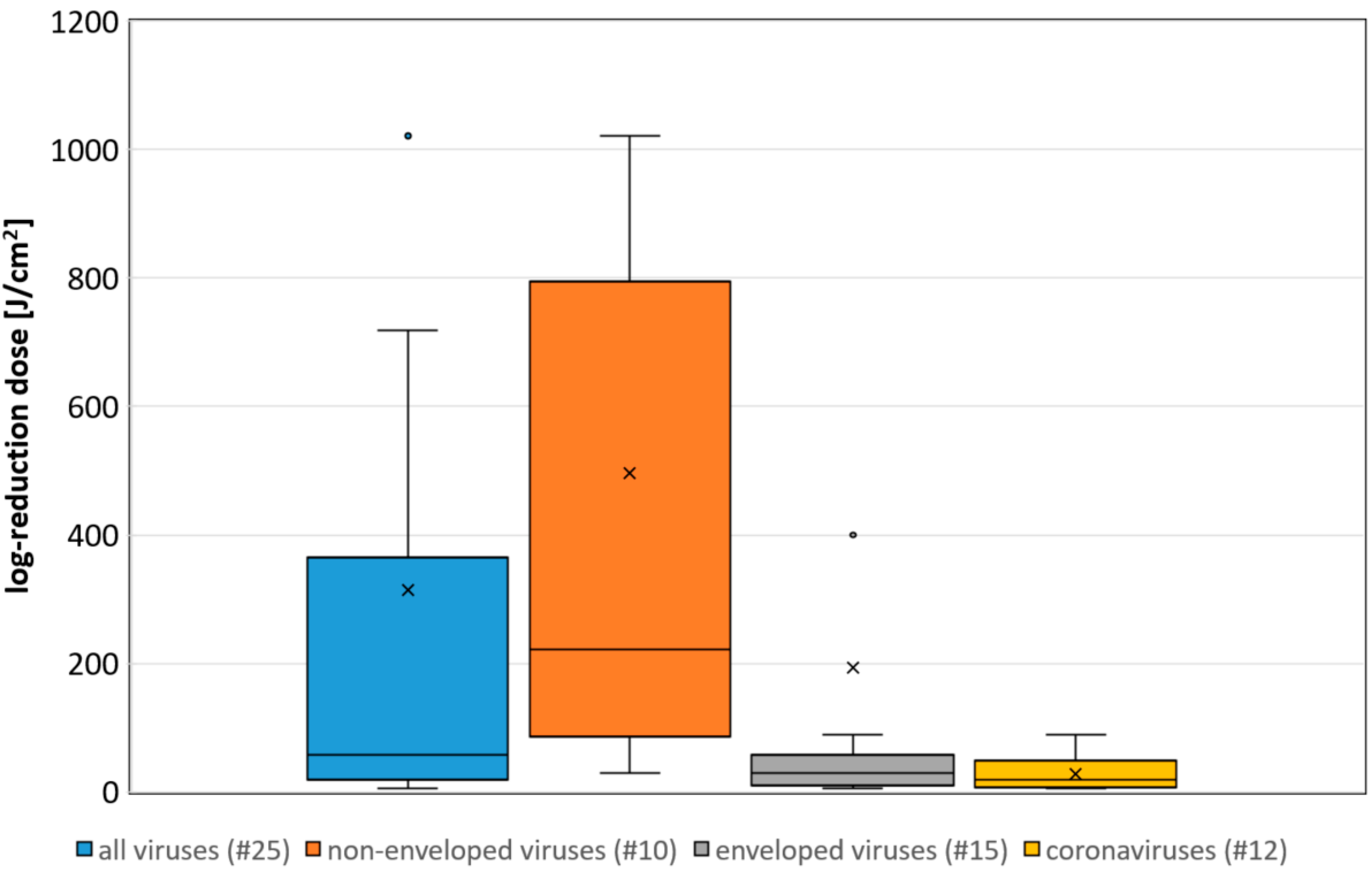
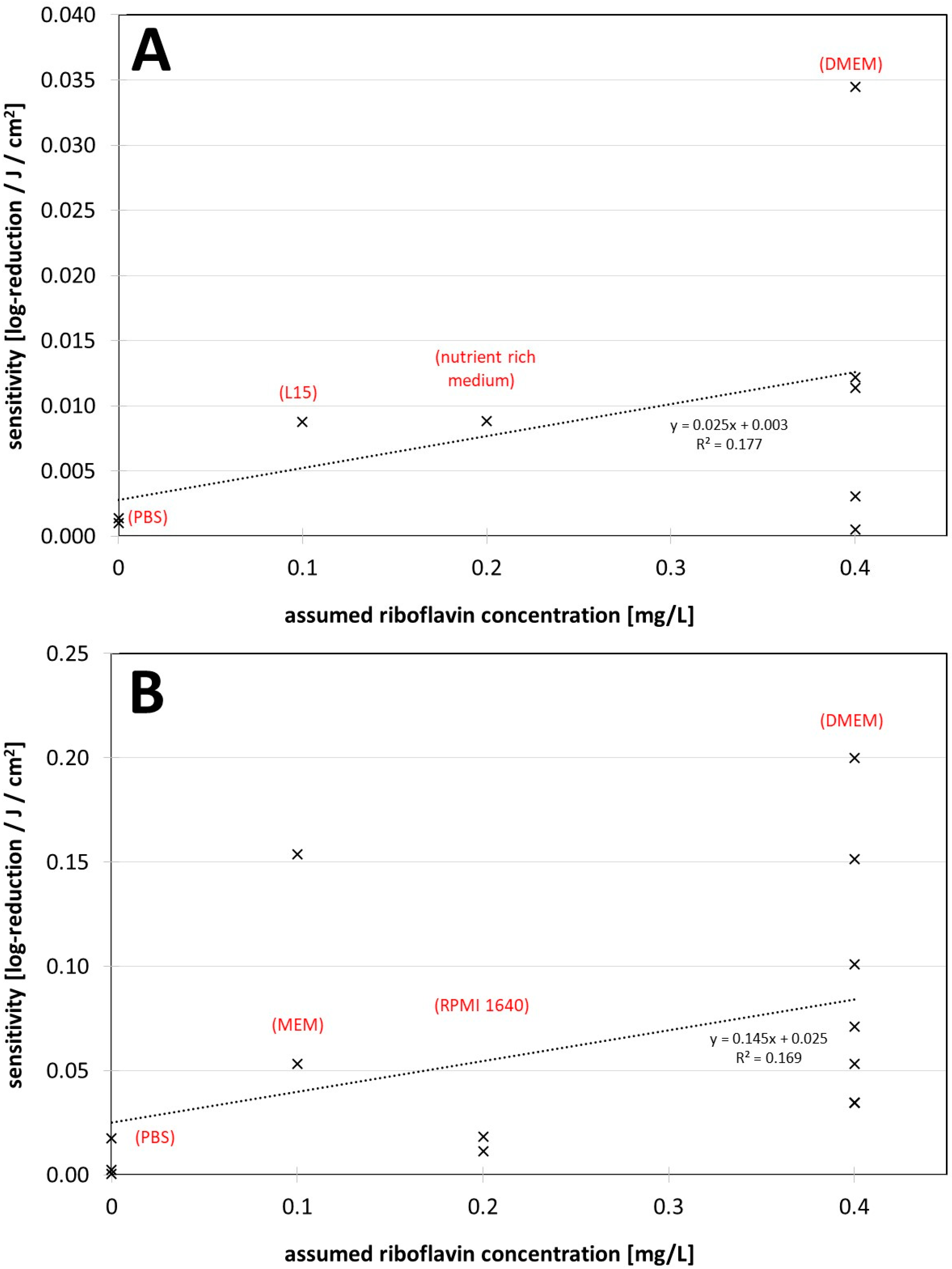
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coronavirus Resource Center. COVID-19 Dashboard: (Global Map). Available online: https://coronavirus.jhu.edu/map.html (accessed on 3 January 2022).
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and their inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampf, G.; Voss, A.; Scheithauer, S. Inactivation of coronaviruses by heat. J. Hosp. Infect. 2020, 105, 348–349. [Google Scholar] [CrossRef] [PubMed]
- Kratzel, A.; Todt, D.; V’kovski, P.; Steiner, S.; Gultom, M.; Thao, T.T.N.; Ebert, N.; Holwerda, M.; Steinmann, J.; Niemeyer, D.; et al. Inactivation of Severe Acute Respiratory Syndrome Coronavirus 2 by WHO-Recommended Hand Rub Formulations and Alcohols. Emerg. Infect. Dis. 2020, 26, 1592–1595. [Google Scholar] [CrossRef] [PubMed]
- Hessling, M.; Hönes, K.; Vatter, P.; Lingenfelder, C. Ultraviolet irradiation doses for coronavirus inactivation—Review and analysis of coronavirus photoinactivation studies. GMS Hyg. Infect. Control 2020, 15, 8. [Google Scholar] [CrossRef]
- Hessling, M.; Hoenes, K.; Lingenfelder, C. Selection of parameters for thermal coronavirus inactivation—A data-based recommendation. GMS Hyg. Infect. Control 2020, 15, 16. [Google Scholar] [CrossRef]
- Ashkenazi, H.; Malik, Z.; Harth, Y.; Nitzan, Y. Eradication of Propionibacterium acnes by its endogenic porphyrins after illumination with high intensity blue light. FEMS Immunol. Med. Microbiol. 2003, 35, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guffey, J.S.; Wilborn, J. In vitro bactericidal effects of 405-nm and 470-nm blue light. Photomed. Laser Surg. 2006, 24, 684–688. [Google Scholar] [CrossRef]
- Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Woolsey, G. High-intensity narrow-spectrum light inactivation and wavelength sensitivity of Staphylococcus aureus. FEMS Microbiol. Lett. 2008, 285, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Feuerstein, O.; Ginsburg, I.; Dayan, E.; Veler, D.; Weiss, E.I. Mechanism of visible light phototoxicity on Porphyromonas gingivalis and Fusobacterium nucleatum. Photochem. Photobiol. 2005, 81, 1186–1189. [Google Scholar] [CrossRef]
- Amin, R.M.; Bhayana, B.; Hamblin, M.R.; Dai, T. Antimicrobial blue light inactivation of Pseudomonas aeruginosa by photo-excitation of endogenous porphyrins: In vitro and in vivo studies. Lasers Surg. Med. 2016, 48, 562–568. [Google Scholar] [CrossRef] [Green Version]
- Plavskii, V.Y.; Mikulich, A.V.; Tretyakova, A.I.; Leusenka, I.A.; Plavskaya, L.G.; Kazyuchits, O.A.; Dobysh, I.I.; Krasnenkova, T.P. Porphyrins and flavins as endogenous acceptors of optical radiation of blue spectral region determining photoinactivation of microbial cells. J. Photochem. Photobiol. B 2018, 183, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Cieplik, F.; Spath, A.; Leibl, C.; Gollmer, A.; Regensburger, J.; Tabenski, L.; Hiller, K.-A.; Maisch, T.; Schmalz, G. Blue light kills Aggregatibacter actinomycetemcomitans due to its endogenous photosensitizers. Clin. Oral Investig. 2014, 18, 1763–1769. [Google Scholar] [CrossRef] [PubMed]
- Hessling, M.; Spellerberg, B.; Hoenes, K. Photoinactivation of bacteria by endogenous photosensitizers and exposure to visible light of different wavelengths—A review on existing data. FEMS Microbiol. Lett. 2016, 364, fnw270. [Google Scholar] [CrossRef]
- Tomb, R.M.; White, T.A.; Coia, J.E.; Anderson, J.G.; MacGregor, S.J.; Maclean, M. Review of the Comparative Susceptibility of Microbial Species to Photoinactivation Using 380-480 nm Violet-Blue Light. Photochem. Photobiol. 2018, 94, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Kleinpenning, M.M.; Smits, T.; Frunt, M.H.A.; van Erp, P.E.J.; van de Kerkhof, P.C.M.; Gerritsen, R.M.J.P. Clinical and histological effects of blue light on normal skin. Photodermatol. Photoimmunol. Photomed. 2010, 26, 16–21. [Google Scholar] [CrossRef]
- McDonald, R.S.; Gupta, S.; Maclean, M.; Ramakrishnan, P.; Anderson, J.G.; Macgregor, S.J.; Meek, R.M.D.; Grant, M.H. 405 nm Light exposure of osteoblasts and inactivation of bacterial isolates from arthroplasty patients: Potential for new disinfection applications? Eur. Cell. Mater. 2013, 25, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Dong, J.; Yin, H.; Zhang, G. Blue light therapy to treat candida vaginitis with comparisons of three wavelengths: An in vitro study. Lasers Med. Sci. 2020, 35, 1329–1339. [Google Scholar] [CrossRef]
- Liebmann, J.; Born, M.; Kolb-Bachofen, V. Blue-light irradiation regulates proliferation and differentiation in human skin cells. J. Investig. Dermatol. 2010, 130, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Bumah, V.V.; Masson-Meyers, D.S.; Awosika, O.; Zacharias, S.; Enwemeka, C.S. The viability of human cells irradiated with 470-nm light at various radiant energies in vitro. Lasers Med. Sci. 2021, 36, 1661–1670. [Google Scholar] [CrossRef]
- Makdoumi, K.; Hedin, M.; Bäckman, A. Different photodynamic effects of blue light with and without riboflavin on methicillin-resistant Staphylococcus aureus (MRSA) and human keratinocytes in vitro. Lasers Med. Sci. 2019, 34, 1799–1805. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, P.; Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Grant, M.H. Differential sensitivity of osteoblasts and bacterial pathogens to 405-nm light highlighting potential for decontamination applications in orthopedic surgery. J. Biomed. Opt. 2014, 19, 105001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, T.; Gupta, A.; Huang, Y.-Y.; Yin, R.; Murray, C.K.; Vrahas, M.S.; Sherwood, M.E.; Tegos, G.P.; Hamblin, M.R. Blue light rescues mice from potentially fatal Pseudomonas aeruginosa burn infection: Efficacy, safety, and mechanism of action. Antimicrob. Agents Chemother. 2013, 57, 1238–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhu, Y.; Gupta, A.; Huang, Y.; Murray, C.K.; Vrahas, M.S.; Sherwood, M.E.; Baer, D.G.; Hamblin, M.R.; Dai, T. Antimicrobial blue light therapy for multidrug-resistant Acinetobacter baumannii infection in a mouse burn model: Implications for prophylaxis and treatment of combat-related wound infections. J. Infect. Dis. 2014, 209, 1963–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grzelak, A.; Rychlik, B.; Bartosz, G. Light-dependent generation of reactive oxygen species in cell culture media. Free Radic. Biol. Med. 2001, 30, 1418–1425. [Google Scholar] [CrossRef]
- Ruane, P.H.; Edrich, R.; Gampp, D.; Keil, S.D.; Leonard, R.L.; Goodrich, R.P. Photochemical inactivation of selected viruses and bacteria in platelet concentrates using riboflavin and light. Transfusion 2004, 44, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Marschner, S.; Goodrich, R. Pathogen Reduction Technology Treatment of Platelets, Plasma and Whole Blood Using Riboflavin and UV Light. Transfus. Med. Hemother. 2011, 38, 8–18. [Google Scholar] [CrossRef]
- Faddy, H.M.; Prow, N.A.; Fryk, J.J.; Hall, R.A.; Keil, S.D.; Goodrich, R.P.; Marks, D.C. The effect of riboflavin and ultraviolet light on the infectivity of arboviruses. Transfusion 2015, 55, 824–831. [Google Scholar] [CrossRef]
- Faddy, H.M.; Fryk, J.J.; Watterson, D.; Young, P.R.; Modhiran, N.; Muller, D.A.; Keil, S.D.; Goodrich, R.P.; Marks, D.C. Riboflavin and ultraviolet light: Impact on dengue virus infectivity. Vox Sang. 2016, 111, 235–241. [Google Scholar] [CrossRef]
- Elikaei, A.; Hosseini, S.M.; Sharifi, Z. Inactivation of model viruses and bacteria in human fresh frozen plasma using riboflavin and long wave ultraviolet rays. Iran. J. Microbiol. 2017, 9, 50–54. [Google Scholar]
- Callahan, S.M.; Wonganan, P.; Obenauer-Kutner, L.J.; Sutjipto, S.; Dekker, J.D.; Croyle, M.A. Controlled inactivation of recombinant viruses with vitamin B2. J. Virol. Methods 2008, 148, 132–145. [Google Scholar] [CrossRef] [Green Version]
- Keil, S.D.; Ragan, I.; Yonemura, S.; Hartson, L.; Dart, N.K.; Bowen, R. Inactivation of severe acute respiratory syndrome coronavirus 2 in plasma and platelet products using a riboflavin and ultraviolet light-based photochemical treatment. Vox Sang. 2020, 115, 495–501. [Google Scholar] [CrossRef]
- Zhou, Z.-Y.; Bi, X.-X. Experimental studies on the inactivation of HBV in blood via riboflavin photochemical treatment. Exp. Ther. Med. 2017, 13, 222–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulbecco, R.; Freeman, G. Plaque production by the polyoma virus. Virology 1959, 8, 396–397. [Google Scholar] [CrossRef]
- Moore, G.E. Culture of Normal Human Leukocytes. JAMA 1967, 199, 519. [Google Scholar] [CrossRef] [PubMed]
- Eagle, H. Propagation in a fluid medium of a human epidermoid carcinoma, strain KB. Proc. Soc. Exp. Biol. Med. 1955, 89, 362–364. [Google Scholar] [CrossRef]
- Martin, C.B.; Wilfong, E.; Ruane, P.; Goodrich, R.; Platz, M. An action spectrum of the riboflavin-photosensitized inactivation of Lambda phage. Photochem. Photobiol. 2005, 81, 474–480. [Google Scholar] [CrossRef]
- Terrosi, C.; Anichini, G.; Docquier, J.D.; Gori Savellini, G.; Gandolfo, C.; Pavone, F.S.; Cusi, M.G. Efficient Inactivation of SARS-CoV-2 and Other RNA or DNA Viruses with Blue LED Light. Pathogens 2021, 10, 1590. [Google Scholar] [CrossRef]
- Tomb, R.M.; Maclean, M.; Coia, J.E.; Graham, E.; McDonald, M.; Atreya, C.D.; MacGregor, S.J.; Anderson, J.G. New Proof-of-Concept in Viral Inactivation: Virucidal Efficacy of 405 nm Light Against Feline Calicivirus as a Model for Norovirus Decontamination. Food Environ. Virol. 2016, 9, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Tomb, R.M.; Maclean, M.; Herron, P.R.; Hoskisson, P.A.; MacGregor, S.J.; Anderson, J.G. Inactivation of Streptomyces phage C31 by 405 nm light: Requirement for exogenous photosensitizers? Bacteriophage 2014, 4, e32129. [Google Scholar] [CrossRef] [Green Version]
- Ho, D.T.; Kim, A.; Kim, N.; Roh, H.J.; Chun, W.-K.; Lee, Y.; Kim, D.-H. Effect of blue light emitting diode on viral hemorrhagic septicemia in olive flounder (Paralichthys olivaceus). Aquaculture 2020, 521, 735019. [Google Scholar] [CrossRef]
- Rathnasinghe, R.; Jangra, S.; Miorin, L.; Schotsaert, M.; Yahnke, C.; Garcίa-Sastre, A. The virucidal effects of 405 nm visible light on SARS-CoV-2 and influenza A virus. Sci. Rep. 2021, 11, 19470. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, D.; Kuis, R.; Perez, R.; Basaldua, I.; Burkins, P.; Marcano, A.; Johnson, A. Oxygen-dependent laser inactivation of murine norovirus using visible light lasers. Virol. J. 2018, 15, 117. [Google Scholar] [CrossRef]
- Tsugita, A.; Okada, Y.; Uehara, K. Photosensitized inactivation of ribonucleic acids in the presence of riboflavin. Biochim. Biophys. Acta—Nucleic Acids Protein Synth. 1965, 103, 360–363. [Google Scholar] [CrossRef]
- Kingsley, D.H.; Perez-Perez, R.E.; Boyd, G.; Sites, J.; Niemira, B.A. Evaluation of 405-nm monochromatic light for inactivation of Tulane virus on blueberry surfaces. J. Appl. Microbiol. 2018, 124, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Skinner, H.H.; Bradish, C.J. Exposure to light as a source of error in the estimation of the infectivity of virus suspensions. J. Gen. Microbiol. 1954, 10, 377–397. [Google Scholar] [CrossRef] [Green Version]
- Appleyard, G. The photosensitivity of Semliki Forest and other viruses. J. Gen. Virol. 1967, 1, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Stasko, N.; Kocher, J.F.; Annas, A.; Henson, I.; Seitz, T.S.; Miller, J.M.; Arwood, L.; Roberts, R.C.; Womble, T.M.; Keller, E.G.; et al. Visible blue light inhibits infection and replication of SARS-CoV-2 at doses that are well-tolerated by human respiratory tissue. Sci. Rep. 2021, 11, 20595. [Google Scholar] [CrossRef]
- Cutchins, E.C.; Dayhuff, T.R. Photoinactivation of measles virus. Virology 1962, 17, 420–425. [Google Scholar] [CrossRef]
- de Santis, R.; Luca, V.; Näslund, J.; Ehmann, R.K.; de Angelis, M.; Lundmark, E.; Nencioni, L.; Faggioni, G.; Fillo, S.; Amatore, D.; et al. Rapid inactivation of SARS-CoV-2 with LED irradiation of visible spectrum wavelengths. J. Photochem. Photobiol. 2021, 8, 100082. [Google Scholar] [CrossRef]
- Biasin, M.; Strizzi, S.; Bianco, A.; Macchi, A.; Utyro, O.; Pareschi, G.; Loffreda, A.; Cavalleri, A.; Lualdi, M.; Trabattoni, D.; et al. UV-A and UV-B Can Neutralize SARS-CoV-2 Infectivity. medRxiv 2021. [Google Scholar] [CrossRef]
- Gardner, A.; Ghosh, S.; Dunowska, M.; Brightwell, G. Virucidal Efficacy of Blue LED and Far-UVC Light Disinfection against Feline Infectious Peritonitis Virus as a Model for SARS-CoV-2. Viruses 2021, 13, 1436. [Google Scholar] [CrossRef] [PubMed]
- Zavestovskaya, I.N.; Guschin, V.A.; Nikiforova, M.A.; Siniavin, A.E.; Russu, L.I.; Cheshev, E.A.; Koromyslov, A.L.; Tupitsyn, I.M.; Fronya, A.A.; Grigoryeva, M.S. Experimental investigation of the effect ov UVA radiation on the coronavirus infective properties. Bull. Lebedev Phys. Inst. 2021, 48, 195–199. [Google Scholar] [CrossRef]
- Enwemeka, C.S.; Bumah, V.V.; Mokili, J.L. Pulsed blue light inactivates two strains of human coronavirus. J. Photochem. Photobiol. B 2021, 222, 112282. [Google Scholar] [CrossRef]
- Lau, B.; Becher, D.; Hessling, M. High Intensity Violet Light (405 nm) Inactivates Coronaviruses in Phosphate Buffered Saline (PBS) and on Surfaces. Photonics 2021, 8, 414. [Google Scholar] [CrossRef]
- Zupin, L.; Caracciolo, I.; Tricarico, P.M.; Ottaviani, G.; D’Agaro, P.; Crovella, S. Photobiomodulation therapy reduces viral load and cell death in ZIKV-infected glioblastoma cell line. Lasers Med. Sci. 2018, 33, 2011–2013. [Google Scholar] [CrossRef] [PubMed]
- Zupin, L.; Caracciolo, I.; Tricarico, P.M.; Ottaviani, G.; D’Agaro, P.; Crovella, S. Antiviral properties of blue laser in an in vitro model of HSV-1 infection. Microbiol. Immunol. 2018, 62, 477–479. [Google Scholar] [CrossRef] [Green Version]
- Vatter, P.; Hoenes, K.; Hessling, M. Photoinactivation of the Coronavirus Surrogate phi6 by Visible Light. Photochem. Photobiol. 2021, 97, 122–125. [Google Scholar] [CrossRef]
- Vatter, P.; Hoenes, K.; Hessling, M. Blue light inactivation of the enveloped RNA virus Phi6. BMC Res. Notes 2021, 14, 187. [Google Scholar] [CrossRef]
- Cartwright, S.F. A cytopathic virus causing a transmissible gastroenteritis in swine. II. Biological and serological studies. J. Comp. Pathol. 1966, 76, 95–106. [Google Scholar] [CrossRef]
- Wallis, C.; Melnick, J.L. Irreversible photosensitization of viruses. Virology 1964, 23, 520–527. [Google Scholar] [CrossRef]
- Wallis, C.; Trulock, S.; Melnick, J.L. Inherent photosensitivity of herpes virus and other enveloped viruses. J. Gen. Virol. 1969, 5, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Nemo, G.J.; Cutchins, E.C. Effect of visible light on canine distemper virus. J. Bacteriol. 1966, 91, 798–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, T.B.; Porter, C.D. Inactivation of murine leukaemia virus by exposure to visible light. Virology 2005, 341, 321–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, J.C.; Stern, H. Photodynamic inactivation of rubella virus. J. Med. Microbiol. 1972, 5, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Zupin, L.; Gratton, R.; Fontana, F.; Clemente, L.; Pascolo, L.; Ruscio, M.; Crovella, S. Blue photobiomodulation LED therapy impacts SARS-CoV-2 by limiting its replication in Vero cells. J. Biophotonics 2021, 14, e202000496. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.; Faustino, M.A.F.; Neves, M.G.P.M.S.; Cunha, A.; Almeida, A. Photodynamic inactivation of mammalian viruses and bacteriophages. Viruses 2012, 4, 1034–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiehe, A.; O’Brien, J.M.; Senge, M.O. Trends and targets in antiviral phototherapy. Photochem. Photobiol. Sci. 2019, 18, 2565–2612. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| (A) Non-Enveloped Viruses | Irradiation Wavelength | Medium during Irradiation | Assumed Riboflavin Concentration [mg/L] | 90% Reduction Dose [J/cm2] |
| lambda phage (dsDNA) | 410 nm | PBS + riboflavin | 18.8 | 4 * [37] |
| PBS | 0 | no reduction after 5 J/cm2 [37] | ||
| adenovirus (dsDNA) | 420 nm | DMEM + FCS | 0.4 | 29 [38] |
| feline calcivirus (ssRNA) | 405 nm | PBS + riboflavin + tyrosine, tryptophan, pyridoxine and folic acid | 0.4 | 82 [39] |
| nutrient rich medium (DMEM + FCS +...) | 0.4 | 88 [39] | ||
| PBS + riboflavin | 0.4 | 329 [39] | ||
| PBS | 0.0 | 719 [39] | ||
| phi C31 (dsDNA) | 405 nm | nutrient rich medium | 0.2 | 113 [40] |
| PBS | 0.0 | 1021 [40] | ||
| viral haemorrhagic septicaemia virus (ssRNA) | 405 nm | L15 + FCS | 0.1 | 114 [41] |
| encephalomyocarditis virus (ssRNA) | 405 nm + blue/white | DMEM + FCS + PBS (ratio unknown) | ? | 178 [42] |
| murine norovirus (ssRNA) | 408 (cw laser) | DMEM + riboflavin | 1.4 | 491 * [43] |
| DMEM | 0.4 | 1976 [43] | ||
| tobacco mosaic virus (ssRNA) | white light | acetate buffer + riboflavin | 0.05 | reduction observed [44] |
| acetate buffer | 0 | no reduction [44] | ||
| Tulane virus (ssRNA) | 405 nm | blueberry surface + riboflavin | reduction [45] | |
| blueberry surface | no reduction after 7.6 J/cm2 [45] | |||
| foot and mouth disease virus (ssRNA) | daylight and artificial light | phosphate saline | 0 | medium reduction [46] |
| poliovirus type 1 (ssRNA) | fluorescent lamp | gelatin saline | 0 | weak reduction [47] |
| human rhinovirus 1b (ssRNA) | 425 nm | DMEM + FCS | 0.4 | no reduction after 45 J/cm2 [48] |
| poliovirus type 3 (ssRNA) | white light of incandescent bulb | medium 199 | 0.01 | no reduction [49] |
| (B) Enveloped Viruses | Irradiation Wavelength | Medium during Irradiation | Assumed Riboflavin Concentration [mg/L] | 90% Reduction Dose [J/cm2] |
| SARS-CoV-2 (ssRNA, coronavirus) | 425 nm | DMEM + FCS | 0.4 | 5 [48] |
| MEM + FCS | 0.1 | 18.8 [48] | ||
| inside vero cells | ? | 6.4 * [48] | ||
| inside epithelial cells | ? | 6.7 * [48] | ||
| SARS-CoV-2 (ssRNA, coronavirus) | 410 nm + blue/white | MEM + FCS (DMEM @ FOI laboratory?) | 0.1 | 6.5 blue (<490 nm) 12.6 total irradiation [50] |
| SARS-CoV-2 (ssRNA, coronavirus) | 405 nm + blue/white | DMEM | 0.4 | 6.6 [51] |
| SARS-CoV-2 (ssRNA, coronavirus) | 405 nm + blue/white | DMEM + FCS + PBS (ratio unknown) | ? | 7.5 (<420 nm) [42] |
| SARS-CoV-1 (ssRNA, coronavirus) | 425 nm | DMEM + FCS | 0.4 | 9.9 [48] |
| feline infectious peritonitis virus (ssRNA, coronavirus) | 405 nm | DMEM + FCS | 0.4 | 14.1 [52] |
| metal wet | 12 * [52] | |||
| metal dry | 20.3 * [52] | |||
| paper wet | 10.8 * [52] | |||
| paper dry | 13 * [52] | |||
| plastic wet | 14.4 * [52] | |||
| plastic dry | 31.8 * [52] | |||
| MERS-CoV (ssRNA, coronavirus) | 425 nm | DMEM + FCS | 0.4 | 18.8 [48] |
| influenza A virus (ssRNA) | 405 nm + blue/white | DMEM + FCS + PBS (ratio unknown) | ? | 23.5 [42] |
| respiratory syncytial virus (ssRNA) | 420 nm | DMEM + FCS | 0.4 | 29 [38] |
| SARS-CoV-2 (ssRNA, coronavirus) | 420 nm | DMEM + FCS | 0.4 | 29 [38] |
| BCoV (ssRNA, coronavirus) | 401 nm | DMEM + FCS | 0.4 | 29 [53] |
| HCoV-229E (ssRNA, coronavirus) | 405 nm (pulsed) | RPMI 1640 | 0.2 | 55 [54] |
| BCoV (ssRNA, coronavirus) | 405 nm | (consumed) RPMI 1640 diluted 1:10 in PBS | 0 | 57.5 [55] |
| steel surface | 96 * [55] | |||
| zika virus (ssRNA) | 445 nm (cw laser) | unknown medium | ? | 64 [56] |
| HCoV-229E (ssRNA, coronavirus) | 405 nm | RPMI 1640 | 0.2 | 89 [54] |
| herpes simplex virus Type 1 (dsDNA) | 445 nm (pulsed) | unknown medium | 112 [57] | |
| phi 6 (dsRNA) | 405 | PBS/SMG | 0 | 400 [58] |
| phi 6 (dsRNA) | 455 | PBS | 0 | 2130 [59] |
| semliki forest virus (ssRNA) | daylight and fluorescent lamp | gelatin saline + riboflavin | 2 | very strong reduction [47] |
| gelatin saline | 0 | strong reduction [47] | ||
| sindbis virus (ssRNA) | daylight and fluorescent lamp | gelatin saline | 0 | strong reduction [47] |
| Murray Valley encephalitis virus (ssRNA) | fluorescent lamp | gelatin saline | 0 | strong reduction [47] |
| transmissible gastroenteritis virus (ssRNA, coronavirus) | daylight | unknown medium | ? | strong reduction [60] |
| influenza B virus (ssRNA) | daylight and artificial light | phosphate saline | 0 | strong reduction [46] |
| vesicular stomatitis virus (ssRNA) | daylight and artificial light | phosphate saline | 0 | strong reduction [46] |
| measles morbillivirus (ssRNA) | white light of fluorescent lamp | salt solutions | 0 | strong reduction [61] |
| herpes simplex virus (dsDNA, JES strain) | white light of fluorescent lamp | MEM | 0.1 | strong reduction [62] |
| riboflavin solution | 0.1 | strong reduction [62] | ||
| salt solutions | 0 | reduction [62] | ||
| distilled water | 0 | no reduction [62] | ||
| canine distemper virus (ssRNA) | artificial visible light | MEM | 0.1 | strong reduction [63] |
| riboflavin solution | 0.1 | strong reduction [63] | ||
| salt solutions | 0 | reduction [63] | ||
| measles morbillivirus (ssRNA) | white light of incandescent bulb | Eagle’s basal medium | 0.1 | strong reduction [49] |
| distilled water | 0 | reduction [49] | ||
| murine leukaemia virus (ssRNA) | 420–430 nm | OptiMEM | 0.1 | reduction [64] |
| rubella virus (ssRNA) | white light of incandescent bulb | PBS | 0 | reduction [65] |
| influenza A virus (ssRNA) | fluorescent lamp | gelatin saline | 0 | reduction [47] |
| parainfluenza virus type 3 (ssRNA) | white light of fluorescent lamp | salt solutions | 0 | reduction [61] |
| SARS-CoV-2 (ssRNA, coronavirus) | 450, 454, 470 nm | inside vero cells | ? | reduction [66] |
| DMEM + FCS | 0.4 | no reduction after 20 J/cm2 [66] | ||
| Newcastle disease virus (ssRNA) | daylight and artificial light | phosphate saline | 0 | weak reduction [46] |
| vaccinia virus (dsDNA) | white light of incandescent bulb | Eagle’s basal medium | 0.1 | weak reduction [49] |
| vaccinia virus (dsDNA) | daylight and artificial light | phosphate saline | 0 | weak reduction [46] |
| influenza A virus (ssRNA) | daylight and artificial light | phosphate saline | 0 | weak reduction [46] |
| fowl plague virus (ssRNA) | daylight and artificial light | phosphate saline | 0 | no reduction [46] |
| rabbit pox virus (dsDNA) | fluorescent lamp | gelatin saline | 0 | no reduction [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hessling, M.; Lau, B.; Vatter, P. Review of Virus Inactivation by Visible Light. Photonics 2022, 9, 113. https://doi.org/10.3390/photonics9020113
Hessling M, Lau B, Vatter P. Review of Virus Inactivation by Visible Light. Photonics. 2022; 9(2):113. https://doi.org/10.3390/photonics9020113
Chicago/Turabian StyleHessling, Martin, Bernhard Lau, and Petra Vatter. 2022. "Review of Virus Inactivation by Visible Light" Photonics 9, no. 2: 113. https://doi.org/10.3390/photonics9020113