Fossil Plant Remains Diagnostics by Laser-Induced Fluorescence and Raman Spectroscopies

 , and
, and 
Abstract
:1. Introduction
2. Experiment
2.1. Instrumentation
2.2. Samples
3. Results and Discussion
3.1. Laser-Induced Fluorescence
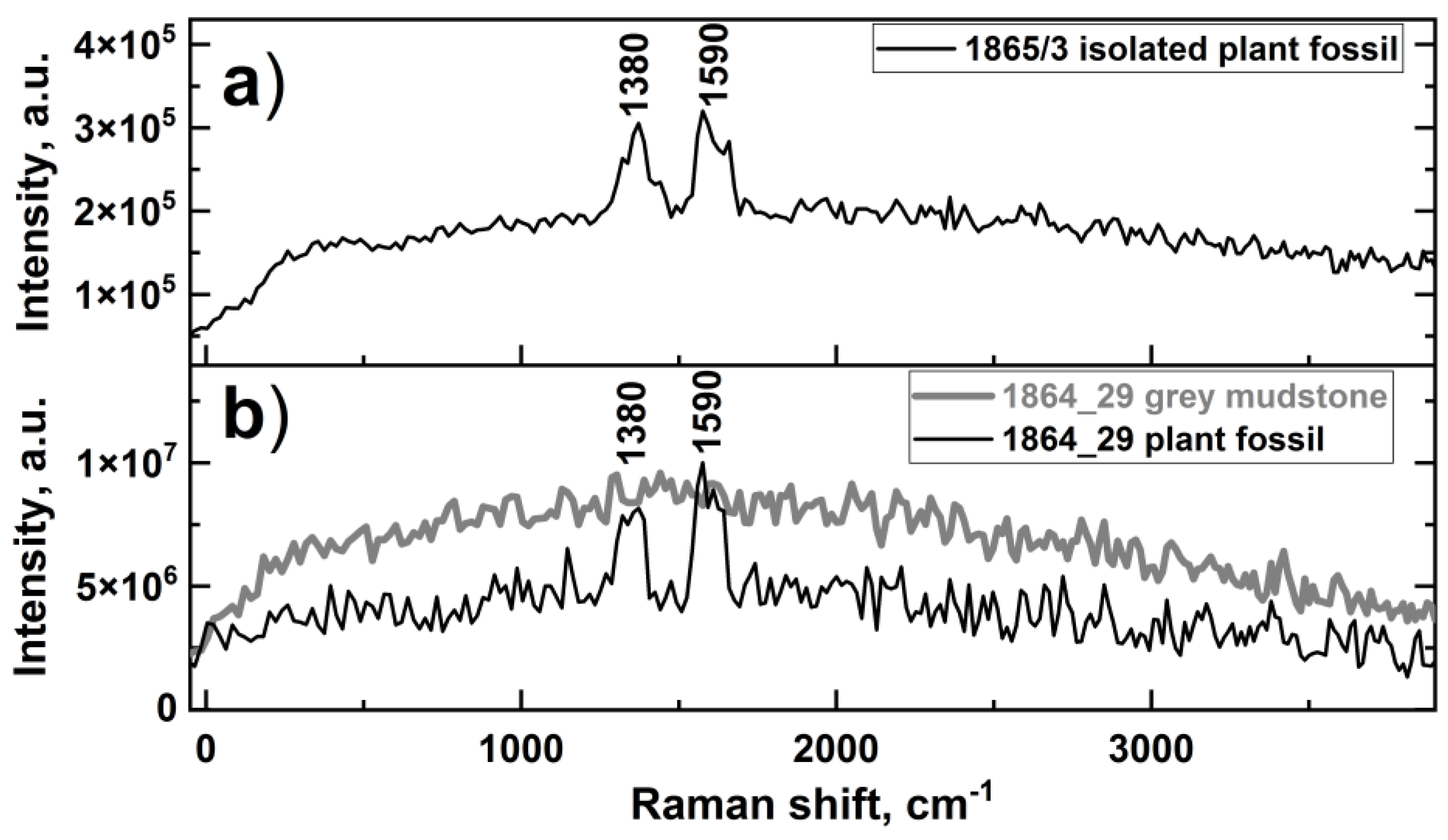
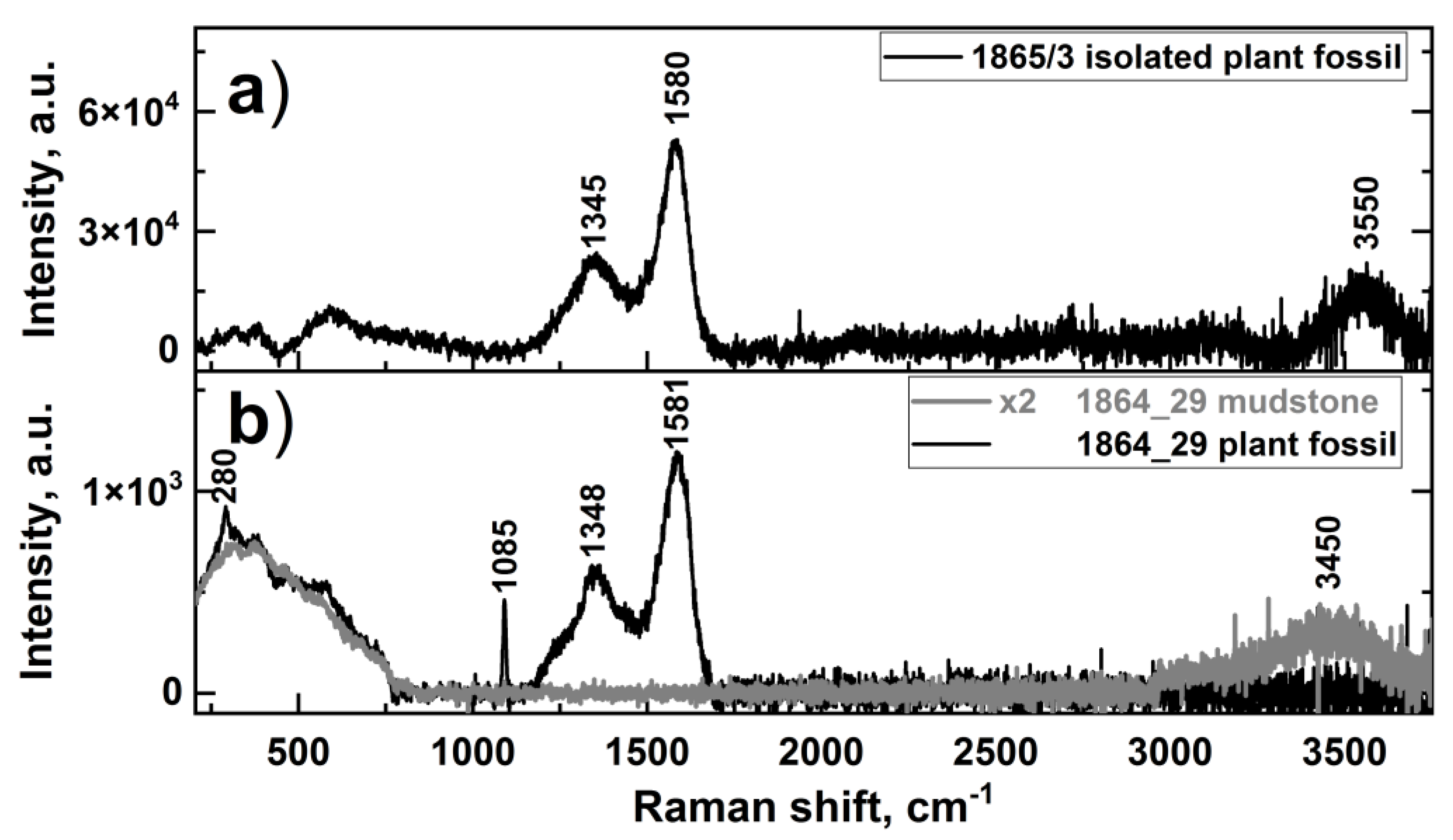
3.2. Raman Spectroscopy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Measures, R.M. Laser Remote Sensing: Fundamentals and Applications; John Wiley & Sons, Ltd.: New York, NY, USA, 1984; ISBN 0894646192. [Google Scholar]
- Bunkin, A.; Voliak, K. Laser Remote Sensing of the Ocean: Methods and Applications; Wiley: New York, NY, USA; Chichester, UK; Weinheim, Germany; Brisbane, Australia; Singapore; Toronto, ON, Canada, 2001; ISBN 0471389277. [Google Scholar]
- Lin, H.; Zhang, Y.; Mei, L. Fluorescence Scheimpflug LiDAR developed for the three-dimension profiling of plants. Opt. Express 2020, 28, 9269–9279. [Google Scholar] [CrossRef] [PubMed]
- Grishin, M.Y.; Lednev, V.N.; Pershin, S.M.; Kapralov, P.O. Ultracompact Fluorescence Lidar Based on a Diode Laser (405 nm, 150 mW) for Remote Sensing of Waterbodies and the Underlying Surface from Unmanned Aerial Vehicles. Dokl. Phys. 2021, 66, 153–155. [Google Scholar] [CrossRef]
- Lednev, V.N.; Bunkin, A.F.; Pershin, S.M.; Grishin, M.Y.; Artemova, D.G.; Zavozin, V.A.; Sdvizhenskii, P.A.; Nunes, R.A. Remote Laser Induced Fluorescence of Soils and Rocks. Photonics 2021, 8, 411. [Google Scholar] [CrossRef]
- Mei, L.; Guan, Z.G.; Zhou, H.J.; Lv, J.; Zhu, Z.R.; Cheng, J.A.; Chen, F.J.; Löfstedt, C.; Svanberg, S.; Somesfalean, G. Agricultural pest monitoring using fluorescence lidar techniques. Appl. Phys. B 2012, 106, 733–740. [Google Scholar] [CrossRef]
- Hackley, P.C.; Jubb, A.M.; McAleer, R.J.; Valentine, B.J.; Birdwell, J.E. A review of spatially resolved techniques and applications of organic petrography in shale petroleum systems. Int. J. Coal Geol. 2021, 241, 103745. [Google Scholar] [CrossRef]
- Saito, Y.; Ichihara, K.; Morishita, K.; Uchiyama, K.; Kobayashi, F.; Tomida, T. Remote Detection of the Fluorescence Spectrum of Natural Pollens Floating in the Atmosphere Using a Laser-Induced-Fluorescence Spectrum (LIFS) Lidar, Y.-I. Remote Detection of the Fluorescence Spectrum of Natural Pollens Floating in the Atmosphere Using a Laser-Induced-Fluorescence Spectrum (LIFS) Lidar. Remote Sens. 2018, 10, 1533. [Google Scholar]
- Zheng, D.; Peng, T.; Zhu, S.; Lian, M.; Li, Y.; Wei, F.; Xiong, J.; Svanberg, S.; Zhao, Q.; Hu, J.; et al. Optical characterization of Chinese hybrid rice using laser-induced fluorescence techniques—laboratory and remote-sensing measurements. Appl. Opt. 2018, 57, 3481–3487. [Google Scholar] [CrossRef]
- Chase, A.F.; Chase, D.Z.; Weishampel, J.F.; Drake, J.B.; Shrestha, R.L.; Slatton, K.C.; Awe, J.J.; Carter, W.E. Airborne LiDAR, archaeology, and the ancient Maya landscape at Caracol, Belize. J. Archaeol. Sci. 2011, 38, 387–398. [Google Scholar] [CrossRef]
- Chand, D.; Anderson, T.L.; Wood, R.; Charlson, R.J.; Hu, Y.; Liu, Z.; Vaughan, M. Quantifying above-cloud aerosol using spaceborne lidar for improved understanding of cloudy-sky direct climate forcing. J. Geophys. Res. Atmos. 2008, 113, D13206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Meyer, K.; Yu, H.; Platnick, S.; Colarco, P.; Liu, Z.; Oreopoulos, L. Shortwave direct radiative effects of above-cloud aerosols over global oceans derived from 8~years of CALIOP and MODIS observations. Atmos. Chem. Phys. 2016, 16, 2877–2900. [Google Scholar] [CrossRef] [Green Version]
- Comerón, A.; Muñoz-Porcar, C.; Rocadenbosch, F.; Rodríguez-Gómez, A.; Sicard, M. Current research in lidar technology used for the remote sensing of atmospheric aerosols. Sensors 2017, 17, 1450. [Google Scholar] [CrossRef] [PubMed]
- Ceolato, R.; Berg, M.J. Aerosol light extinction and backscattering: A review with a lidar perspective. J. Quant. Spectrosc. Radiat. Transf. 2021, 262, 107492. [Google Scholar] [CrossRef]
- Manrique, J.A.; Lopez-Reyes, G.; Cousin, A.; Rull, F.; Maurice, S.; Wiens, R.C.; Madsen, M.B.; Madariaga, J.M.; Gasnault, O.; Aramendia, J.; et al. SuperCam Calibration Targets: Design and Development. Space Sci. Rev. 2020, 216, 138. [Google Scholar] [CrossRef]
- Maurice, S.; Wiens, R.C.; Bernardi, P.; Caïs, P.; Robinson, S.; Nelson, T.; Gasnault, O.; Reess, J.-M.; Deleuze, M.; Rull, F.; et al. The SuperCam Instrument Suite on the Mars 2020 Rover: Science Objectives and Mast-Unit Description. Space Sci. Rev. 2021, 217, 47. [Google Scholar] [CrossRef]
- Pershin, S.M.; Pungin, V.G. The Search for Water on Mars by the Differential Absorption Method Using the 970 nm Band of the Second Overtone of OH Vibrations. Phys. Vib. 2001, 9, 84–89. [Google Scholar]
- Bhartia, R.; Beegle, L.W.; DeFlores, L.; Abbey, W.; Razzell Hollis, J.; Uckert, K.; Monacelli, B.; Edgett, K.S.; Kennedy, M.R.; Sylvia, M.; et al. Perseverance’s Scanning Habitable Environments with Raman and Luminescence for Organics and Chemicals (SHERLOC) Investigation. Space Sci. Rev. 2021, 217, 58. [Google Scholar] [CrossRef]
- Beegle, L.; Bhartia, R.; White, M.; DeFlores, L.; Abbey, W.; Wu, Y.-H.; Cameron, B.; Moore, J.; Fries, M.; Burton, A.; et al. SHERLOC: Scanning habitable environments with Raman & luminescence for organics & chemicals. In Proceedings of the 2015 IEEE Aerospace Conference, Montana, MT, USA, 7–14 March 2015; pp. 1–11. [Google Scholar]
- Glauser, A.L.; Harper, C.J.; Taylor, T.N.; Taylor, E.L.; Marshall, C.P.; Marshall, A.O. Reexamination of cell contents in Pennsylvanian spores and pollen grains using Raman spectroscopy. Rev. Palaeobot. Palynol. 2014, 210, 62–68. [Google Scholar] [CrossRef]
- Marshall, A.O.; Wehrbein, R.L.; Lieberman, B.S.; Marshall, C.P. Raman Spectroscopic Investigations of Burgess Shale–Type Preservation: A New Way Forward. Palaios 2012, 27, 288–292. [Google Scholar] [CrossRef]
- Shkolyar, S.; Eshelman, E.J.; Farmer, J.D.; Hamilton, D.; Daly, M.G.; Youngbull, C. Detecting Kerogen as a Biosignature Using Colocated UV Time-Gated Raman and Fluorescence Spectroscopy. Astrobiology 2018, 18, 431–453. [Google Scholar] [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B.; Agresti, D.G.; Czaja, A.D.; Wdowiak, T.J. Raman Imagery: A New Approach to Assess the Geochemical Maturity and Biogenicity of Permineralized Precambrian Fossils. Astrobiology 2005, 5, 333–371. [Google Scholar] [CrossRef]
- Marshall, C.P.; Edwards, H.G.M.; Jehlicka, J. Understanding the Application of Raman Spectroscopy to the Detection of Traces of Life. Astrobiology 2010, 10, 229–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonneville, S.; Delpomdor, F.; Préat, A.; Chevalier, C.; Araki, T.; Kazemian, M.; Steele, A.; Schreiber, A.; Wirth, R.; Benning, L.G. Molecular identification of fungi microfossils in a Neoproterozoic shale rock. Sci. Adv. 2022, 6, eaax7599. [Google Scholar] [CrossRef]
- Hackley, P.C.; Jubb, A.M.; Burruss, R.C.; Beaven, A.E. Fluorescence spectroscopy of ancient sedimentary organic matter via confocal laser scanning microscopy (CLSM). Int. J. Coal Geol. 2020, 223, 103445. [Google Scholar] [CrossRef]
- Jiang, Y.L.; Liu, Y.-S. A simple and convenient determination of perylene preserved in the Late Neogene wood from northeastern Tennessee using fluorescence spectroscopy. Org. Geochem. 2008, 39, 1462–1465. [Google Scholar] [CrossRef]
- Da Conceição, D.M.; Da Silva, J.H.; Cisneros, J.C.; Iannuzzi, R.; Viana, B.C.; Saraiva, G.D.; Sousa, J.P.; Freire, P.T.C. Spectroscopic studies on Permian plant fossils in the Pedra de Fogo Formation from the Parnaíba Basin, Brazil. J. King Saud Univ. Sci. 2018, 30, 483–488. [Google Scholar] [CrossRef]
- Tahoun, M.; Gee, C.T.; McCoy, V.E.; Sander, P.M.; Müller, C.E. Chemistry of porphyrins in fossil plants and animals. RSC Adv. 2021, 11, 7552–7563. [Google Scholar] [CrossRef] [PubMed]
- McElwain, J.C.; Chaloner, W.G. The fossil cuticle as a skeletal record of environmental change. Palaios 1996, 11, 376–388. [Google Scholar] [CrossRef]
- Mösle, B.; Collinson, M.E.; Finch, P.; Stankiewicz, B.A.; Scott, A.C.; Wilson, R. Factors influencing the preservation of plant cuticles: A comparison of morphology and chemical composition of modern and fossil examples. Org. Geochem. 1998, 29, 1369–1380. [Google Scholar] [CrossRef]
- Taylor, A.T.; Lai, E.P.C. Current State of Laser-Induced Fluorescence Spectroscopy for Designing Biochemical Sensors. Chemosensors 2021, 9, 275. [Google Scholar] [CrossRef]
- Naumov, A.V. Low-temperature spectroscopy of organic molecules in solid matrices: From the Shpol’skii effect to laser luminescent spectromicroscopy for all effectively emitting single molecules. Phys. Uspekhi 2013, 56, 605. [Google Scholar] [CrossRef]
- Johnson, M.E.; Landers, J.P. Fundamentals and practice for ultrasensitive laser-induced fluorescence detection in microanalytical systems. Electrophoresis 2004, 25, 3513–3527. [Google Scholar] [CrossRef] [PubMed]
- Bunkin, A.F.; Kolesnikov, M.P.; Pershin, S.M. Laser Fluorescence of Soil: Online Remote Sensing of the Earth’s Surface. Phys. Vib. 1998, 6, 249–255. [Google Scholar]
- Agati, G.; Biricolti, S.; Guidi, L.; Ferrini, F.; Fini, A.; Tattini, M. The biosynthesis of flavonoids is enhanced similarly by UV radiation and root zone salinity in L. vulgare leaves. J. Plant Physiol. 2011, 168, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Koşar, M.; Dorman, H.J.D.; Hiltunen, R. Effect of an acid treatment on the phytochemical and antioxidant characteristics of extracts from selected Lamiaceae species. Food Chem. 2005, 91, 525–533. [Google Scholar] [CrossRef]
- Florovskaya, V.N.; Zezin, R.B.; Ovchinnikova, L.I.; Pikovsky, Y.I.; Teplitskaya, T.A. Diagnostics of Organic Substances in the Rocks and Minerals of Magmatic and Hydrothermal Origin; Moscow, Russia, 1968. (In Russian) [Google Scholar]
- Kolesnikov, M.P.; Voronova, N.I.; Egorov, I.A. Metalloporphyrins and Perylene in the Products of Volcano Eruption. Dokl. Akad. Nauk SSSR 1979, 247, 749–752. [Google Scholar]
- Hodgson, G.W.; Hitchon, B.; Taguchi, K.; Baker, B.L.; Peake, E. Geochemistry of porphyrins, chlorins and polycyclic aromatics in soils, sediments and sedimentary rocks. Geochim. Cosmochim. Acta 1968, 32, 737–772. [Google Scholar] [CrossRef]
- Kolesnikov, M.P.; Egorov, I.A. Chlorophyll derivatives in modern soils in connection with the problem of the chemical evolution and origin of life on earth. Dokl. Biochem. Akad Nauk SSSR 1977, 235, 228–231. [Google Scholar]
- O’Malley, C.E.; Ausich, W.I.; Chin, Y.-P. Deep echinoderm phylogeny preserved in organic molecules from Paleozoic fossils. Geology 2016, 44, 379–382. [Google Scholar] [CrossRef] [Green Version]
- Blumer, M.; Omenn, G.S. Fossil porphyrins: Uncomplexed chlorins in a Triassic sediment. Geochim. Cosmochim. Acta 1961, 25, 81–90. [Google Scholar] [CrossRef]
- Kolesnikov, M.P.; Egorov, I.A. Metalloporphyrins in precambrian deposits as probable evidence of ancient photosynthesis [Spirulina platensis, Algae]. Dokl. Biochem. Akad. Nauk 1979, 244, 470–473. [Google Scholar]
- Svec, W.A. The isolation, preparation, characterization, and estimation of the chlorophylls and the bacteriochlorophylls. Porphyrins 1978, 5, 341–399. [Google Scholar]
- Koifman, O.I.; Askarov, K.A.; Berezin, B.D. Enikolopyan NS Natural Sources of Porphyrins. Methods for Isolation and Modification of Natural Porphyrins. Porphyr. Struct. Prop. Synth. 1985, 1, 75–204. [Google Scholar]
- Ellsworth, R.K.; Carney, C.F. Preparation of Chlorin-E6 from Microgram Quantities of Pheophorbide-A and Its Absorption-Spectrum in Diethyl-Ether. Photosynthetica 1975, 9, 333–336. [Google Scholar]
- Dolphin, D. The Porphyrins V1: Structure and Synthesis, Part A; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Watts, C.D.; Simoneit, B.R.; Maxwell, J.R.; Ragot, J.P. The quincyite pigments. A novel series of fossil “dyes” from an Eocene sediment. In Proceedings of the Advances in Organic Geochemistry 1975: Actas del 7th Congreso Internacional de Geoquimica Organica, Madrid, Spain, 16–19 September 1975; Volume 19, pp. 223–235. [Google Scholar]
- Shuvalov, V.A.; Krasnovskii, A.A. The luminescence of zinc-porphyrins in microorganisms and plants: Phosphorescence and delayed fluorescence. Mol. Biol. 1971, 5, 557–566. [Google Scholar] [PubMed]
- Schopf, W.J.; Kudryavtsev, B.A.; Agresti, G.D.; Wdowiak, J.T.; Czaja, D.A. Laser--Raman imagery of Earth’s earliest fossils. Nature 2002, 416, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Gan, T.; Luo, T.; Pang, K.; Zhou, C.; Zhou, G.; Wan, B.; Li, G.; Yi, Q.; Czaja, A.D.; Xiao, S. Cryptic terrestrial fungus-like fossils of the early Ediacaran Period. Nat. Commun. 2021, 12, 641. [Google Scholar] [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B. Three-dimensional Raman imagery of precambrian microscopic organisms. Geobiology 2005, 3, 1–12. [Google Scholar] [CrossRef]
- Pasteris, J.D.; Wopenka, B. Necessary, but Not Sufficient: Raman Identification of Disordered Carbon as a Signature of Ancient Life. Astrobiology 2003, 3, 727–738. [Google Scholar] [CrossRef]
- Tuinstra, F.; Koenig, J.L. Raman Spectrum of Graphite. J. Chem. Phys. 1970, 53, 1126–1130. [Google Scholar] [CrossRef]
- Bower, D.M.; Steele, A.; Fries, M.D.; Green, O.R.; Lindsay, J.F. Raman imaging spectroscopy of a putative microfossil from the∼ 3.46 Ga Apex chert: Insights from quartz grain orientation. Astrobiology 2016, 16, 169–180. [Google Scholar] [CrossRef]
- Rozanov, A.Y.; Hoover, R.B.; Krasavin, E.A.; Samyilina, O.S.; Ryumin, A.K.; Kapralov, M.I.; Saprykin, E.A.; Afanasyieva, A.N. The Orgueil Meteorite (Atlas of Microfossils); JINR: Moscow, Russia, 2020; ISBN 978-5-903825-42-4. [Google Scholar]








{kind=link}
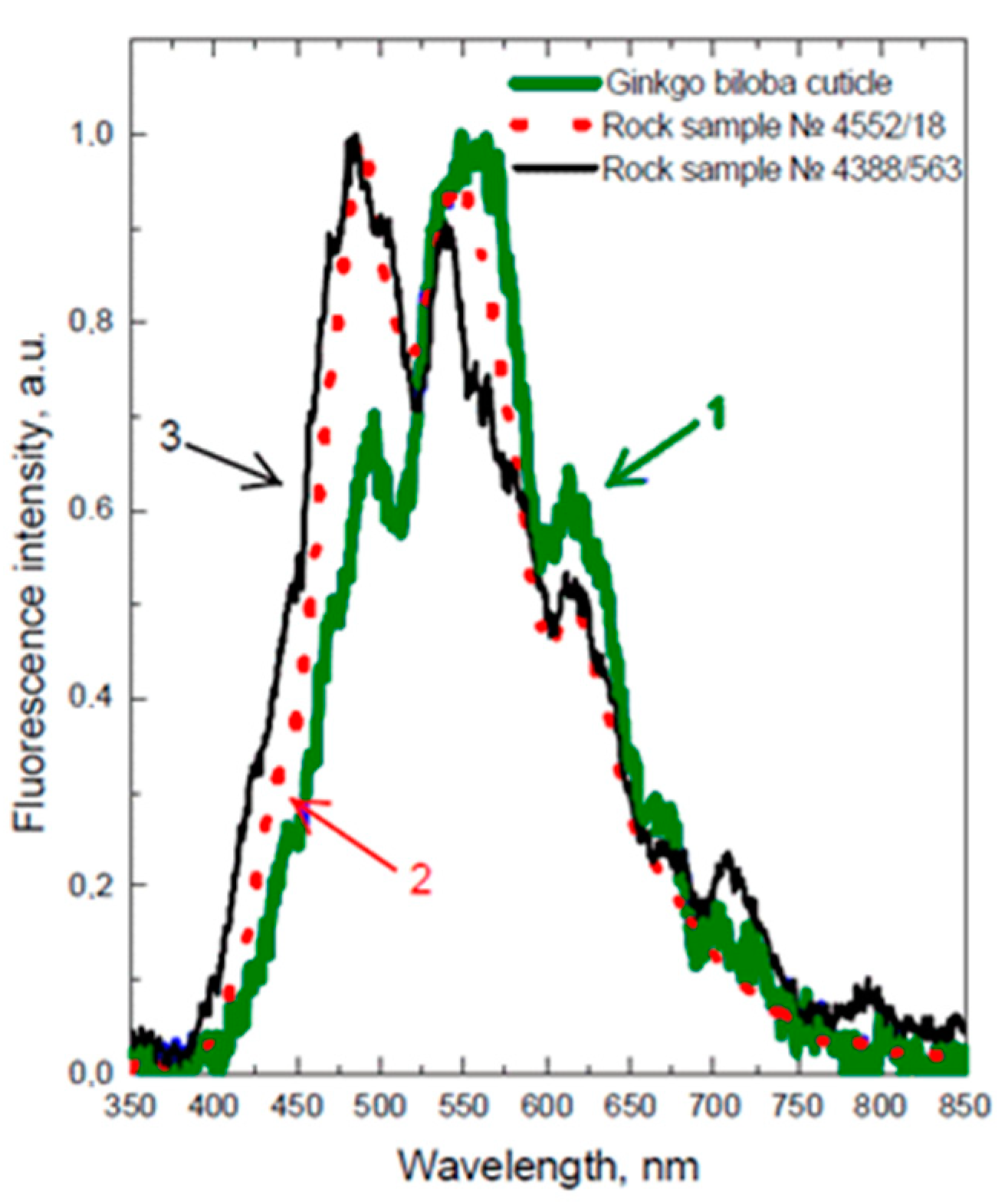{kind=link}
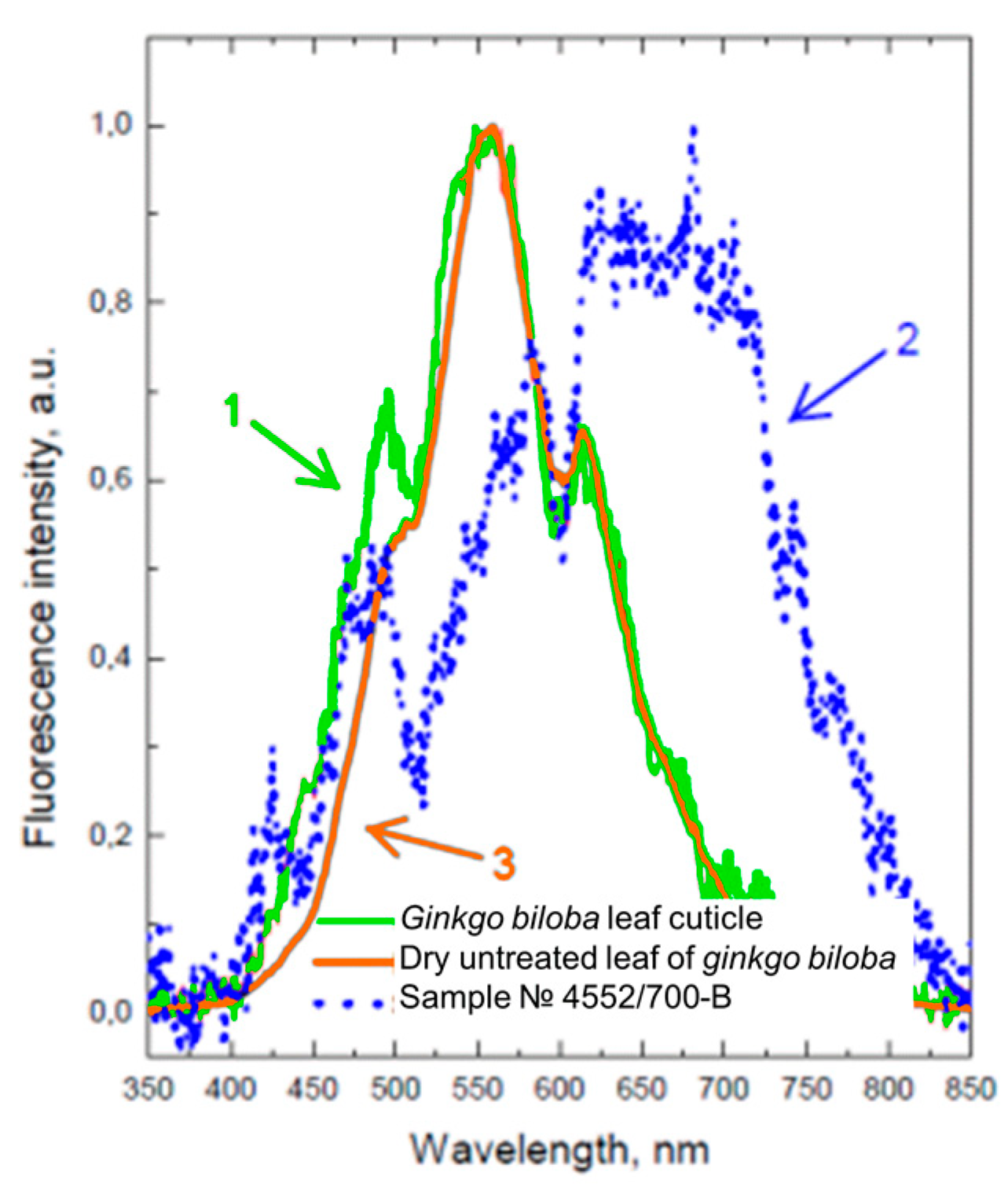{kind=link}
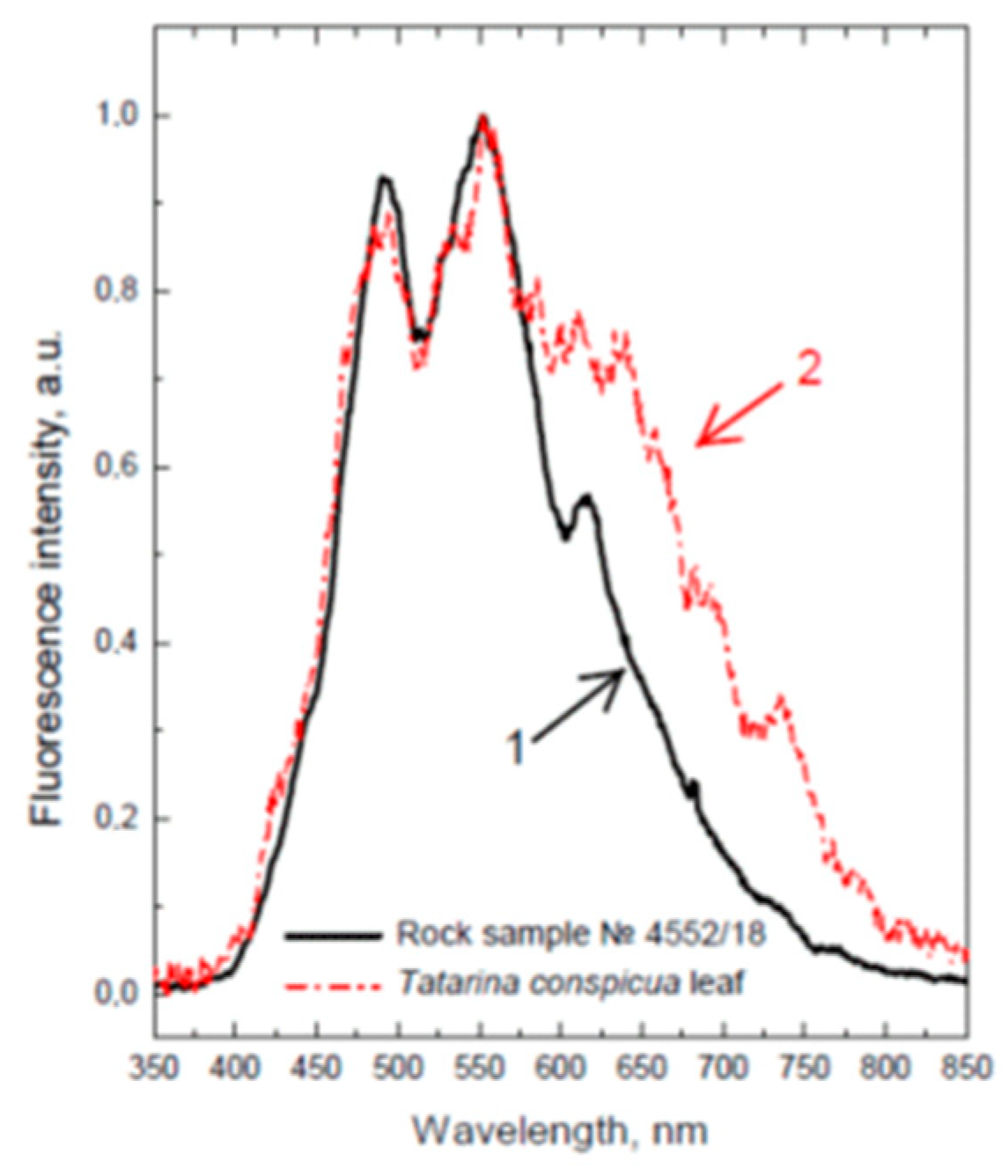{kind=link}
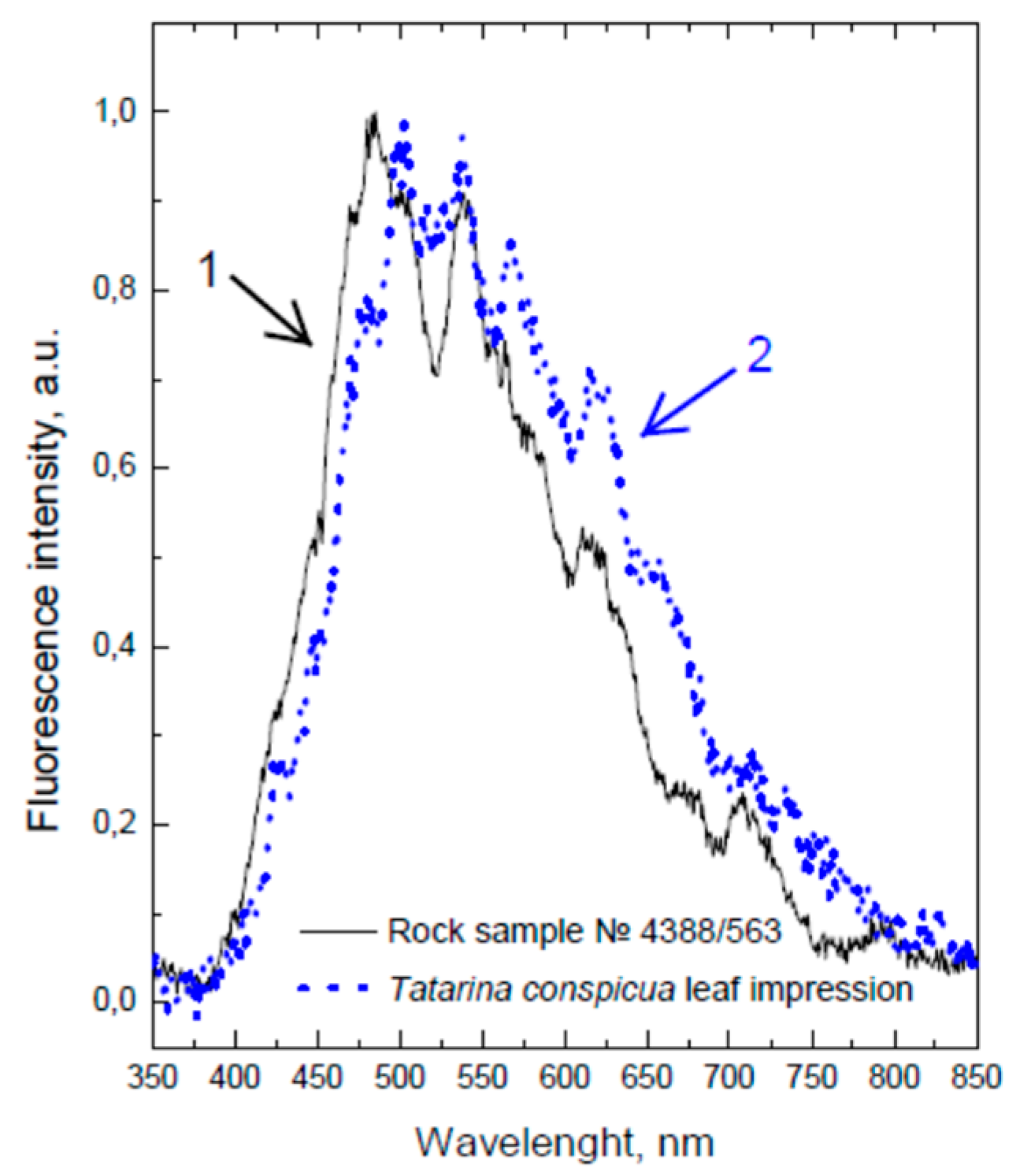{kind=link}
{kind=link}
{kind=link}
| Sample | Photo | Description | Origin | Age, Myr |
|---|---|---|---|---|
| 1865/3 |  | isolated chemically untreated fossilized material of Schuguria ornata | Kursk Region, Mikhailovsky quarry, Kursk Magnetic Anomaly | 383 |
| 4552/18 |  | well-preserved fossilized leaf of Tatarina conspicua in rock (siltstone) | Vologda Region, the right bank of the Malaya Severnaya Dvina River at the Aristovo pier | 257 |
| 4388/563 |  | impression of the gymnospermous plant Tatarina conspicua (family Peltaspermaceae, order Peltaspermales) leaf in rock (siltstone) with fossilized material residues | Kirov Region, the right bank of the Vyatka River 13 km below Kotelnich city | 260 |
| 4552/700 |  | isolated chemically untreated fossilized leaf of the gymnospermous plant Phylladoderma (Aequistomia) annulata (family Cardiolepidaceae, order Peltaspermales) | Vologda Region, the right bank of the Malaya Severnaya Dvina River at the Aristovo pier | 257 |
| 4552/700-B | pure cuticle of the same leaf (Phylladoderma (Aequistomia) annulata) | Vologda Region, the right bank of the Malaya Severnaya Dvina River at the Aristovo pier | 257 | |
| 1860/218A |  | impressions of leaves of cordaites (primitive gymnosperms) Cordaites sp. with scarcely preserved coal matter in grey siltstone | Yaman-Us well, South Mongolia | 251 |
| 1864/29 |  | impression of conifer twig Taxodiella bardaeana with scarcely preseved organic matter in grey mudstone | Orenburg Region, Dubensky quarry | 270 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bunkin, A.F.; Pershin, S.M.; Artemova, D.G.; Gudkov, S.V.; Gomankov, A.V.; Sdvizhenskii, P.A.; Grishin, M.Y.; Lednev, V.N. Fossil Plant Remains Diagnostics by Laser-Induced Fluorescence and Raman Spectroscopies. Photonics 2023, 10, 15. https://doi.org/10.3390/photonics10010015
Bunkin AF, Pershin SM, Artemova DG, Gudkov SV, Gomankov AV, Sdvizhenskii PA, Grishin MY, Lednev VN. Fossil Plant Remains Diagnostics by Laser-Induced Fluorescence and Raman Spectroscopies. Photonics. 2023; 10(1):15. https://doi.org/10.3390/photonics10010015
Chicago/Turabian StyleBunkin, Alexey F., Sergey M. Pershin, Diana G. Artemova, Sergey V. Gudkov, Alexey V. Gomankov, Pavel A. Sdvizhenskii, Mikhail Ya. Grishin, and Vasily N. Lednev. 2023. "Fossil Plant Remains Diagnostics by Laser-Induced Fluorescence and Raman Spectroscopies" Photonics 10, no. 1: 15. https://doi.org/10.3390/photonics10010015