
Dipteran Prey Vulnerability in Intraguild Predation (IGP) System Involving Heteropteran Predators: Density and Habitat Effects


Abstract
:1. Introduction
2. Material and Methods
2.1. Collection and Maintenance of Water Bugs
2.2. Collection and Maintenance of Mosquito Larvae
2.3. Experimental Design
- (1)
- Simple or open habitat: without pebbles and vegetation.
- (2)
- Complex habitat: with pebbles and vegetation (4 Ipomoea sticks at 2 corners of aquarium and some Pistia were given, which were in a floating state).
2.4. Data Analysis
2.4.1. Multiplicative Risk Model
2.4.2. Clearance Rates
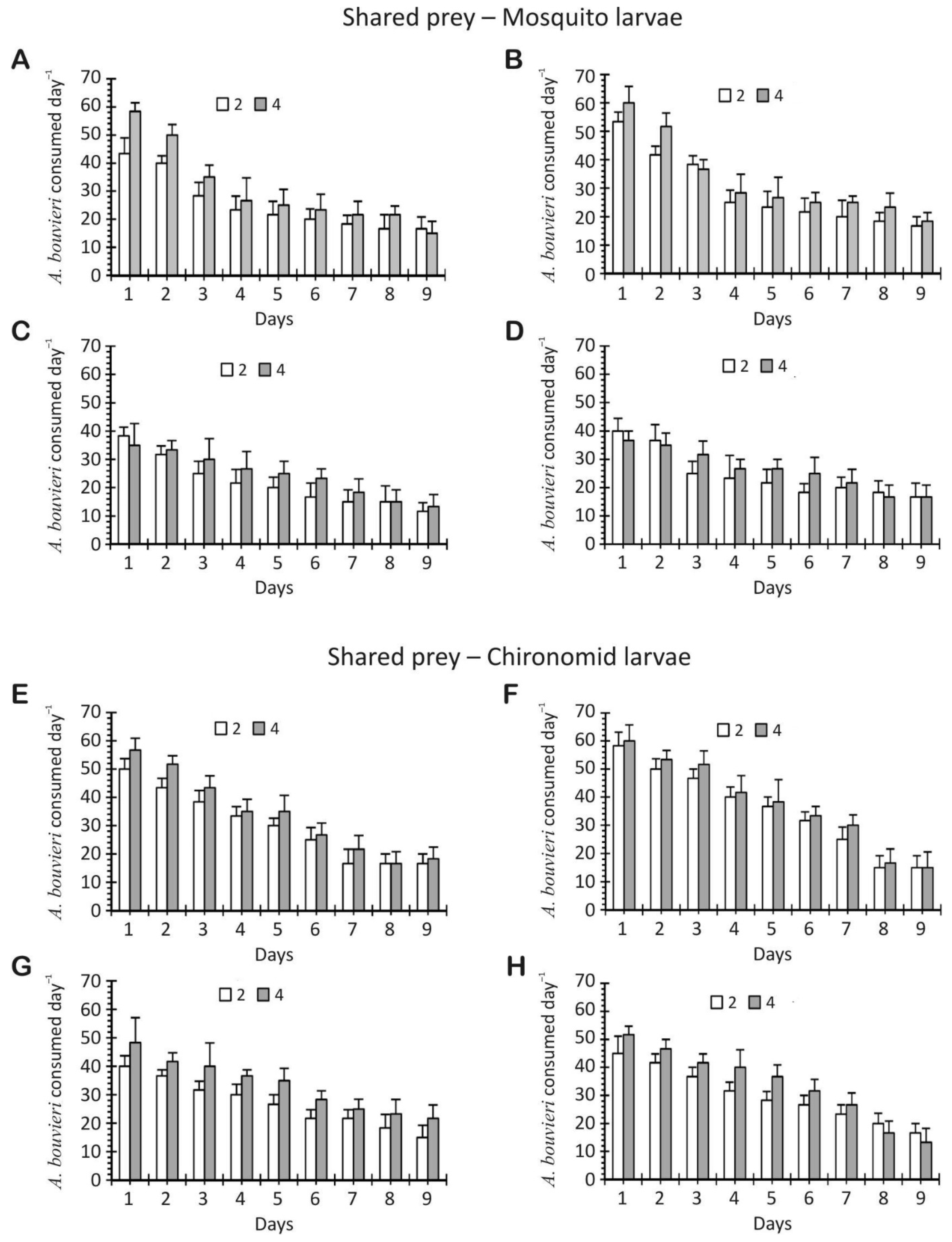
3. Results
- (a)
- IG prey mortality was IG-predator dependent—high mortality at high IG predator density;
- (b)
- IG prey mortality was shared-prey-density dependent—low mortality at high shared-prey density;
- (c)
- IG prey mortality was habitat-condition dependent—high mortality in complex habitat condition.
3.1. Multiplicative Risk Model
3.2. Clearance Rates
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wissinger, S.A. Niche overlap and the potential for competition and intraguild predation between size-structured populations. Ecology 1992, 73, 1431–1444. [Google Scholar] [CrossRef]
- Wissinger, S.A.; McGrady, J. Intraguild predation and competition between larval dragonflies: Direct and indirect effects on shared prey. Ecology 1993, 74, 207–218. [Google Scholar] [CrossRef]
- Crumrine, P.W. Size structure and substitutability in an odonate intraguild predation system. Oecologia 2005, 145, 132–139. [Google Scholar] [CrossRef]
- Denno, R.F.; Finke, D.L.; Langellotto, G.A. Direct and indirect effects of vegetation structure and habitat complexity on predator-prey and predator-predator interactions. In Ecology of Predator-Prey Interactions; Barbosa, P., Castellanos, I., Eds.; Oxford University Press: London, UK, 2005; pp. 211–239. [Google Scholar]
- Saha, N.; Aditya, G.; Saha, G.K. Habitat complexity reduces vulnerability of preys: An experimental analysis using aquatic insect predators and dipteran immature. J. Asia-Pac. Entomol. 2009, 12, 233–239. [Google Scholar] [CrossRef]
- Flynn, K.E.; Moon, D.C. Effects of habitat complexity, prey type, and abundance on in traguild predation between larval odonates. Hydrobiologia 2011, 675, 97–104. [Google Scholar] [CrossRef]
- Gagnon, A.-È.; Heimpel, G.E.; Brodeur, J. The ubiquity of intraguild predation among predatory arthropods. PLoS ONE 2011, 6, e28061. [Google Scholar] [CrossRef] [PubMed]
- Jana, D.; Bairagi, N. Habitat complexity, dispersal and metapopulations: Macroscopic study of a predator-prey system. Ecol. Complex. 2014, 17, 131–139. [Google Scholar] [CrossRef]
- Crowder, L.B.; Cooper, W.E. Habitat structure complexity and the interaction be tween Bluegills and their prey. Ecology 1982, 63, 1802–1813. [Google Scholar] [CrossRef]
- Summerson, H.C.; Peterson, C.H. Role of predation in organizing benthic communities of a temperate-zone seagrass bed. Mar. Ecol. Prog. Ser. 1984, 15, 63–77. [Google Scholar] [CrossRef]
- Hughes, A.R.; Gabrowsky, J.H. Habitat context influences predator interference interactions and the strength of resource partitioning. Oecologia 2006, 146, 256–264. [Google Scholar] [CrossRef]
- Finke, D.L.; Denno, R.F. Intraguild predation diminished in complex habitats: Implications for top-down suppression of prey populations. Ecology 2002, 83, 643–652. [Google Scholar] [CrossRef]
- Siddon, C.E.; Witman, J.E. Behavioral indirect interactions: Multiple predator effects and prey switching in rocky subtidal. Ecology 2004, 85, 2938–2945. [Google Scholar] [CrossRef]
- Griffen, B.D.; Byers, J.E. Intraguild predation reduces redundancy of predator species in multiple predator assemblage. J. Anim. Ecol. 2006, 75, 959–966. [Google Scholar] [CrossRef]
- Buxton, M.; Cuthbert, R.N.; Dalu, T.; Nyamukondiwaa, C.; Wassermana, R.J. Predator density modifies mosquito regulation in increasingly complex environment. Pest. Manag. Sci. 2020, 76, 2079–2086. [Google Scholar] [CrossRef] [PubMed]
- De Roy, E.M.; Scott, R.; Hussey, N.E.; Maclsaac, H.J. Density dependence mediates the ecological impact of an invasive fish. Divers. Distrib. 2020, 26, 867–880. [Google Scholar] [CrossRef]
- Vance-Chalcraft, H.D.; Soluk, D.A. Multiple predator effects result in risk reduction for prey across multiple prey densities. Oecologia 2005, 144, 472–480. [Google Scholar] [CrossRef]
- Brahma, S.; Sharma, D.; Kundu, M.; Saha, N.; Aditya, G.; Saha, G.K. Mosquito prey vulnerability in intraguild predation between Ranatra filiformis and Anisops bouvieri: Implications in biological control. Proc. Zool. Soc. 2015, 68, 36–44. [Google Scholar] [CrossRef]
- Brahma, S.; Sharma, D.; Kundu, M.; Saha, N.; Saha, G.K.; Aditya, G. Intraguild predation in Heteroptera: Effects of density and predator identity on dipteran prey. Neotrop. Entomol. 2015, 44, 374–384. [Google Scholar] [CrossRef]
- Fischer, S.; Marinone, M.C.; Fontanarrosa, M.S.; Nieves, M.; Schweigmann, N. Urban rain pools: Seasonal dynamics and entomofauna in a park of Buenos Aires. Hydrobiologia 2000, 444, 45–63. [Google Scholar] [CrossRef]
- Bambaradeniya, C.N.B.; Edirisinghe, J.P.; Silva, D.N.; Gunatilleke, C.V.S.; Ranawana, K.B.; Wijekoon, S. Biodiversity associated with rice agro-ecosystem in Sri Lanka. Biodivers. Conserv. 2004, 13, 1715–1753. [Google Scholar] [CrossRef]
- Das, P.K.; Sivagnaname, N.; Amalraj, D.D. Population interactions between Culex vishnui mosquitoes and their natural enemies in Pondicherry, India. J. Vector. Ecol. 2006, 31, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Aditya, G.; Saha, N.; Saha, G.K. An assessment of macroinvertebrate as semblages in mosquito larval habitats –space and diversity relationship. Environ. Monit. Assess. 2010, 168, 597–611. [Google Scholar] [CrossRef]
- Urcola, J.I.; Fischer, S. Seasonal and environmental variables related to the abundance of immature mosquitoes in rain-pools of a peri-urban park of Buenos Aires (Argentina). J. Med. Entomol. 2019, 56, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Kundu, M.; Sharma, D.; Brahma, S.; Pramanik, S.; Saha, G.K.; Aditya, G. Insect predators of mosquitoes of rice fields: Portrayal of indirect interactions with alternative prey. J. Entomol. Zool. Stud. 2014, 2, 97–103. [Google Scholar]
- Polis, G.A.; Myers, C.A.; Holt, R.D. The ecology and evolution of intraguild predation: Potential competitors that eat each other. Annu. Rev. Ecol. Evol. Syst. 1989, 20, 297–330. [Google Scholar] [CrossRef]
- Polis, G.A.; Holt, R.D. Intraguild predation: The dynamics of complex trophic interacttion. Trends Ecol. Evol. 1992, 7, 151–154. [Google Scholar] [CrossRef]
- Holt, R.D.; Polis, G.A. A theoretical framework for intraguild predation. Am. Nat. 1997, 149, 745–764. [Google Scholar] [CrossRef]
- Arim, M.; Marquet, P.A. Intraguild predation: A widespread interaction related to species biology. Ecol. Lett. 2004, 7, 557–564. [Google Scholar] [CrossRef]
- Brahma, S.; Aditya, G.; Sharma, D.; Saha, N.; Kundu, M.; Saha, G.K. Influence of density on intraguild predation of aquatic hemipteran (Heteroptera): Implications in biological control of mosquito. J. Entomol. Acarol. Res. 2014, 46, 1977. [Google Scholar] [CrossRef]
- Diehl, S. Foraging efficiency of three freshwater fishes: Effects of structural complexity and light. Oikos 1988, 53, 207–214. [Google Scholar] [CrossRef]
- Hurst, T.P.; Kay, B.H.; Brown, M.D.; Ryan, P.A. Laboratory evaluation of the effect of alternative prey and vegetation on predation of Culex annulirostris immatures by Australian native fish species. J. Am. Mosq. Control Assoc. 2006, 22, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Kopp, K.; Wachlevski, M.; Eterovick, P.C. Environmental complexity reduces tadpole predation by water bugs. Can. J. Zool. 2006, 84, 136–140. [Google Scholar] [CrossRef]
- Babbitt, K.J.; Jordan, F. Predation on Bufo terrestris tadpoles: Effects of cover and predator identity. Copeia 1996, 1996, 485–488. [Google Scholar] [CrossRef]
- Soluk, D.A.; Collins, N.C. Synergistic interactions between fish and stone flies-facilitation and interference among stream predators. Oikos 1988, 52, 94–100. [Google Scholar] [CrossRef]
- Soluk, D.A. Multiple predator effects: Predicting combined functional response of stream fish and invertebrate predators. Ecology 1993, 74, 219–225. [Google Scholar] [CrossRef]
- Sih, A.; Englund, G.; Wooster, D. Emergent impacts of multiple predators on prey. Trends Ecol. Evol. 1998, 13, 350–355. [Google Scholar] [CrossRef]
- Crumrine, P.W.; Crowley, P.H. Partitioning components of risk reduction in a dragonfly-fish intraguild predation system. Ecology 2003, 84, 1588–1597. [Google Scholar] [CrossRef]
- Nishi, R.; Venkatesan, P. Predation ingestion rate and its bearing on prey death rate in Anisops bouvieri Kirkaldy. J. Entomol. Res. (New Delhi) 1989, 13, 140–145. [Google Scholar]
- Victor, T.J.; Reuben, R. Population dynamics of mosquito immatures and the successsion in abundance of aquatic insects in rice fields in Madurai, South India. Indian J. Malariol. 1999, 36, 19–32. [Google Scholar]
- Sunish, I.P.; Reuben, R. Factors influencing the abundance of Japanese encephalitis vectors in rice fields in India–II. Biotic. Med. Vet. Entomol. 2002, 16, 1–9. [Google Scholar] [CrossRef]
- Aditya, G.; Bhattacharya, S.; Kundu, N.; Saha, G.K.; Raut, S.K. Predatory efficiency of the water bug Sphaerodema annulatum on mosquito larvae (Culex quinquefasciatus) and its effect on adult emergence. Bioresour. Technol. 2004, 95, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Aditya, G.; Bhattacharya, S.; Kundu, N.; Saha, G.K. Frequency dependent prey selection of predacious water-bugs on Armigeris subalbatus immature. J. Vector Borne Dis. 2005, 42, 9–14. [Google Scholar] [PubMed]
- Saha, N.; Aditya, G.; Bal, A.; Saha, G.K. A comparative study of predation of three aquatic hemipteran bugs on Culex quinquefasciatus larvae. Limnology 2007, 8, 73–80. [Google Scholar] [CrossRef]
- Saha, N.; Aditya, G.; Bal, A.; Saha, G.K. Comparative study of functional response of common Hemipteran bugs of East Calcutta wetlands, India. Int. Rev. Hydrobiol. 2007, 92, 242–257. [Google Scholar] [CrossRef]
- Mogi, M.; Memah, V.; Miyagi, I.; Toma, T.; Sembel, D.T. Mosquito(Diptera: Culicidae) and predator abundance in irrigated and rain-fed rice-fields in North-Sulawesi, Indonesia. J. Med. Entomol. 1995, 32, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Shaalan, E.A.-S.; Canyon, D.V. Aquatic insect predators and mosquito control. Trop. Biomed. 2009, 26, 223–261. [Google Scholar]
- Juliano, S.A. Species interactions among larval mosquitoes: Context dependence across habitat gradients. Ann. Rev. Entomol. 2009, 54, 37–56. [Google Scholar] [CrossRef]
- Gilbert, J.J.; Burns, C.W. Some observations on the diet of the Backswimmer Anisops wakefieldi(Hemiptera: Notonectidae). Hydrobiologia 1999, 412, 111–118. [Google Scholar] [CrossRef]
- Hampton, S.E. Habitat overlap of enemies: Temporal patterns and the role of spatial complexity. Oecologia 2004, 138, 475–484. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis; Pearson Education (Singapore) Pte. Ltd. New Delhi (Indian Branch): New Delhi, India, 1999; 667p. [Google Scholar]
- Warfe, D.M.; Barmuta, L.A. Habitat structural complexity mediates food web dynamics in a freshwater macrophyte community. Oecologia 2006, 150, 141–154. [Google Scholar] [CrossRef]
- Saha, N.; Aditya, G.; Bal, A.; Saha, G.K. Light and habitat structure influences predation of Culex quinquefasciatus larvae by the water bugs (Hemiptera: Heteroptera). Insect Sci. 2008, 15, 461–469. [Google Scholar]
- Saha, N.; Aditya, G.; Saha, G.K. Prey preferences of aquatic insects: Potential imply cations for the regulation of wetland mosquitoes. Med. Vet. Entomol. 2014, 28, 1–9. [Google Scholar] [CrossRef]
- Saha, N.; Kundu, M.; Saha, G.K.; Aditya, G. Alternative prey influence the predation of mosquito larvae by three water bug species (Heteroptera: Nepidae). Limnol. Rev. 2020, 20, 173–184. [Google Scholar] [CrossRef]
- Fischer, S.; Zanotti, G.; Castro, A.; Quiroga, L.; Vargas, D.V. Effect of habitat complexity on the predation of Buenoa fuscipennis (Heteroptera: Notonectidae) on mosquito imamture stages and alternative prey. J. Vector. Ecol. 2013, 38, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.; Sabelis, M.W.; Magalhaes, S.; Montserrat, M.; der Hammen, T.V. Habitat structure affects intraguild predation. Ecology 2007, 88, 2713–2719. [Google Scholar] [CrossRef] [PubMed]
- Fulan, J.Â.; Almeida, S.C. Effect of the spatial heterogeneity on the predation of Scinax fuscovarius and Physalaemus cuvieri tadpoles by Odonata larvae. Acta Sci. Biol. Sci. 2010, 32, 121–124. [Google Scholar] [CrossRef]
- Swisher, B.J.; Soluk, D.A.; Wahl, D.H. Non-additive predation in littoral habitats: Influences of habitat complexity. Oikos 1998, 81, 30–37. [Google Scholar] [CrossRef]
- Hossie, T.J.; Murray, D.L. You can’t run but you can hide: Refuge use in frog tadpoles elicits density-dependent predation by dragonfly larvae. Oecologia 2010, 163, 395–404. [Google Scholar] [CrossRef]
- Anderson, T.L.; Semlitcsh, R.D. Top predator and habitat complexity alter an intraguild predation module in pond communities. J. Anim. Ecol. 2016, 85, 548–558. [Google Scholar] [CrossRef]
- Eklov, P.; van Kooten, T. Facilitation among piscivorous predators: Effects of prey habitat use. Ecology 2001, 82, 2486–2494. [Google Scholar] [CrossRef]
- Grabowsky, J.H.; Hughes, A.R.; Kimbro, D.L. Habitat complexity influences cascading effects of multiple predators. Ecology 2008, 89, 3413–3422. [Google Scholar] [CrossRef] [PubMed]
- Griffen, B.D.; Byers, J.E. Partitioning mechanisms of predator interference in different habitats. Oecologia 2006, 146, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Dambach, P. The use of aquatic predators for larval control of mosquito disease vectors. Biol. Control 2020, 150, 104357. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Details | Remarks |
|---|---|---|
| IG predator | Diplonychus rusticus | Density 2 (low) and 4 (high) individuals |
| IG prey | Anisops bouvieri | Density 10 individuals |
| Shared prey | Instar IV larvae of Culex quinquefasciatus and Chironomid midge | Density of 50 and 200 for each prey species respectively |
| IG predator: IG prey | 2:10 and 4:10 | Low and high predator density |
| Habitat | Open and complex | Simple and complex conditions |
| Days | Consecutive 9 days | Long term |
| Replicates | 6 × 2 prey densities per shared prey type per IG predator density per habitat per day | 864 replicates |
| Prey consumption | Noted after 24 h for 9 days | Shared prey and IG prey replenished everyday |
| Analysis | Factorial ANOVA on the predation data and multiplicative risk analysis [17], followed by post hoc multiple comparison tests; use of clearance rate [49,50] as a composite indicator of predation. Multiplicative risk analysis for mosquito and chironomid midges | For both IG prey and shared prey |
| A. Mosquito Larvae as Shared Prey | B. Chironomid Larvae as Shared Prey | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Within-Subject Effects (Sphericity Assumed) | Within-Subject Effects (Sphericity Assumed) | ||||||||||
| Source | Sum of Squares | df | Mean Square | F | Partial η2 | Source | Sum of Squares | df | Mean Square | F | Partial η2 |
| Days | 14,395.41 | 8 | 1799.43 | 179.73 | 0.82 | Days | 5714.96 | 8 | 714.37 | 71.35 | 0.64 |
| Days * predator density (PRDD) | 50.33 | 8 | 6.29 | 0.63 | 0.02 | Days * predator density (PRDD) | 510.99 | 8 | 63.87 | 6.38 | 0.14 |
| Days * prey density (PRD) | 2708.75 | 8 | 338.59 | 33.82 | 0.46 | Days * prey density (PRD) | 2653.69 | 8 | 331.71 | 33.13 | 0.45 |
| Days * habitat (HAB) | 567.41 | 8 | 70.93 | 7.08 | 0.15 | Days * habitat (HAB) | 45.21 | 8 | 5.65 | 0.56 | 0.01 |
| Days * PRDD * PRD | 20.99 | 8 | 2.62 | 0.26 | 0.01 | Days * PRDD * PRD | 322.92 | 8 | 40.36 | 4.03 | 0.09 |
| Days * PRDD * HAB | 42 | 8 | 5.25 | 0.52 | 0.01 | Days * PRDD * HAB | 67.8 | 8 | 8.47 | 0.85 | 0.02 |
| Days * PRD * HAB | 326.08 | 8 | 40.76 | 4.07 | 0.09 | Days * PRD * HAB | 13.88 | 8 | 1.73 | 0.17 | 0 |
| Days * PRDD * PRD * HAB | 11.25 | 8 | 1.41 | 0.14 | 0 | Days * PRDD * PRD * HAB | 60.07 | 8 | 7.51 | 0.75 | 0.02 |
| Error (days) | 3203.78 | 320 | 10.01 | Error (days) | 3204.04 | 320 | 10.01 | ||||
| Within-Subject Contrasts | Within-Subject Contrasts | ||||||||||
| Source | Sum of Squares | df | Mean Square | F | Partial η2 | Source | Sum of Squares | df | Mean Square | F | Partial η2 |
| Days | 14,226.67 | 1 | 14226.67 | 1268.7 | 0.969 | Days | 4774.05 | 1 | 4774.05 | 377.24 | 0.9 |
| Days * predator density (PRDD) | 16.05 | 1 | 16.05 | 1.43 | 0.035 | Days * predator density (PRDD) | 225.57 | 1 | 225.57 | 17.82 | 0.31 |
| Days * prey density (PRD) | 2506.93 | 1 | 2506.93 | 223.56 | 0.848 | Days * prey density (PRD) | 2405.36 | 1 | 2405.36 | 190.07 | 0.83 |
| Days * habitat (HAB) | 411.78 | 1 | 411.78 | 36.72 | 0.479 | Days * habitat (HAB) | 5.87 | 1 | 5.87 | 0.46 | 0.01 |
| Days * PRDD * PRD | 1.75 | 1 | 1.75 | 0.16 | 0.004 | Days * PRDD * PRD | 150.33 | 1 | 150.33 | 11.88 | 0.23 |
| Days * PRDD * HAB | 14.03 | 1 | 14.03 | 1.25 | 0.03 | Days * PRDD * HAB | 10.27 | 1 | 10.27 | 0.81 | 0.02 |
| Days * PRD * HAB | 0.01 | 1 | 0.01 | 0 | 0 | Days * PRD * HAB | 0.23 | 1 | 0.23 | 0.02 | 0 |
| Days * PRDD * PRD * HAB | 0.06 | 1 | 0.06 | 0.01 | 0 | Days * PRDD * PRD * HAB | 0.14 | 1 | 0.14 | 0.01 | 0 |
| Error (days) | 448.54 | 40 | 11.21 | Error (days) | 506.21 | 40 | 12.66 | ||||
| Between-Subjects Effect | Between Subjects Effect | ||||||||||
| Source | Sum of Squares | df | Mean Square | F | Partial η2 | Source | Sum of Squares | df | Mean Square | F | Partial η2 |
| Predator density (PRDD) | 901.33 | 1 | 901.33 | 51.56 | 0.56 | Predator density (PRDD) | 978.01 | 1 | 978.01 | 140.4 | 0.78 |
| Prey density (PRD) | 69,464.08 | 1 | 69464.08 | 3973.79 | 0.99 | Prey density (PRD) | 71353.48 | 1 | 71353.48 | 10,243.49 | 1 |
| Habitat (HAB) | 2987.26 | 1 | 2987.26 | 170.89 | 0.81 | Habitat (HAB) | 778.7 | 1 | 778.7 | 111.79 | 0.74 |
| PRDD * PRD | 166.26 | 1 | 166.26 | 9.51 | 0.19 | PRDD * PRD | 176.33 | 1 | 176.33 | 25.31 | 0.39 |
| PRDD * HAB | 24.08 | 1 | 24.08 | 1.38 | 0.03 | PRDD * HAB | 78.37 | 1 | 78.37 | 11.25 | 0.22 |
| PRD * HAB | 27 | 1 | 27 | 1.54 | 0.04 | PRD * HAB | 131.12 | 1 | 131.12 | 18.82 | 0.32 |
| PRDD * PRD * HAB | 0.75 | 1 | 0.75 | 0.04 | PRDD * PRD * HAB | 73.34 | 1 | 73.34 | 10.53 | 0.21 | |
| Error | 699.22 | Error | 278.63 | 40 | 6.97 | ||||||
| Multivariate Test (Wilk’s Lambda) | Multivariate Test (Wilk’s Lambda) | ||||||||||
| Effect | Value | F | Hypothesis df | Error df | Partial η2 | Effect | Value | F | Hypothesis df | Error df | Partial η2 |
| Days | 0.02 | 164.74 | 8 | 33 | 0.98 | Days | 0.06 | 70.77 | 8 | 33 | 0.94 |
| Days * predator density (PRDD) | 0.88 | 0.56 | 8 | 33 | 0.12 | Days * predator density (PRDD) | 0.36 | 7.18 | 8 | 33 | 0.64 |
| Days * prey density (PRD) | 0.11 | 34.45 | 8 | 33 | 0.89 | Days * prey density (PRD) | 0.11 | 33.59 | 8 | 33 | 0.89 |
| Days * habitat (HAB) | 0.41 | 5.97 | 8 | 33 | 0.59 | Days * habitat (HAB) | 0.87 | 0.64 | 8 | 33 | 0.13 |
| Days * PRDD * PRD | 0.92 | 0.37 | 8 | 33 | 0.08 | Days * PRDD * PRD | 0.49 | 4.23 | 8 | 33 | 0.51 |
| Days * PRDD * HAB | 0.88 | 0.55 | 8 | 33 | 0.12 | Days * PRDD * HAB | 0.86 | 0.69 | 8 | 33 | 0.14 |
| Days * PRD * HAB | 0.43 | 5.45 | 8 | 33 | 0.57 | Days * PRD * HAB | 0.96 | 0.18 | 8 | 33 | 0.04 |
| Days * PRDD * PRD * HAB | 0.95 | 0.2 | 8 | 33 | 0.05 | Days * PRDD * PRD * HAB | 0.8 | 1.01 | 8 | 33 | 0.2 |
| A. Mosquito Larvae as Shared Prey | B. Chironomid Larvae as Shared Prey | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Within-Subject Effects (Sphericity Assumed) | Within-Subject Effects (Sphericity Assumed) | ||||||||||
| Source | Sum of Squares | Df | Mean Square | F | Partial η2 | Source | Sum of Squares | df | Mean Square | F | Partial η2 |
| Days | 399.83 | 8 | 49.98 | 46.02 | 0.54 | Days | 566 | 8 | 70.75 | 73.44 | 0.65 |
| Days * predator density (PRDD) | 2.05 | 8 | 0.26 | 0.24 | 0.01 | Days * predator density (PRDD) | 3.58 | 8 | 0.45 | 0.46 | 0.01 |
| Days * prey density (PRD) | 27.88 | 8 | 3.48 | 3.21 | 0.07 | Days * prey density (PRD) | 19.24 | 8 | 2.41 | 2.5 | 0.06 |
| Days * habitat (HAB) | 0.75 | 8 | 0.09 | 0.09 | 0.002 | Days * habitat (HAB) | 9.99 | 8 | 1.25 | 1.3 | 0.03 |
| Days * PRDD * PRD | 9.35 | 8 | 1.17 | 1.08 | 0.03 | Days * PRDD * PRD | 1.71 | 8 | 0.21 | 0.22 | 0.01 |
| Days * PRDD * HAB | 0.67 | 8 | 0.08 | 0.08 | 0.002 | Days * PRDD * HAB | 0.96 | 8 | 0.12 | 0.12 | 0 |
| Days * PRD * HAB | 1.63 | 8 | 0.2 | 0.19 | 0.005 | Days * PRD * HAB | 1.25 | 8 | 0.16 | 0.16 | 0 |
| Days * PRDD * PRD * HAB | 1.66 | 8 | 0.21 | 0.19 | 0.005 | Days * PRDD * PRD * HAB | 1.67 | 8 | 0.21 | 0.22 | 0.01 |
| error (days) | 347.52 | 320 | 1.09 | error (days) | 308.26 | 320 | 0.96 | ||||
| Within-Subject Contrasts | Within-Subject Contrasts | ||||||||||
| Source | Sum of Squares | df | Mean Square | F | Partial η2 | Source | Sum of Squares | df | Mean Square | F | Partial η2 |
| Days | 358.42 | 1 | 358.42 | 356.44 | 0.9 | Days | 562.68 | 1 | 562.68 | 653.49 | 0.94 |
| Days * predator density (PRDD) | 1.01 | 1 | 1.01 | 1.01 | 0.02 | Days * predator density (PRDD) | 2.63 | 1 | 2.63 | 3.05 | 0.07 |
| Days * prey density (PRD) | 15.61 | 1 | 15.61 | 15.52 | 0.28 | Days * prey density (PRD) | 17.27 | 1 | 17.27 | 20.05 | 0.33 |
| Days * habitat (HAB) | 0.03 | 1 | 0.03 | 0.03 | 0 | Days * habitat (HAB) | 5.25 | 1 | 5.25 | 6.1 | 0.13 |
| Days * PRDD * PRD | 2.11 | 1 | 2.11 | 2.1 | 0.05 | Days * PRDD * PRD | 0.13 | 1 | 0.13 | 0.15 | 0 |
| Days * PRDD * HAB | 0.45 | 1 | 0.45 | 0.45 | 0.01 | Days * PRDD * HAB | 0.06 | 1 | 0.06 | 0.07 | 0 |
| Days * PRD * HAB | 0.73 | 1 | 0.73 | 0.73 | 0.02 | Days * PRD * HAB | 0.02 | 1 | 0.02 | 0.02 | 0 |
| Days * PRDD * PRD * HAB | 0.67 | 1 | 0.67 | 0.67 | 0.02 | Days * PRDD * PRD * HAB | 0.98 | 1 | 0.98 | 1.13 | 0.03 |
| error (days) | 40.22 | 40 | 1.01 | error (days) | 34.44 | 40 | 0.86 | ||||
| Between-Subjects Effect | Between-Subjects Effect | ||||||||||
| Source | Sum of Squares | df | Mean Square | F | Partial η2 | Source | Sum of Squares | df | Mean Square | F | Partial η2 |
| Predator density (PRDD) | 13.37 | 1 | 13.37 | 4.55 | 0.1 | Predator density (PRDD) | 18.75 | 1 | 18.75 | 8.58 | 0.18 |
| Prey density (PRD) | 29.04 | 1 | 29.04 | 9.89 | 0.2 | Prey density (PRD) | 11.34 | 1 | 11.34 | 5.19 | 0.11 |
| Habitat (HAB) | 6.75 | 1 | 6.75 | 2.3 | 0.05 | Habitat (HAB) | 11.34 | 1 | 11.34 | 5.19 | 0.11 |
| PRDD * PRD | 1.56 | 1 | 1.56 | 0.53 | 0.01 | PRDD * PRD | 1.12 | 1 | 1.12 | 0.51 | 0.01 |
| PRDD * HAB | 0.33 | 1 | 0.33 | 0.11 | 0 | PRDD * HAB | 1.12 | 1 | 1.12 | 0.51 | 0.01 |
| PRD * HAB | 0.04 | 1 | 0.04 | 0.01 | 0 | PRD * HAB | 2.08 | 1 | 2.08 | 0.95 | 0.02 |
| PRDD * PRD * HAB | 0.01 | 1 | 0.009 | 0 | 0 | PRDD * PRD * HAB | 0.08 | 1 | 0.08 | 0.04 | 0 |
| Error | 117.48 | 40 | 2.94 | Error | 87.41 | 40 | 2.19 | ||||
| Multivariate Test (Wilk’s Lambda) | Multivariate Test (Wilk’s Lambda) | ||||||||||
| Effect | Value | F | Hypothesis df | Error df | Partial η2 | Effect | Value | F | Hypothesis df | Error df | Partial η2 |
| Days | 0.08 | 47.49 | 8 | 33 | 0.92 | Days | 0.04 | 91.58 | 8 | 33 | 0.96 |
| Days * predator density (PRDD) | 0.94 | 0.27 | 8 | 33 | 0.06 | Days * predator density (PRDD) | 0.85 | 0.73 | 8 | 33 | 0.15 |
| Days * prey density (PRD) | 0.61 | 2.59 | 8 | 33 | 0.39 | Days * prey density (PRD) | 0.57 | 3.09 | 8 | 33 | 0.43 |
| Days * habitat (HAB) | 0.98 | 0.1 | 8 | 33 | 0.02 | Days * habitat (HAB) | 0.71 | 1.67 | 8 | 33 | 0.29 |
| Days * PRDD * PRD | 0.83 | 0.85 | 8 | 33 | 0.17 | Days * PRDD * PRD | 0.93 | 0.29 | 8 | 33 | 0.07 |
| Days * PRDD * HAB | 0.98 | 0.07 | 8 | 33 | 0.02 | Days * PRDD * HAB | 0.97 | 0.12 | 8 | 33 | 0.03 |
| Days * PRD * HAB | 0.96 | 0.17 | 8 | 33 | 0.04 | Days * PRD * HAB | 0.97 | 0.14 | 8 | 33 | 0.03 |
| Days * PRDD * PRD * HAB | 0.96 | 0.17 | 8 | 33 | 0.04 | Days * PRDD * PRD * HAB | 0.91 | 0.4 | 8 | 33 | 0.09 |
| A. | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shared Prey | Predatory Density | Prey Density | Days | |||||||||
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | D9 | ||||
| M | 2 | 50 | 1.60 | −0.21 | −4.43 | −4.18 | −5.00 | 0.04 | −4.55 | 0.07 | −0.93 | |
| 2 | 200 | −11.44 | −9.58 | −13.15 | −6.88 | −8.82 | −3.99 | −18.34 | −7.07 | −12.08 | ||
| 4 | 50 | −0.83 | −0.69 | −0.95 | −10.32 | −4.09 | −2.92 | −3.34 | −15.37 | −7.53 | ||
| 4 | 200 | −31.58 | −20.53 | −14.63 | −18.61 | −20.25 | −22.08 | −13.47 | −47.26 | −7.36 | ||
| C | 2 | 50 | 0.77 | 0.05 | −10.37 | −9.13 | −0.11 | −0.47 | −1.09 | 1.85 | 1.43 | |
| 2 | 200 | −10.76 | −23.14 | −14.46 | −10.51 | −6.85 | −14.84 | −11.49 | −12.36 | −7.97 | ||
| 4 | 50 | −0.89 | −6.29 | −1.20 | 0.62 | 0.61 | −1.25 | −1.06 | −3.64 | −2.13 | ||
| 4 | 200 | −23.87 | −9.21 | −14.34 | −53.24 | −10.16 | −6.97 | −9.74 | −13.53 | −5.38 | ||
| B. | ||||||||||||
| Shared Prey | Predatory Density | Predatory Density | Habitat | Days | ||||||||
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | D9 | ||||
| M | 2 | 50 | S | −13.96 | −15.57 | −12.87 | −35.79 | −20.59 | −19.54 | −44.37 | −26.40 | −90.41 |
| 2 | 50 | C | −8.89 | −7.15 | −8.14 | −7.51 | −8.88 | −12.95 | −26.76 | −18.14 | −23.19 | |
| 2 | 200 | S | −26.01 | −44.48 | −55.88 | −31.32 | −37.10 | −54.00 | −56.41 | −68.70 | −59.83 | |
| 2 | 200 | C | −17.69 | −16.27 | −31.33 | −36.27 | −20.48 | −15.99 | −46.64 | −34.87 | −26.33 | |
| 4 | 50 | S | −18.66 | −18.99 | −22.06 | −37.20 | −23.25 | −22.90 | −22.32 | −22.99 | −84.63 | |
| 4 | 50 | C | −8.55 | −11.84 | −9.93 | −9.52 | −12.31 | −13.96 | −55.29 | −1.85 | −4.77 | |
| 4 | 200 | S | −36.55 | −110.27 | −127.77 | −107.96 | −80.31 | −72.83 | −69.03 | −175.05 | −95.50 | |
| 4 | 200 | C | −4.94 | −6.71 | −9.81 | −10.84 | −18.22 | −31.80 | −12.43 | −65.63 | −14.27 | |
| C | 2 | 50 | S | −13.96 | −31.59 | −111.96 | −40.64 | −64.05 | −32.20 | −48.85 | −33.89 | −25.57 |
| 2 | 50 | C | −30.52 | −29.80 | −22.49 | −30.34 | −24.43 | −56.95 | −35.75 | −20.26 | −47.04 | |
| 2 | 200 | S | −29.75 | −65.17 | −33.73 | −75.55 | −82.65 | −46.17 | −35.72 | −33.58 | −57.09 | |
| 2 | 200 | C | −10.77 | −23.17 | −12.71 | −21.81 | −13.87 | −55.00 | −13.67 | −14.82 | −14.43 | |
| 4 | 50 | S | −52.21 | −48.76 | −48.85 | −42.79 | −25.35 | −53.83 | −49.84 | −55.50 | −48.59 | |
| 4 | 50 | C | −17.35 | −31.61 | −43.71 | −25.12 | −25.41 | −32.98 | −34.56 | −38.47 | −32.07 | |
| 4 | 200 | S | −58.66 | −34.62 | −83.69 | −96.11 | −37.61 | −96.20 | −92.02 | −116.45 | −84.37 | |
| 4 | 200 | C | −12.31 | −13.86 | −26.90 | −39.60 | −22.41 | −21.74 | −25.93 | −50.08 | −25.90 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brahma, S.; Sharma, D.; Saha, G.K.; Aditya, G. Dipteran Prey Vulnerability in Intraguild Predation (IGP) System Involving Heteropteran Predators: Density and Habitat Effects. Limnol. Rev. 2023, 23, 1-20. https://doi.org/10.3390/limnolrev23010001
Brahma S, Sharma D, Saha GK, Aditya G. Dipteran Prey Vulnerability in Intraguild Predation (IGP) System Involving Heteropteran Predators: Density and Habitat Effects. Limnological Review. 2023; 23(1):1-20. https://doi.org/10.3390/limnolrev23010001
Chicago/Turabian StyleBrahma, Shreya, Dipendra Sharma, Goutam K. Saha, and Gautam Aditya. 2023. "Dipteran Prey Vulnerability in Intraguild Predation (IGP) System Involving Heteropteran Predators: Density and Habitat Effects" Limnological Review 23, no. 1: 1-20. https://doi.org/10.3390/limnolrev23010001