
Estimating Surface EMG Activity of Human Upper Arm Muscles Using InterCriteria Analysis


Abstract
:1. Introduction
2. Methods
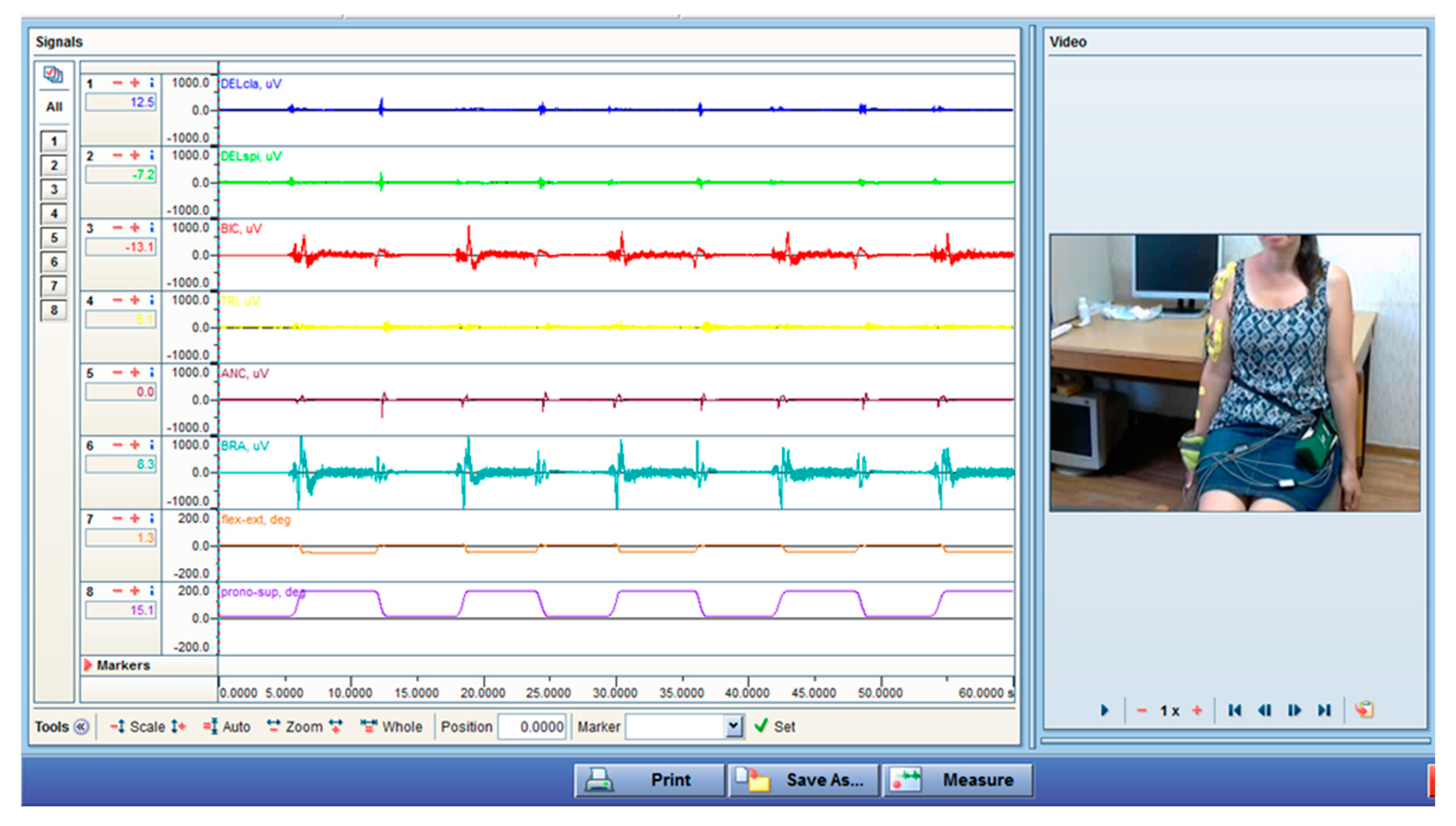
2.1. EMG Method
- Rest position. Both arms were in a relaxed position beside the body. The participant held that position for one minute.
- Maximal isometric contractions. The subject was asked to assume several positions of the elbow and shoulder. The examiner applied adequate force to provoke separately the maximum isometric effort of the investigated muscles.
- Flexion in the sagittal plane. From the rest position, the participant performed some cycles of full-range elbow flexion in the sagittal plane: full flexion; a 5 s rest period in the reaching position; an extension to the initial position; a 5 s rest period in the reaching position. These movement cycles were repeated for a minute. These motions were performed with four different velocities, from very slow to fastest, and these velocities were controlled by a computer tabata program (each change from motion to the held position is regulated by sound and visual markers on the computer’s monitor). The time durations for flexion and extension were 10 s (1flex/1ext), 6 s (2flex/2ext), 2 s (3flex/3ext), and 1 s (4flex/4ext) consecutively. The symbols in the parenthesis are the abbreviations used in this paper. The rest at the end position was 5 s.
- Flexion in the sagittal plane with added weight. A wristband with a weight of 0.5 kg was placed at the wrist, and the same flexion-rest-extension tasks were performed. Periods for flexion and extension were 10 s (1flexW/1extW), 6 s (2flexW/2extW), 2 s (3flexW/3extW), and 1 s (4flexW/4extW). As noted above, the accepted abbreviations are given in parentheses.
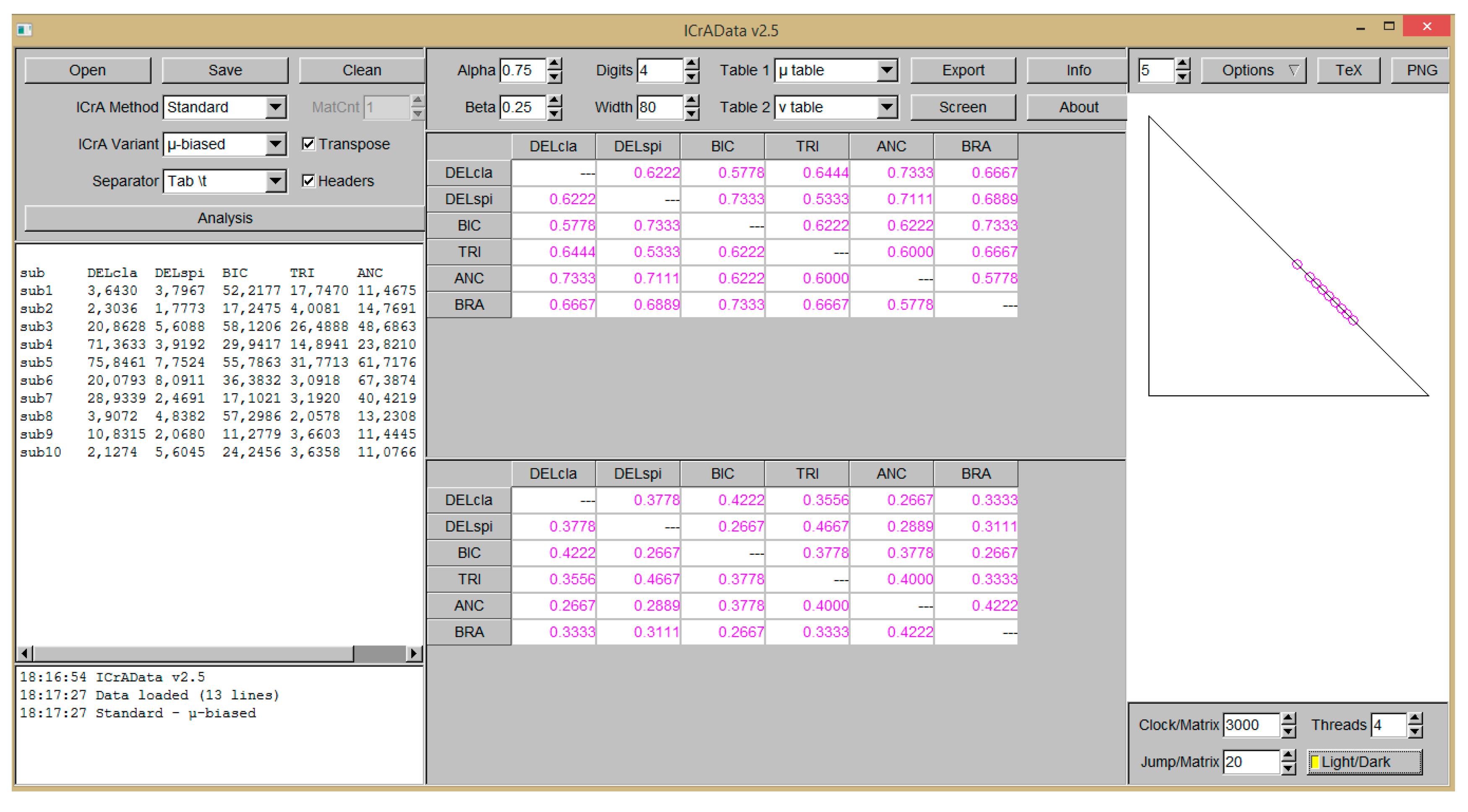
2.2. InterCriteria Analysis Approach
| A = | O1 | … | Oi | … | Oj | … | Ok | |
| C1 | eC1,O1 | … | eC1,Oi | … | eC1,Oj | … | eC1,Ok | |
| … | … | … | … | … | … | … | … | |
| Cp | eCp,O1 | … | eCp,Oi | … | eCp,Oj | … | eCp,Ok | |
| … | … | … | … | … | … | … | … | |
| Cq | eCq,O1 | … | eCq,Oi | … | eCq,Oj | … | eCq,Ok | |
| … | … | … | … | … | … | … | … | |
| Cm | eCm,O1 | … | eCm,Oi | … | eCm,Oj | … | eCm,Ok |
| A* = | C1 | … | Cm | |
| C1 | … | |||
| … | … | … | … | |
| Cm | … |
3. Results
3.1. Results for Flexion with Different Velocities with and without an Additional Load in the Sagittal Plane after ICrA Application
3.2. Results for Extension with Different Velocities with and without an Additional Load in Sagittal Plane after ICrA Application
3.3. Results for Examined Subjects Performing Flexion and Extension Movements after ICrA Application
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cheung, V.C.; d’Avella, A.; Tresch, M.C.; Bizzi, E. Central and sensory contributions to activating and organizing muscle synergies during natural motor behaviors. J. Neurosci. 2005, 25, 6419–6434. [Google Scholar] [CrossRef]
- Tresch, M.C.; Bizzi, E. Responses to spinal microstimulation in the chronically spinalized rat and their relationship to spinal systems activated by low threshold cutaneous stimulation. Exp. Brain Res. 1999, 129, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Mussa-Ivaldi, F.A.; Giszter, S.F.; Bizzi, E. Linear combinations of primitives in vertebrate motor control. Proc. Natl. Acad. Sci. USA 1994, 91, 7534–7538. [Google Scholar] [CrossRef]
- Bizzi, E.; Mussa-Ivaldi, F.A.; Giszter, S.F. Computations underlying the execution of movement: A biological perspective. Science 1991, 253, 287–291. [Google Scholar] [CrossRef] [PubMed]
- d’Avella, A.; Bizzi, E. Shared and specific muscle synergies in natural motor behaviors. Proc. Natl. Acad. Sci. USA 2005, 102, 3076–3081. [Google Scholar] [CrossRef] [PubMed]
- Ivanenko, Y.P.; Cappellini, G.; Dominici, N.; Poppele, R.E.; Lacquaniti, F. Coordination of locomotion with voluntary movements in humans. J. Neurosci. 2005, 25, 7238–7253. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Pérez, D.C.; Rodríguez-Reséndiz, J.; Gómez-Loenzo, R.A.; Jauregui-Correa, J.C. Support Vector Machine-Based EMG Signal Classification Techniques: A Review. Appl. Sci. 2019, 9, 4402. [Google Scholar] [CrossRef]
- Aviles, M.; Rodríguez-Reséndiz, J.; Ibrahimi, D. Optimizing EMG Classification through Metaheuristic Algorithms. Technologies 2023, 11, 87. [Google Scholar] [CrossRef]
- Aviles, M.; Sánchez-Reyes, L.-M.; Fuentes-Aguilar, R.Q.; Toledo-Pérez, D.C.; Rodríguez-Reséndiz, J. A Novel Methodology for Classifying EMG Movements Based on SVM and Genetic Algorithms. Micromachines 2022, 13, 2108. [Google Scholar] [CrossRef]
- Toledo-Pérez, D.C.; Rodríguez-Reséndiz, J.; Gómez-Loenzo, R.A. A Study of Computing Zero Crossing Methods and an Improved Proposal for EMG Signals. IEEE Access 2020, 8, 8783–8790. [Google Scholar] [CrossRef]
- D’Avella, A.; Portone, A.; Fernandez, L.; Lacquaniti, F. Control of Fast-Reaching Movements by Muscle Synergy Combinations. J. Neurosci. 2006, 26, 7791–7810. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, T.S.; Rovai, G.P.; Rymer, W.Z. Strategies for muscle activation during isometric torque generation at the human elbow. J. Neurophysiol. 1989, 62, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Jamison, J.C.; Caldwell, G.E. Muscle synergies and isometric torque production: Influence of supination and pronation level on elbow flexion. J. Neurophysiol. 1993, 70, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Atanassov, K.; Mavrov, D.; Atanassova, V. Intercriteria Decision Making: A New Approach for Multicriteria Decision Making, Based on Index Matrices and Intuitionistic Fuzzy Sets. Issues Intuitionistic Fuzzy Sets Gen. Nets 2014, 11, 1–8. [Google Scholar]
- Todinova, S.; Mavrov, D.; Krumova, S.; Marinov, P.; Atanassova, V.; Atanassov, K.; Taneva, S.G. Blood plasma thermograms dataset analysis by means of intercriteria and correlation analyses for the case of colorectal cancer. Int. J. Bioautom. 2016, 20, 115–124. [Google Scholar]
- Jekova, I.; Vassilev, P.; Stoyanov, T.; Pencheva, T. InterCriteria Analysis: Application for ECG Data Analysis. Mathematics 2021, 9, 854. [Google Scholar] [CrossRef]
- Andreev, N.; Attanassov, K.; Bureva, V. New InterCriteria analysis on blood collection data. Annu. Inform. Sect. Union Sci. 2020, 10, 30–53. (In Bulgarian) [Google Scholar]
- Jereva, D.; Angelova, M.; Tsakovska, I.; Alov, P.; Pajeva, I.; Miteva, M.; Pencheva, T. InterCriteria Analysis Approach for Decision-making in Virtual Screening: Comparative Study of Various Scoring Functions. In Lecture Notes in Networks and Systems; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Jereva, D.; Alov, P.; Tsakovska, I.; Angelova, M.; Atanassova, V.; Vassilev, P.; Ikonomov, N.; Atanassov, K.; Pajeva, I.; Pencheva, T. Application of InterCriteria Analysis to Assess the Performance of Scoring Functions in Molecular Docking Software Packages. Mathematics 2022, 10, 2549. [Google Scholar] [CrossRef]
- Ilkova, T.; Petrov, M. InterCriteria analysis for evaluation of the pollution of the Struma river in the Bulgarian section. Notes IFSs 2016, 22, 120–130. [Google Scholar]
- Georgieva, V.; Angelova, N.; Roeva, O.; Pencheva, T. Intercriteria analysis of wastewater treatment quality. J. Int. Sci. Publ. Ecol. Saf. 2016, 10, 365–376. [Google Scholar]
- Angelova, M. InterCriteria Analysis of Control Parameters Relations in Artificial Bee Colony Algorithm. WSEAS Trans. Math. 2019, 18, 123–128. [Google Scholar]
- Fidanova, S.; Roeva, O.; Luque, G.; Paprzycki, M. InterCriteria analysis of different hybrid ant colony optimization algorithms for workforce planning. In Recent Advances in Computational Optimization. Studies in Computational Intelligence; Fidanova, S., Ed.; Springer: Cham, Switzerland, 2020; Volume 838, pp. 61–81. [Google Scholar]
- Mucherino, A.; Fidanova, S.; Ganzha, M. Ant Colony Optimization with environment changes: An application to GPS surveying. In Proceedings of the 2015 Federated Conference on Computer Science and Information Systems (FedCSIS), Lodz, Poland, 13–16 September 2015; Volume 5, pp. 495–500. [Google Scholar] [CrossRef]
- Krawczak, M.; Bureva, V.; Sotirova, E.; Szmidt, E. Application of the intercriteria decision making method to universities ranking. Adv. Intell. Syst. Comput. 2016, 401, 365–372. [Google Scholar]
- Available online: http://www.seniam.org/ (accessed on 9 November 2023).
- Hug, F. Can muscle coordination be precisely studied by surface electromyography? J. Electromyogr. Kinesiol. 2011, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Martinek, R.; Ladrova, M.; Sidikova, M.; Jaros, R.; Behbehani, K.; Kahankova, R.; Kawala-Sterniuk, A. Advanced Bioelectrical Signal Processing Methods: Past, Present, and Future Approach—Part III: Other Biosignals. Sensors 2021, 21, 6064. [Google Scholar] [CrossRef] [PubMed]
- Ikonomov, N.; Vassilev, P.; Roeva, O. ICrAData—Software for InterCriteria Analysis. Int. J. Bioautom. 2018, 22, 1–10. [Google Scholar] [CrossRef]
- Basmagian, J. Muscles Alive Their Functions Revealed by Electromyography. 1967. Available online: https://archive.org/details/basmajian-muscles-alive-their-functions-revealed-by-electromyography/page/176/mode/2up (accessed on 9 November 2023).
- Diplock, B.; Hing, W.; Marks, D. The long head of biceps at the shoulder: A scoping review. BMC Musculoskelet. Disord. 2023, 24, 232. [Google Scholar] [CrossRef] [PubMed]
- Papaxanthis, C.; Pozzo, T.; Stapley, P. Effects of movement direction upon kinematic characteristics of vertical arm pointing movements in man. Neurosci. Lett. 1998, 253, 103–106. [Google Scholar] [CrossRef]
- Kim, P.-T.; Isogai, S.; Murakami, G.; Wada, T.; Aoki, M.; Yamashita, T.; Ishii, S. The Lateral Collateral Ligament Complex and Related Muscles Act as a Dynamic Stabilizer as well as a Static Supporting Structure at the Elbow Joint: An Anatomical and Experimental Study. Okajimas Folia Anat. Jpn. 2002, 79, 55–61. [Google Scholar] [CrossRef]
- Basmajian, J.V.; Latif, A. Archive Integrated Actions and Functions of the Chief Flexors of the Elbow. A Detailed Electromyographic Analysis. J. Bone Joint Surg. Am. 1957, 39, 1106–1118. [Google Scholar] [CrossRef]
- Naito, A.; Yajima, M.; Fukamachi, H.; Ushikoshit, K.; Sun, Y.-J.; Shimiz, Y. Electromyographic (EMG) study of the elbow flexors during supination and pronation of the forearm. Tohoku J. Exp. Med. 1998, 186, 267–277. [Google Scholar] [CrossRef]
- Naito, A.; Shimizu, Y.; Handa, Y.; Ichie, M.; Hoshimiya, N. Functional Anatomical Studies of the Elbow Movements I. Electromyographic (EMG) Analysis. Okajimas Folia Anat. 1991, 68, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Baratta, R.; Solomonow, M.; Zhou, B.H.; Letson, D.; Chuinard, R.; D’Ambrosia, R. Muscular coactivation: The role of the antagonist musculature in maintaining knee stability. Am. J. Sports Med. 1998, 16, 113–122. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.J.; Mambrito, B. Voluntary control of motor units in the human antagonist muscles: Coactivation and reciprocal activation. J. Neurophysiol. 1987, 58, 525–542. [Google Scholar] [CrossRef] [PubMed]
- GRAY’S Anatomy: The Anatomical Basis of Clinical Practice. Susan Standring, 41st ed.; Elsevier Limited: Amsterdam, The Netherlands, 2016.
- Villaseñor-Ovies, P.; Vargas, A.; Chiapas-Gasca, K.; Canoso, J.J.; Hernández-Díaz, C.; Saavedra, M.Á.; Navarro-Zarza, J.E.; Kalishd, R.A. Clinical Anatomy of the Elbow and Shoulder. Anatomía Clínica Del Hombro Y El Codo 2012, 8, 13–24. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Meaning of Consonance and Dissonance According to µ-Values |
|---|
| (0.95, 1.00]—strong positive consonance |
| (0.85, 0.95]—positive consonance |
| (0.75, 0.85]—weak positive consonance |
| (0.67, 0.75]—weak dissonance |
| (0.57, 0.67]—dissonance |
| (0.43, 0.57]—strong dissonance |
| (0.33, 0.43]—dissonance |
| (0.25, 0.33]—weak dissonance |
| (0.15, 0.25]—weak negative consonance |
| (0.05, 0.15]—negative consonance |
| [0.00, 0.05]—strong negative consonance |
| Flexion | 1flex | 2flex | 3flex | 4flex | 1flexW | 2flexW | 3flexW | 4flexW |
|---|---|---|---|---|---|---|---|---|
| DELcla-DELspi | 0.51 | 0.49 | 0.47 | 0.51 | 0.67 | 0.56 | 0.62 | 0.53 |
| DELcla-BIC | 0.47 | 0.53 | 0.53 | 0.62 | 0.62 | 0.56 | 0.58 | 0.49 |
| DELcla-TRI | 0.56 | 0.69 | 0.64 | 0.73 | 0.73 | 0.60 | 0.64 | 0.69 |
| DELcla-ANC | 0.62 | 0.73 | 0.80 | 0.76 | 0.49 | 0.62 | 0.73 | 0.76 |
| DELcla-BRA | 0.44 | 0.60 | 0.49 | 0.62 | 0.73 | 0.56 | 0.67 | 0.44 |
| DELspi-BIC | 0.51 | 0.73 | 0.76 | 0.62 | 0.78 | 0.78 | 0.73 | 0.69 |
| DELspi-TRI | 0.60 | 0.67 | 0.56 | 0.56 | 0.67 | 0.64 | 0.53 | 0.62 |
| DELspi-ANC | 0.67 | 0.71 | 0.62 | 0.67 | 0.42 | 0.62 | 0.71 | 0.69 |
| DELspi-BRA | 0.49 | 0.67 | 0.67 | 0.67 | 0.80 | 0.87 | 0.69 | 0.64 |
| BIC-TRI | 0.73 | 0.80 | 0.67 | 0.67 | 0.62 | 0.69 | 0.62 | 0.62 |
| BIC-ANC | 0.62 | 0.62 | 0.64 | 0.64 | 0.51 | 0.62 | 0.62 | 0.56 |
| BIC-BRA | 0.84 | 0.84 | 0.82 | 0.73 | 0.76 | 0.78 | 0.73 | 0.73 |
| TRI-ANC | 0.71 | 0.69 | 0.62 | 0.58 | 0.53 | 0.53 | 0.60 | 0.67 |
| TRI-BRA | 0.80 | 0.82 | 0.71 | 0.62 | 0.73 | 0.69 | 0.67 | 0.53 |
| ANC-BRA | 0.60 | 0.60 | 0.56 | 0.60 | 0.62 | 0.58 | 0.58 | 0.56 |
| Consonance/Dissonance | Number of Muscle Pairs in Consonance/Dissonance | Interacting Muscles and the Number of Detected Cases in Consonance |
|---|---|---|
| Positive consonance: (0.75, 0.85]—weak positive consonance | 16 | DELcla-ANC–3; DELspi-BIC–3; DELspi-BRA–2; BIC-TRI–1; BIC-BRA–5; TRI-BRA–2; |
| Dissonance | 104 | - |
| Negative consonance | 0 | - |
| Extension | 1ext | 2ext | 3ext | 4ext | 1extW | 2extW | 3extW | 4extW |
|---|---|---|---|---|---|---|---|---|
| DELcla-DELspi | 0.51 | 0.40 | 0.36 | 0.40 | 0.47 | 0.58 | 0.60 | 0.62 |
| DELcla-BIC | 0.47 | 0.49 | 0.56 | 0.60 | 0.58 | 0.49 | 0.56 | 0.58 |
| DELcla-TRI | 0.58 | 0.67 | 0.73 | 0.71 | 0.71 | 0.62 | 0.80 | 0.69 |
| DELcla-ANC | 0.69 | 0.60 | 0.53 | 0.64 | 0.64 | 0.62 | 0.64 | 0.62 |
| DELcla-BRA | 0.69 | 0.56 | 0.60 | 0.64 | 0.73 | 0.71 | 0.78 | 0.67 |
| DELspi-BIC | 0.56 | 0.69 | 0.71 | 0.62 | 0.67 | 0.69 | 0.73 | 0.78 |
| DELspi-TRI | 0.80 | 0.64 | 0.53 | 0.64 | 0.58 | 0.64 | 0.76 | 0.80 |
| DELspi-ANC | 0.73 | 0.58 | 0.38 | 0.62 | 0.51 | 0.51 | 0.51 | 0.60 |
| DELspi-BRA | 0.69 | 0.67 | 0.44 | 0.53 | 0.64 | 0.78 | 0.69 | 0.69 |
| BIC-TRI | 0.71 | 0.73 | 0.64 | 0.76 | 0.60 | 0.60 | 0.67 | 0.71 |
| BIC-ANC | 0.60 | 0.53 | 0.44 | 0.60 | 0.53 | 0.51 | 0.47 | 0.69 |
| BIC-BRA | 0.60 | 0.67 | 0.64 | 0.69 | 0.71 | 0.69 | 0.73 | 0.69 |
| TRI-ANC | 0.71 | 0.71 | 0.49 | 0.53 | 0.62 | 0.60 | 0.58 | 0.53 |
| TRI-BRA | 0.80 | 0.80 | 0.60 | 0.62 | 0.67 | 0.60 | 0.76 | 0.67 |
| ANC-BRA | 0.69 | 0.60 | 0.53 | 0.56 | 0.60 | 0.64 | 0.56 | 0.64 |
| Consonance/Dissonance | Number of Muscle Pairs in Consonance/Dissonance | Interacting Muscles and the Number of Detected Cases in Consonance |
|---|---|---|
| Positive consonance: (0.75, 0.85]—weak positive consonance | 11 | DELcla-TRI–1; DELcla-BRA–1; DELspi-BIC–1; DELspi-TRI–3; DELspi-BRA–1; BIC-TRI–1; TRI-BRA–3; |
| Dissonance | 109 | - |
| Negative consonance | 0 | - |
| Flexion | sub1 | sub2 | sub3 | sub4 | sub5 | sub6 | sub7 | sub8 | sub9 | sub10 |
|---|---|---|---|---|---|---|---|---|---|---|
| DELcla-DELspi | 0.93 | 0.64 | 0.82 | 0.89 | 0.79 | 0.89 | 0.68 | 0.71 | 0.89 | 0.79 |
| DELcla-BIC | 0.5 | 0.64 | 0.75 | 0.75 | 0.57 | 0.89 | 0.68 | 0.89 | 0.68 | 0.25 |
| DELcla-TRI | 0.5 | 0.61 | 0.86 | 0.75 | 0.57 | 0.50 | 0.79 | 0.82 | 0.61 | 0.57 |
| DELcla-ANC | 0.32 | 0.86 | 0.61 | 0.79 | 0.54 | 0.86 | 0.82 | 0.86 | 0.50 | 0.68 |
| DELcla-BRA | 0.43 | 0.71 | 0.87 | 0.68 | 0.54 | 0.93 | 0.79 | 0.89 | 0.64 | 0.21 |
| DELspi-BIC | 0.5 | 0.64 | 0.71 | 0.86 | 0.58 | 0.86 | 0.57 | 0.75 | 0.79 | 0.32 |
| DELspi-TRI | 0.5 | 0.68 | 0.75 | 0.86 | 0.64 | 0.54 | 0.68 | 0.89 | 0.71 | 0.64 |
| DELspi-ANC | 0.32 | 0.57 | 0.5 | 0.89 | 0.32 | 0.96 | 0.79 | 0.79 | 0.61 | 0.68 |
| DELspi-BRA | 0.43 | 0.57 | 0.82 | 0.71 | 0.61 | 0.89 | 0.68 | 0.82 | 0.75 | 0.29 |
| BIC-TRI | 0.93 | 0.96 | 0.89 | 1 | 0.93 | 0.54 | 0.89 | 0.86 | 0.93 | 0.61 |
| BIC-ANC | 0.82 | 0.64 | 0.79 | 0.89 | 0.61 | 0.89 | 0.5 | 0.83 | 0.75 | 0.29 |
| BIC-BRA | 0.93 | 0.43 | 0.89 | 0.86 | 0.96 | 0.96 | 0.89 | 0.93 | 0.89 | 0.96 |
| TRI-ANC | 0.82 | 0.68 | 0.68 | 0.89 | 0.61 | 0.57 | 0.61 | 0.82 | 0.82 | 0.68 |
| TRI-BRA | 0.93 | 0.46 | 0.93 | 0.86 | 0.96 | 0.57 | 1 | 0.86 | 0.96 | 0.57 |
| ANC-BRA | 0.89 | 0.79 | 0.68 | 0.75 | 0.57 | 0.93 | 0.61 | 0.82 | 0.86 | 0.25 |
| Flexion | Consonances |
|---|---|
| DELcla-DELspi | 7/10 |
| DELcla-BIC | 2/10 |
| DELcla-TRI | 3/10 |
| DELcla-ANC | 5/10 |
| DELcla-BRA | 5/10 |
| DELspi-BIC | 3/10 |
| DELspi-TRI | 2/10 |
| DELspi-ANC | 4/10 |
| DELspi-BRA | 3/10 |
| BIC-TRI | 8/10 |
| BIC-ANC | 5/10 |
| BIC-BRA | 9/10 |
| TRI-ANC | 4/10 |
| TRI-BRA | 7/10 |
| ANC-BRA | 5/10 |
| Extension | sub1 | sub2 | sub3 | sub4 | sub5 | sub6 | sub7 | sub8 | sub9 | sub10 |
|---|---|---|---|---|---|---|---|---|---|---|
| DELcla-DELspi | 0.86 | 0.50 | 0.89 | 0.93 | 0.86 | 0.79 | 0.64 | 0.50 | 0.82 | 0.46 |
| DELcla-BIC | 0.64 | 0.25 | 0.82 | 0.82 | 0.57 | 0.96 | 0.75 | 0.54 | 0.61 | 0.68 |
| DELcla-TRI | 0.68 | 0.54 | 0.75 | 0.68 | 0.89 | 0.61 | 0.50 | 0.54 | 0.86 | 0.61 |
| DELcla-ANC | 0.71 | 0.71 | 0.68 | 0.79 | 0.36 | 0.79 | 0.79 | 0.64 | 0.75 | 0.43 |
| DELcla-BRA | 0.82 | 0.75 | 0.75 | 0.75 | 0.64 | 0.89 | 0.68 | 0.61 | 0.75 | 0.68 |
| DELspi-BIC | 0.57 | 0.68 | 0.86 | 0.75 | 0.64 | 0.82 | 0.46 | 0.25 | 0.64 | 0.21 |
| DELspi-TRI | 0.82 | 0.68 | 0.71 | 0.68 | 0.82 | 0.61 | 0.79 | 0.82 | 0.75 | 0.57 |
| DELspi-ANC | 0.71 | 0.36 | 0.57 | 0.86 | 0.29 | 0.79 | 0.50 | 0.71 | 0.64 | 0.89 |
| DELspi-BRA | 0.82 | 0.32 | 0.86 | 0.68 | 0.64 | 0.75 | 0.89 | 0.54 | 0.78 | 0.21 |
| BIC-TRI | 0.39 | 0.57 | 0.86 | 0.71 | 0.54 | 0.57 | 0.25 | 0.14 | 0.53 | 0.36 |
| BIC-ANC | 0.36 | 0.25 | 0.57 | 0.68 | 0.21 | 0.82 | 0.68 | 0.18 | 0.43 | 0.25 |
| BIC-BRA | 0.68 | 0.14 | 0.71 | 0.86 | 0.79 | 0.93 | 0.57 | 0.36 | 0.50 | 1 |
| TRI-ANC | 0.82 | 0.68 | 0.57 | 0.54 | 0.32 | 0.75 | 0.50 | 0.82 | 0.89 | 0.54 |
| TRI-BRA | 0.71 | 0.50 | 0.64 | 0.79 | 0.68 | 0.57 | 0.68 | 0.71 | 0.89 | 0.36 |
| ANC-BRA | 0.68 | 0.75 | 0.50 | 0.68 | 0.21 | 0.82 | 0.61 | 0.61 | 0.86 | 0.25 |
| Extension | Consonances |
|---|---|
| DELcla-DELspi | 6/10 |
| DELcla-BIC | 3/10 |
| DELcla-TRI | 2/10 |
| DELcla-ANC | 3/10 |
| DELcla-BRA | 2/10 |
| DELspi-BIC | 3/10 |
| DELspi-TRI | 4/10 |
| DELspi-ANC | 3/10 |
| DELspi-BRA | 4/10 |
| BIC-TRI | 2/10 |
| BIC-ANC | 3/10 |
| BIC-BRA | 5/10 |
| TRI-ANC | 3/10 |
| TRI-BRA | 2/10 |
| ANC-BRA | 3/10 |
| Movement | Antagonistic Muscle Interaction | Agonistic Muscle Interaction | Muscle Pair Function in the Sagittal Plane |
|---|---|---|---|
| Flexion, extension | DELcla-DELspi (one-joint muscles) Acts together in the shoulder joint | The three parts of the m. deltoideus are active in all movements of the arm [30,37,38]. They are considered to be dynamic stabilizers along with rotary cuff muscles and the long head of the m. biceps brachii [40]. Anterior fibers of m. deltoideus have an assistive function in drawing the arms forwards, and posterior fibers act with m. latissimus dorsi and m. teres major in drawing the arm into extension [39]. | |
| Flexion | BIC-TRI (two-joint muscles) Acts together in the shoulder and elbow joint | These two muscles are two-joint muscles and act in both the elbow and shoulder joints but in different directions. The BIC is involved in anterior stability of the elbow in the sagittal plane. The posterior stability is enhanced by the m. triceps brachii tendon [31,39]. In the shoulder, the long head of m. triceps brachii keeps the humeral head in the glenoid cavity. It assists in the extension of the shoulder joint. The BIC weakly assists the arm movements at the glenohumeral joint in forward flexion. | |
| Flexion | BIC-BRA (two-joint muscle and one-joint muscle) Acts together in the elbow joint | BIC and BRA are flexors in the elbow joints. BIC is a flexor in a neutral position in the presence of added weight, and BRA is active during flexion in all positions of the forearm [34]. Again, according to these authors, both muscles act simultaneously and are most active in weight-bearing flexion in the neutral position of the forearm. In addition to this statement, Naito et al. [35] demonstrate a clear decrease in m. brachialis and m. brachioradialis activity together with an increase in m. biceps brachii activity during rapid prono-supination movements at the elbow from different positions. However, the authors outline the existence of ingenious reciprocal connections between the elbow flexors, which also confirms the sustained interaction shown as a result here between BIC and BRA. | |
| Flexion | TRI-BRA (two-joint muscle and one-joint muscle) Acts together in the elbow joint | The BRA is highly active in flexion in all forearm positions [30]. The long head of the triceps is the least active in the extension direction compared to the other two heads [36]. The m. brachialis is involved in anterior elbow stabilization in the sagittal plane, the triceps tendon supports the posterior [39]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelova, S.; Angelova, M.; Raikova, R. Estimating Surface EMG Activity of Human Upper Arm Muscles Using InterCriteria Analysis. Math. Comput. Appl. 2024, 29, 8. https://doi.org/10.3390/mca29010008
Angelova S, Angelova M, Raikova R. Estimating Surface EMG Activity of Human Upper Arm Muscles Using InterCriteria Analysis. Mathematical and Computational Applications. 2024; 29(1):8. https://doi.org/10.3390/mca29010008
Chicago/Turabian StyleAngelova, Silvija, Maria Angelova, and Rositsa Raikova. 2024. "Estimating Surface EMG Activity of Human Upper Arm Muscles Using InterCriteria Analysis" Mathematical and Computational Applications 29, no. 1: 8. https://doi.org/10.3390/mca29010008