
Application of Encapsulated Quorum Quenching Strain Acinetobacter pittii HITSZ001 to a Membrane Bioreactor for Biofouling Control

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
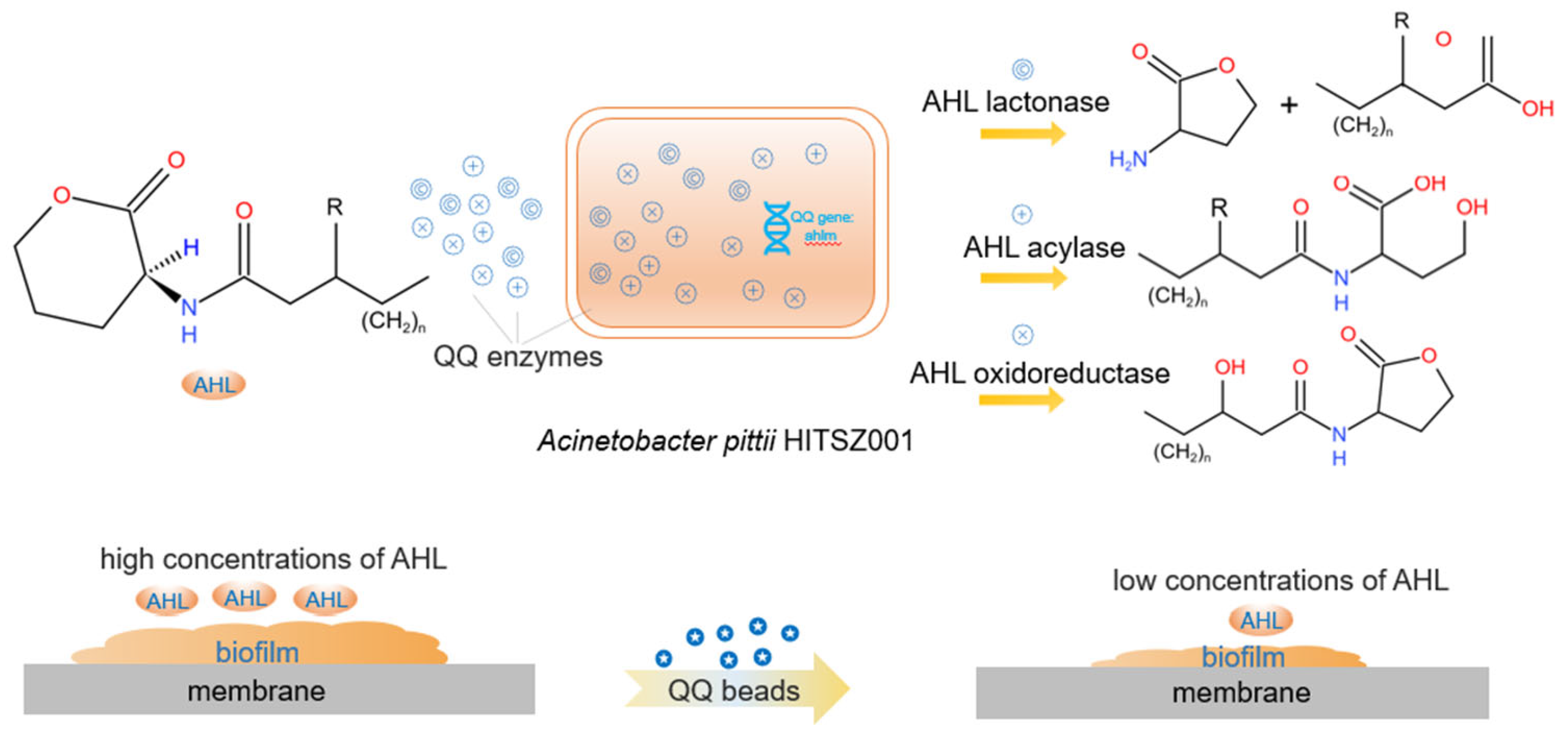
2.1. Isolation and Identification of QQ Bacteria
2.2. Immobilization of Quorum Quenching Strain
2.3. MBR System
2.4. Biomass Assay
2.5. Extraction of EPS and SMP
2.6. Analytical Methods
2.7. High-Throughput Sequencing
2.8. Statistical Analyses
3. Results and Discussion
3.1. Biofouling Inhibition by Quorum Quenching in MBR System
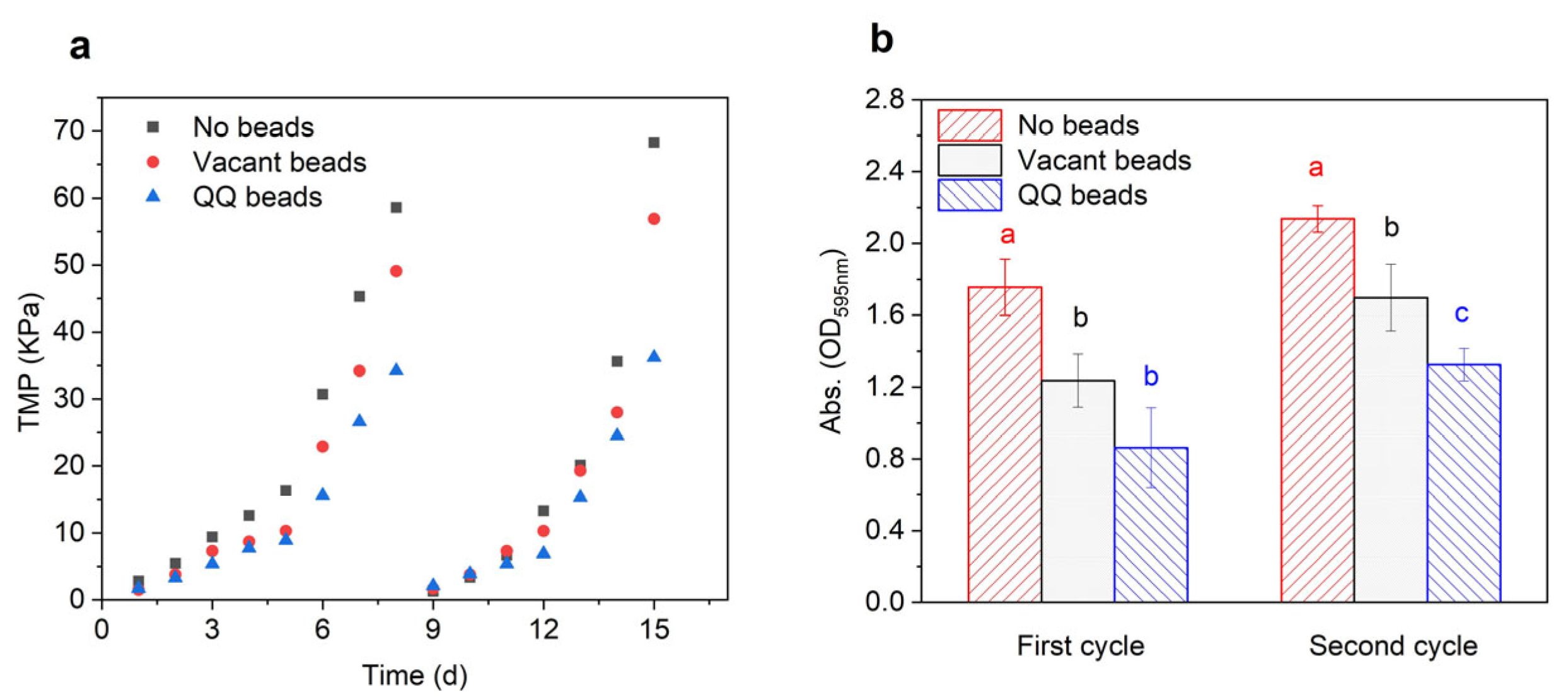
3.1.1. Variations of Trans-Membrane Pressure (TMP)
3.1.2. Variations in Biomass
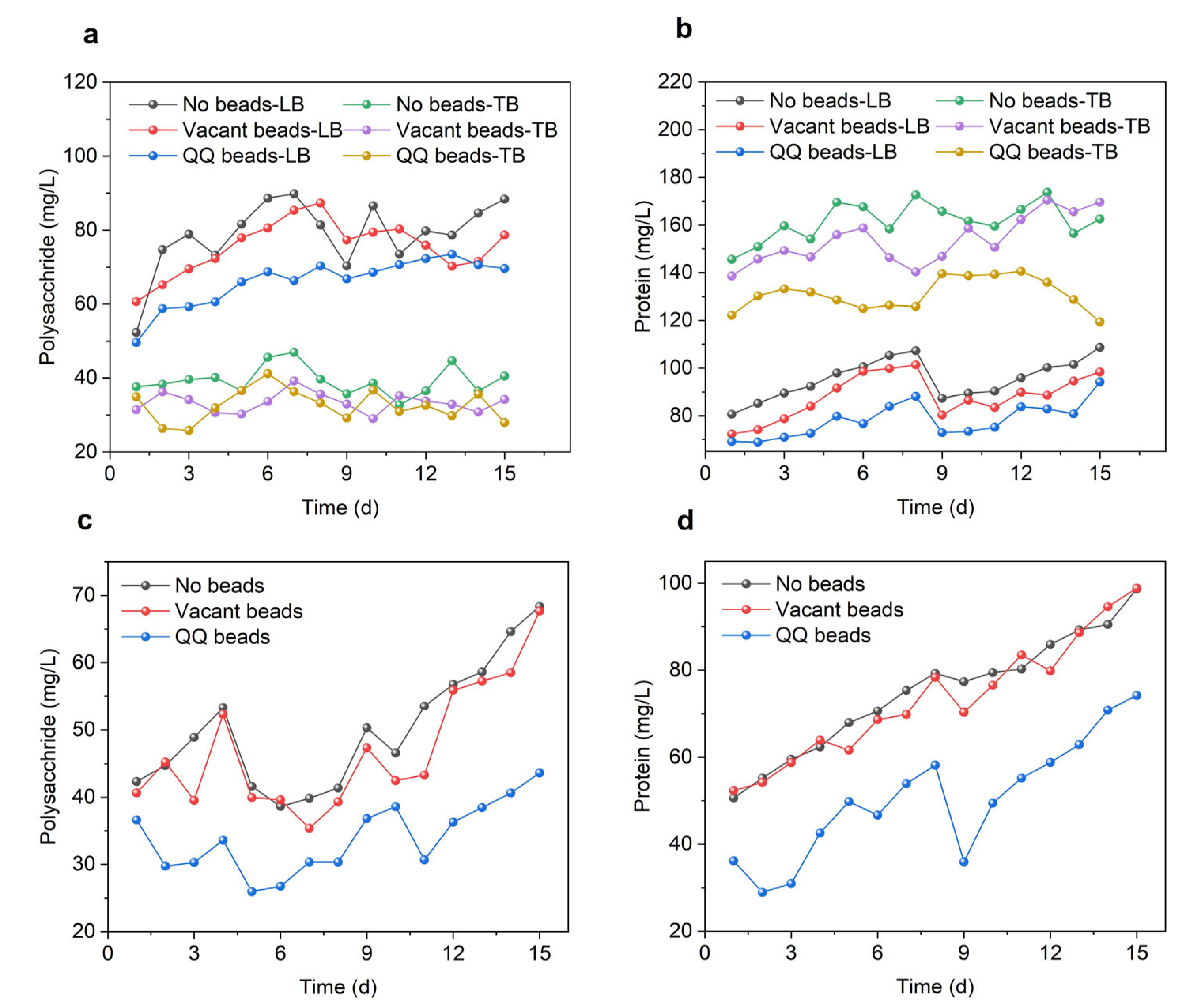
3.2. EPS and SMP Concentration in the Bioreactor Mixed Liquor
3.2.1. EPS
3.2.2. SMP
3.3. Effect of Quorum Quenching on Sludge Characteristics
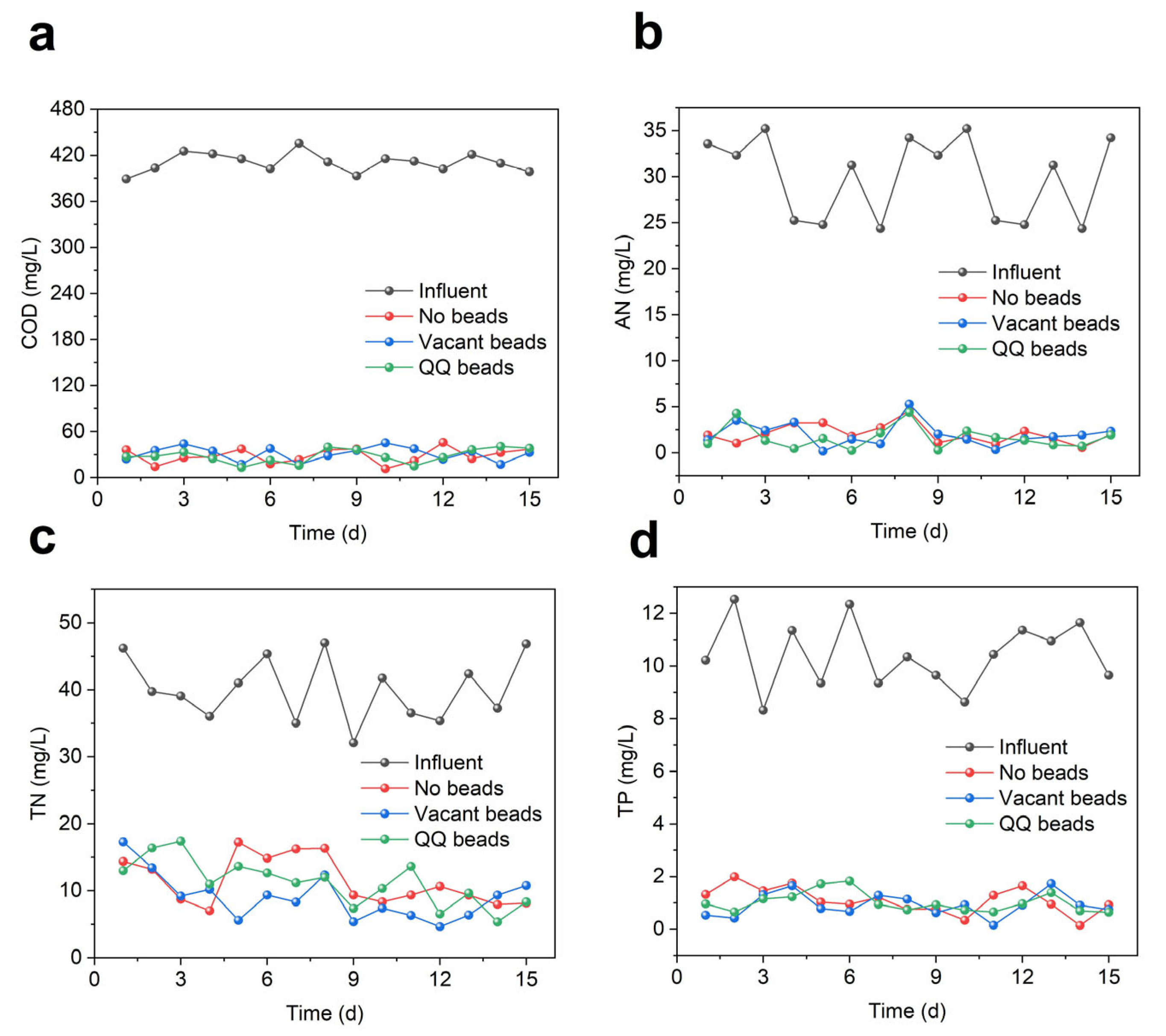
3.4. Effect of Quorum Quenching on MBR Treatment Efficiencies
3.5. Microbial Community Structure Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bijtenhoorn, P.; Mayerhofer, H.; Müller-Dieckmann, J.; Utpatel, C.; Schipper, C.; Hornung, C.; Szesny, M.; Grond, S.; Thürmer, A.; Brzuszkiewicz, E.; et al. A Novel Metagenomic Short-Chain Dehydrogenase/Reductase Attenuates Pseudomonas Aeruginosa Biofilm Formation and Virulence on Caenorhabditis Elegans. PLoS ONE 2011, 6, e26278. [Google Scholar] [CrossRef]
- Yamamoto, K.; Hiasa, M.; Mahmood, T.; Matsuo, T. Direct solid-liquid separation using hollow fiber membrane in an activated sludge aeration tank. Water Pollut. Res. Control. Brighton 1988, 21, 43–54. [Google Scholar] [CrossRef]
- Huang, L.; Lee, D.J. Membrane Bioreactor: A Mini Review on Recent R&D Works. Bioresour. Technol. 2015, 194, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xiao, K.; Shen, Y. Recent Advances in Membrane Bioreactor Technology for Wastewater Treatment in China. Front. Environ. Sci. Eng. China 2010, 4, 245–271. [Google Scholar] [CrossRef]
- Judd, S.J. The Status of Industrial and Municipal Effluent Treatment with Membrane Bioreactor Technology. Chem. Eng. J. 2016, 305, 37–45. [Google Scholar] [CrossRef]
- Krzeminski, P.; Leverette, L.; Malamis, S.; Katsou, E. Membrane Bioreactors—A Review on Recent Developments in Energy Reduction, Fouling Control, Novel Configurations, LCA and Market Prospects. J. Memb. Sci. 2017, 527, 207–227. [Google Scholar] [CrossRef]
- Meng, F.; Zhang, S.; Oh, Y.; Zhou, Z.; Shin, H.S.; Chae, S.R. Fouling in Membrane Bioreactors: An Updated Review. Water Res. 2017, 114, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Drews, A. Membrane Fouling in Membrane Bioreactors—Characterisation, Contradictions, Cause and Cures. J. Memb. Sci. 2010, 363, 1–28. [Google Scholar] [CrossRef]
- Young, T.; Muftugil, M.; Smoot, S.; Peeters, J. MBR vs. CAS: Capital and Operating Cost Evaluation. Water Pract. Technol. 2012, 7, wpt2012075. [Google Scholar] [CrossRef]
- Gao, T.; Xiao, K.; Zhang, J.; Zhang, X.; Wang, X.; Liang, S.; Sun, J.; Meng, F.; Huang, X. Cost-Benefit Analysis and Technical Efficiency Evaluation of Full-Scale Membrane Bioreactors for Wastewater Treatment Using Economic Approaches. J. Clean. Prod. 2021, 301, 126984. [Google Scholar] [CrossRef]
- Tang, K.; Xie, J.; Pan, Y.; Zou, X.; Sun, F.; Yu, Y.; Xu, R.; Jiang, W.; Chen, C. The Optimization and Regulation of Energy Consumption for MBR Process: A Critical Review. J. Environ. Chem. Eng. 2022, 10, 108406. [Google Scholar] [CrossRef]
- Gkotsis, P.K.; Batsari, E.L.; Peleka, E.N.; Tolkou, A.K.; Zouboulis, A.I. Fouling Control in a Lab-Scale MBR System: Comparison of Several Commercially Applied Coagulants. J. Environ. Manag. 2017, 203, 838–846. [Google Scholar] [CrossRef]
- Cui, Z.; Wang, J.; Zhang, H.; Ngo, H.H.; Jia, H.; Guo, W.; Gao, F.; Yang, G.; Kang, D. Investigation of Backwashing Effectiveness in Membrane Bioreactor (MBR) Based on Different Membrane Fouling Stages. Bioresour. Technol. 2018, 269, 355–362. [Google Scholar] [CrossRef]
- Wang, Z.; Meng, F.; He, X.; Zhou, Z.; Huang, L.N.; Liang, S. Optimisation and Performance of NaClO-Assisted Maintenance Cleaning for Fouling Control in Membrane Bioreactors. Water Res. 2014, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.K.; Kashif, A.; Fuwad, A.; Choi, Y. Current Advances in Treatment Technologies for Removal of Emerging Contaminants from Water–A Critical Review. Coord. Chem. Rev. 2021, 442, 213993. [Google Scholar] [CrossRef]
- Cui, Y.; Gao, H.; Yu, R.; Gao, L.; Zhan, M. Biological-Based Control Strategies for MBR Membrane Biofouling: A Review. Water Sci. Technol. 2021, 83, 2597–2614. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.G.; Yin, W.F.; Sam, C.K.; Koh, C.L. A Novel Medium for the Isolation of N-Acylhomoserine Lactone-Degrading Bacteria. J. Ind. Microbiol. Biotechnol. 2009, 36, 247. [Google Scholar] [CrossRef] [PubMed]
- Chong, T.M.; Koh, C.L.; Sam, C.K.; Choo, Y.M.; Yin, W.F.; Chan, K.G. Characterization of Quorum Sensing and Quorum Quenching Soil Bacteria Isolated from Malaysian Tropical Montane Forest. Sensors 2012, 12, 4846. [Google Scholar] [CrossRef]
- Davies, D.G.; Parsek, M.R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The Involvement of Cell-to-Cell Signals in the Development of a Bacterial Biofilm. Science 1998, 280, 295–298. [Google Scholar] [CrossRef]
- Fast, W.; Tipton, P.A. The Enzymes of Bacterial Census and Censorship. Trends Biochem. Sci. 2012, 37, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum Sensing and Quorum Quenching: The Yin and Yang of Bacterial Communication. ChemBioChem 2009, 10, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Bzdrenga, J.; Daudé, D.; Rémy, B.; Jacquet, P.; Plener, L.; Elias, M.; Chabrière, E. Biotechnological Applications of Quorum Quenching Enzymes. Chem. Biol. Interact. 2017, 267, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum Quenching: Role in Nature and Applied Developments. FEMS Microbiol. Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef] [PubMed]
- Maddela, N.R.; Sheng, B.; Yuan, S.; Zhou, Z.; Villamar-Torres, R.; Meng, F. Roles of Quorum Sensing in Biological Wastewater Treatment: A Critical Review. Chemosphere 2019, 221, 616–629. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, D.C.; Yeon, K.M.; Kim, S.R.; Lee, C.H. Enzyme-Immobilized Nanofiltration Membrane to Mitigate Biofouling Based on Quorum Quenching. Environ. Sci. Technol. 2011, 45, 1601–1607. [Google Scholar] [CrossRef]
- Oh, H.-S.; Yeon, K.-M.; Yang, C.-S.; Kim, S.-R.; Lee, C.-H.; Park, S.Y.; Han, J.Y.; Lee, J.-K. Control of Membrane Biofouling in MBR for Wastewater Treatment by Quorum Quenching Bacteria Encapsulated in Microporous Membrane. Environ. Sci. Technol. 2012, 46, 4877–4884. [Google Scholar] [CrossRef]
- Jahangir, D.; Oh, H.S.; Kim, S.R.; Park, P.K.; Lee, C.H.; Lee, J.K. Specific Location of Encapsulated Quorum Quenching Bacteria for Biofouling Control in an External Submerged Membrane Bioreactor. J. Memb. Sci. 2012, 411–412, 130–136. [Google Scholar] [CrossRef]
- Yeon, K.M.; Lee, C.H.; Kim, J. Magnetic Enzyme Carrier for Effective Biofouling Control in the Membrane Bioreactor Based on Enzymatic Quorum Quenching. Environ. Sci. Technol. 2009, 43, 7403–7409. [Google Scholar] [CrossRef]
- Lee, S.; Park, S.K.; Kwon, H.; Lee, S.H.; Lee, K.; Nahm, C.H.; Jo, S.J.; Oh, H.S.; Park, P.K.; Choo, K.H.; et al. Crossing the Border between Laboratory and Field: Bacterial Quorum Quenching for Anti-Biofouling Strategy in an MBR. Environ. Sci. Technol. 2016, 50, 1788–1795. [Google Scholar] [CrossRef]
- Yu, H.; Lee, K.; Zhang, X.; Choo, K.H. In Situ versus Pre-Quorum Quenching of Microbial Signaling for Enhanced Biofouling Control in Membrane Bioreactors. J. Memb. Sci. 2019, 592, 117387. [Google Scholar] [CrossRef]
- Kampouris, I.D.; Karayannakidis, P.D.; Banti, D.C.; Sakoula, D.; Konstantinidis, D.; Yiangou, M.; Samaras, P.E. Evaluation of a Novel Quorum Quenching Strain for MBR Biofouling Mitigation. Water Res. 2018, 143, 56–65. [Google Scholar] [CrossRef]
- Ham, S.Y.; Kim, H.S.; Cha, E.; Park, J.H.; Park, H.D. Mitigation of Membrane Biofouling by a Quorum Quenching Bacterium for Membrane Bioreactors. Bioresour. Technol. 2018, 258, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, X.; Shi, H.; Wang, W.; Jiang, C.; Xiao, Z.; Xu, Y.; Ren, N. New Insights into Biofilm Control and Inhibitory Mechanism Analysis Based on the Novel Quorum Quenching Bacterium Acinetobacter Pittii HITSZ001. J. Memb. Sci. 2022, 663, 121012. [Google Scholar] [CrossRef]
- Kim, S.R.; Oh, H.S.; Jo, S.J.; Yeon, K.M.; Lee, C.H.; Lim, D.J.; Lee, C.H.; Lee, J.K. Biofouling Control with Bead-Entrapped Quorum Quenching Bacteria in Membrane Bioreactors: Physical and Biological Effects. Environ. Sci. Technol. 2013, 47, 836–842. [Google Scholar] [CrossRef]
- Feng, X.C.; Guo, W.Q.; Zheng, H.S.; Wu, Q.L.; Luo, H.C.; Ren, N.Q. Effect of Metabolic Uncoupler, 3,3′,4′,5-Tetrachlorosalicylanilide (TCS) on Bacillus Subtilis: Biofilm Formation, Flocculability and Surface Characteristics. RSC Adv. 2018, 8, 16178–16186. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhang, C.; Lian, J.; Lu, C.; Chen, Z.; Song, Y.; Guo, Y.; Xing, Y. Effect of Thiosulfate on Rapid Start-up of Sulfur-Based Reduction of High Concentrated Perchlorate: A Study of Kinetics, Extracellular Polymeric Substances (EPS) and Bacterial Community Structure. Bioresour. Technol. 2017, 243, 932–940. [Google Scholar] [CrossRef]
- Somani, B.L.; Khanade, J.; Sinha, R. A Modified Anthrone-Sulfuric Acid Method for the Determination of Fructose in the Presence of Certain Proteins. Anal. Biochem. 1987, 167, 327–330. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, Z.; Tang, S. Extracellular Polymeric Substances (EPS) Properties and Their Effects on Membrane Fouling in a Submerged Membrane Bioreactor. Water Res. 2009, 43, 2504–2512. [Google Scholar] [CrossRef]
- Nwachukwu, I.D.; Girgih, A.T.; Malomo, S.A.; Onuh, J.O.; Aluko, R.E. Thermoase-Derived Flaxseed Protein Hydrolysates and Membrane Ultrafiltration Peptide Fractions Have Systolic Blood Pressure-Lowering Effects in Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2014, 15, 18131–18147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, F.; Sun, S.; Jing, Y.; Wang, F.; Wei, Q.; Wang, X.; Zhu, X. A Rapid and Sensitive Method for Determination of Trace Amounts of Glucose by Anthrone-Sulfuric Acid Method. Bulg. Chem. Commun. 2016, 48, 109–113. [Google Scholar]
- Rice, E.W.; Baird, R.B.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
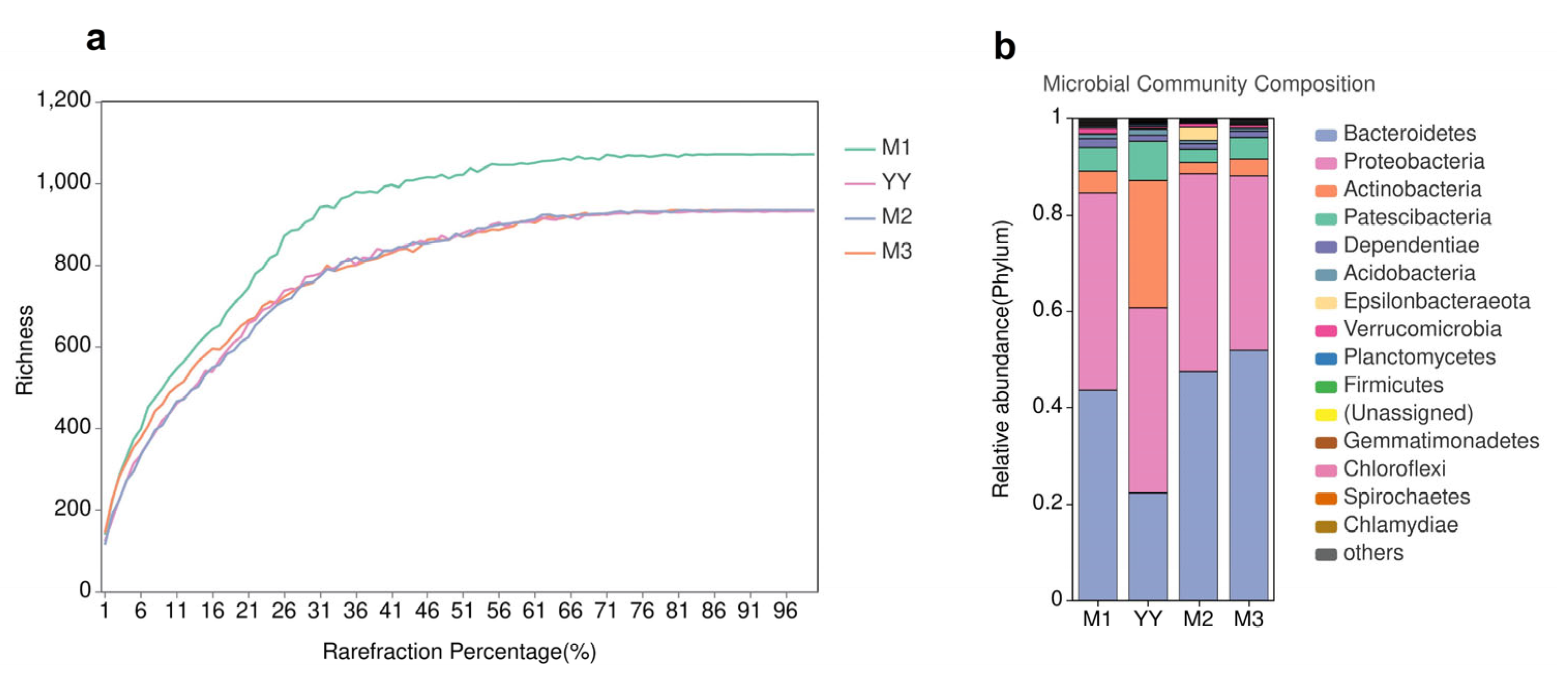{kind=link}
| Samples | Sequence Number | OTU | Chao1 | Shannon | Simpson |
|---|---|---|---|---|---|
| YY | 60,159 | 1029 | 1029.4 | 4.59 | 0.0376 |
| M1 | 61,418 | 1204 | 1204.2 | 4.22 | 0.05 |
| M2 | 59,082 | 1097 | 1097.4 | 4.16 | 0.069 |
| M3 | 71,561 | 1084 | 1084.6 | 4.18 | 0.0773 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Feng, X.; Wang, W.; Shi, H.; Xiao, Z.; Jiang, C.; Xu, Y.; Zhang, X.; Ren, N. Application of Encapsulated Quorum Quenching Strain Acinetobacter pittii HITSZ001 to a Membrane Bioreactor for Biofouling Control. Separations 2023, 10, 127. https://doi.org/10.3390/separations10020127
Wang Y, Feng X, Wang W, Shi H, Xiao Z, Jiang C, Xu Y, Zhang X, Ren N. Application of Encapsulated Quorum Quenching Strain Acinetobacter pittii HITSZ001 to a Membrane Bioreactor for Biofouling Control. Separations. 2023; 10(2):127. https://doi.org/10.3390/separations10020127
Chicago/Turabian StyleWang, Yongmei, Xiaochi Feng, Wenqian Wang, Hongtao Shi, Zijie Xiao, Chenyi Jiang, Yujie Xu, Xin Zhang, and Nanqi Ren. 2023. "Application of Encapsulated Quorum Quenching Strain Acinetobacter pittii HITSZ001 to a Membrane Bioreactor for Biofouling Control" Separations 10, no. 2: 127. https://doi.org/10.3390/separations10020127