
Production of Phenyllactic Acid from Porphyra Residues by Lactic Acid Bacterial Fermentation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Porphyra Residues and Untrafiltration Elute
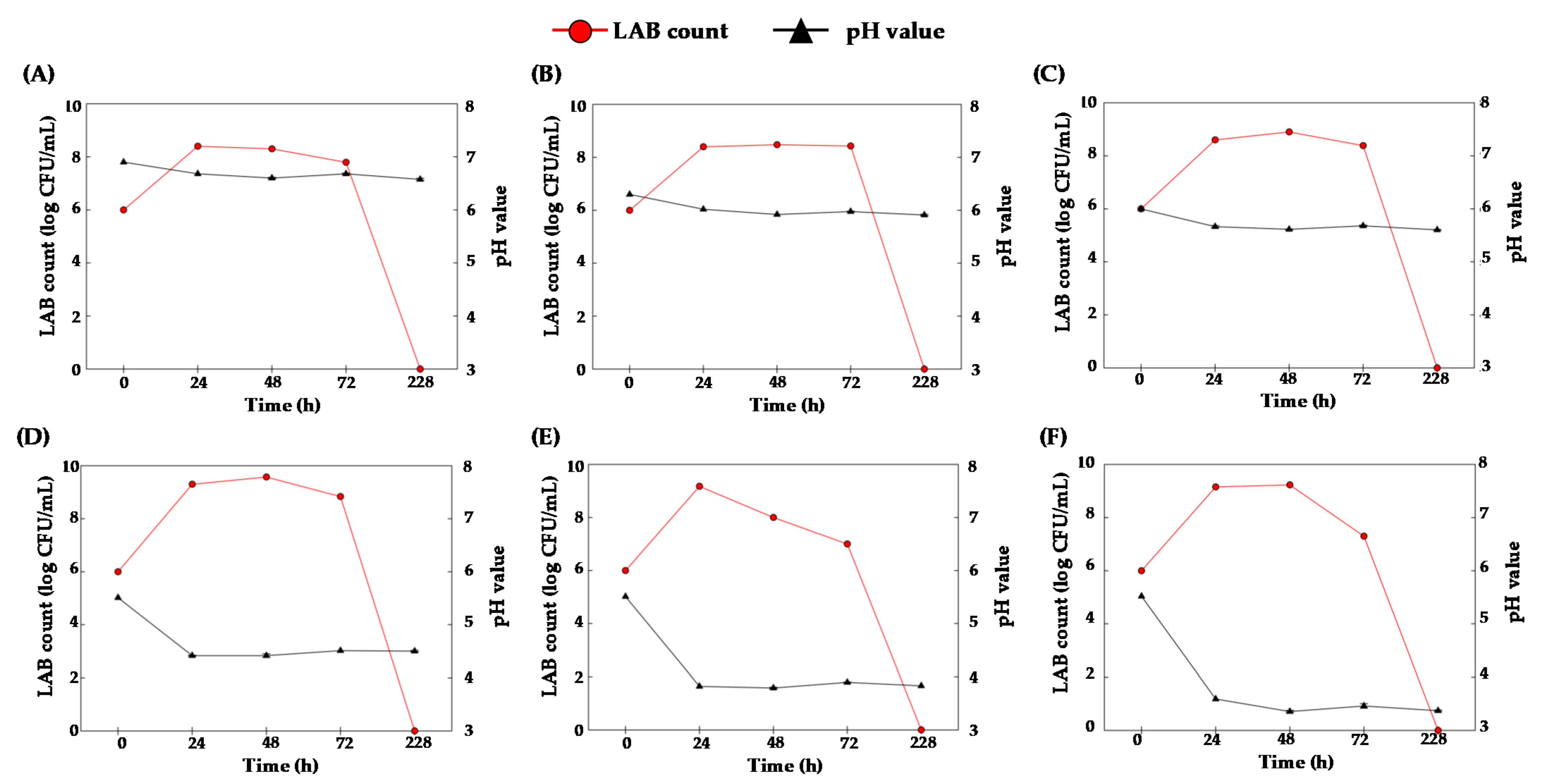
2.3. Screening the Strains of LAB for PhLA Production
2.4. Fermentation of Porphyra Residues by LAB
2.5. Determination of Total Sugar and Reducing Sugar Contents
2.6. Extraction and Determination of PhLA, Lactic Acid, and Phe
2.7. Statistical Analysis
3. Results and Discussion
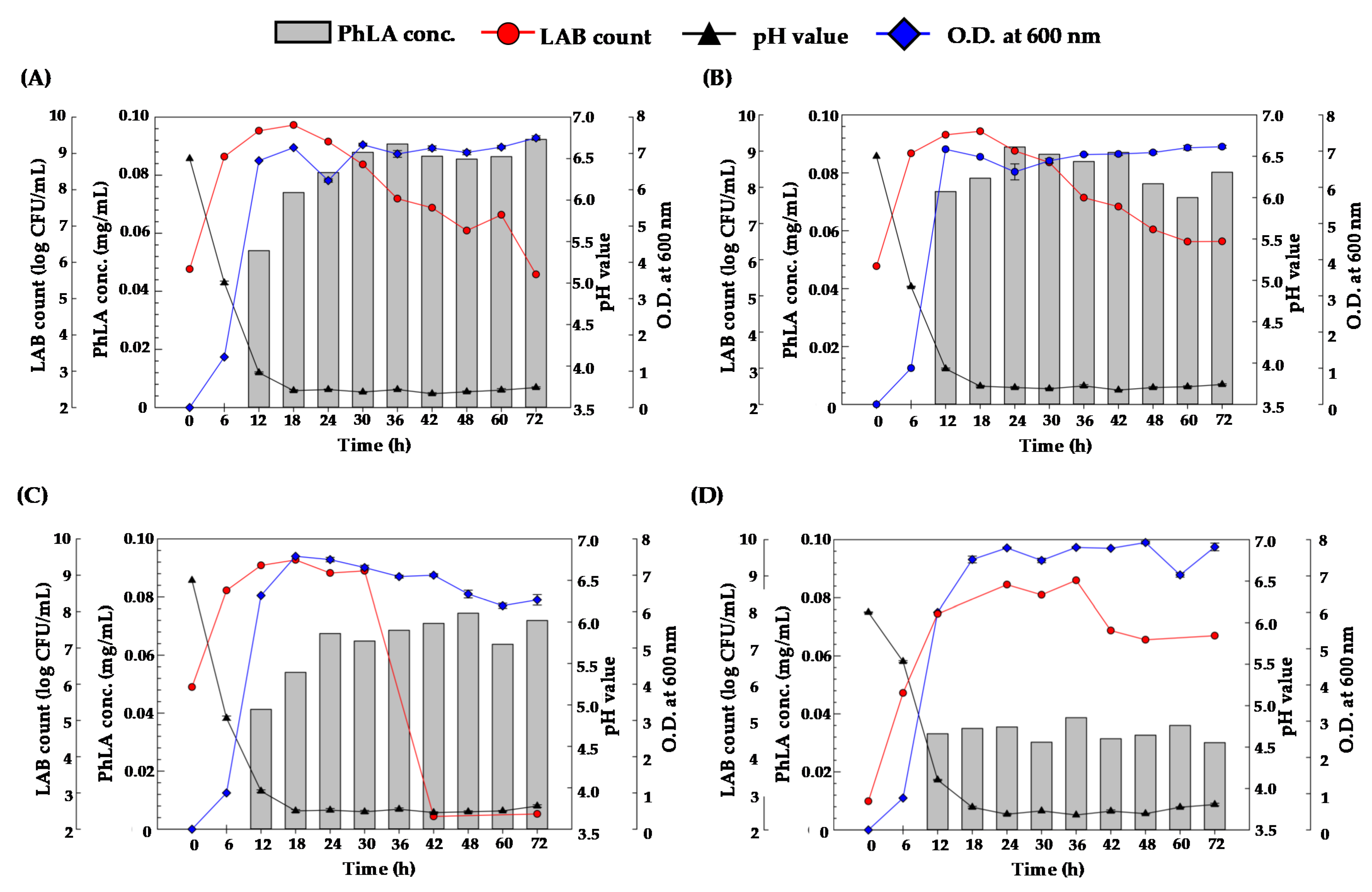
3.1. The Feasibility of PhLA Production from Porphyra Residues by Fermentation Process
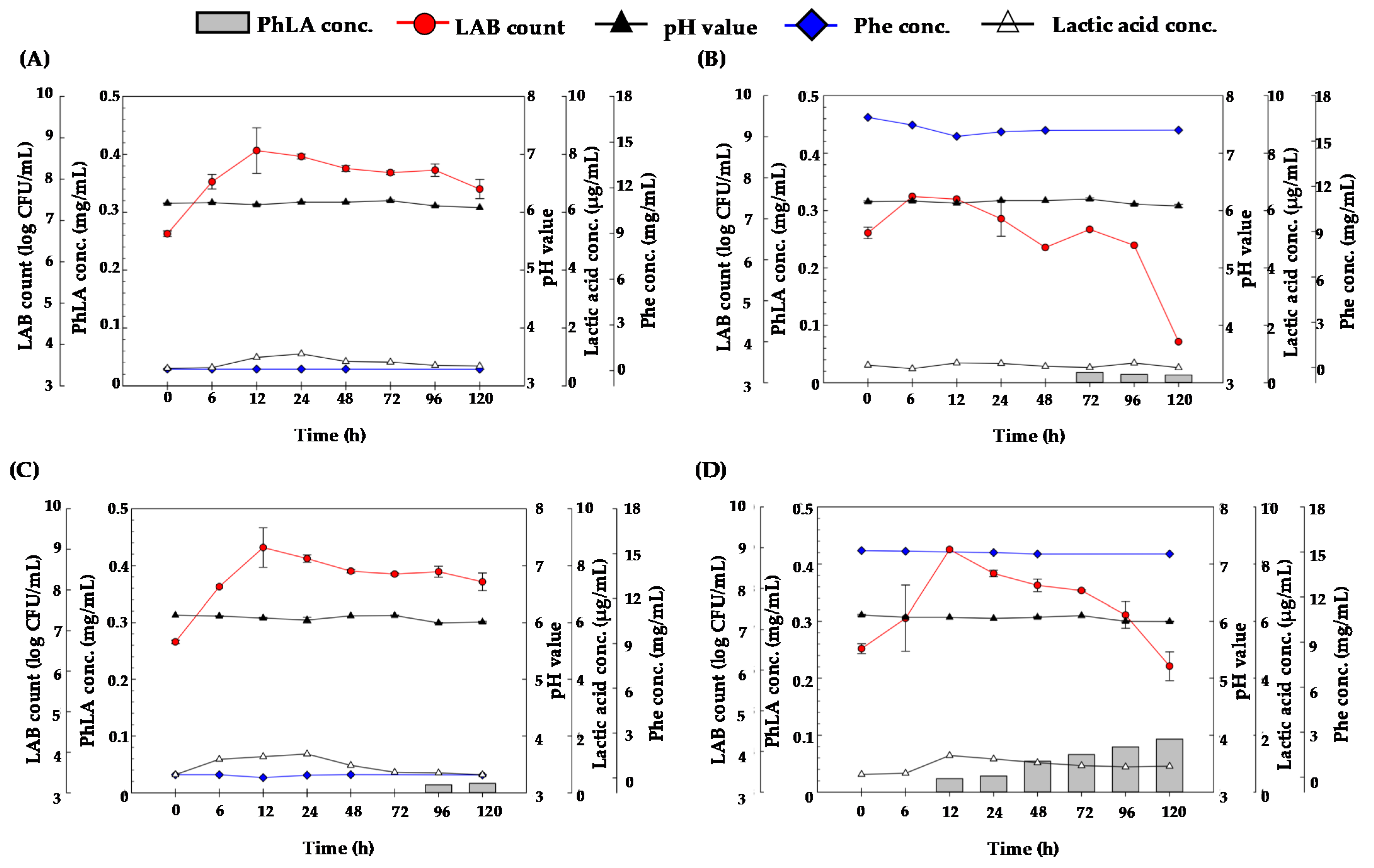
3.2. PhLA Production from Porphyra Residues by Fermentation with KP3 and KP4 Strains
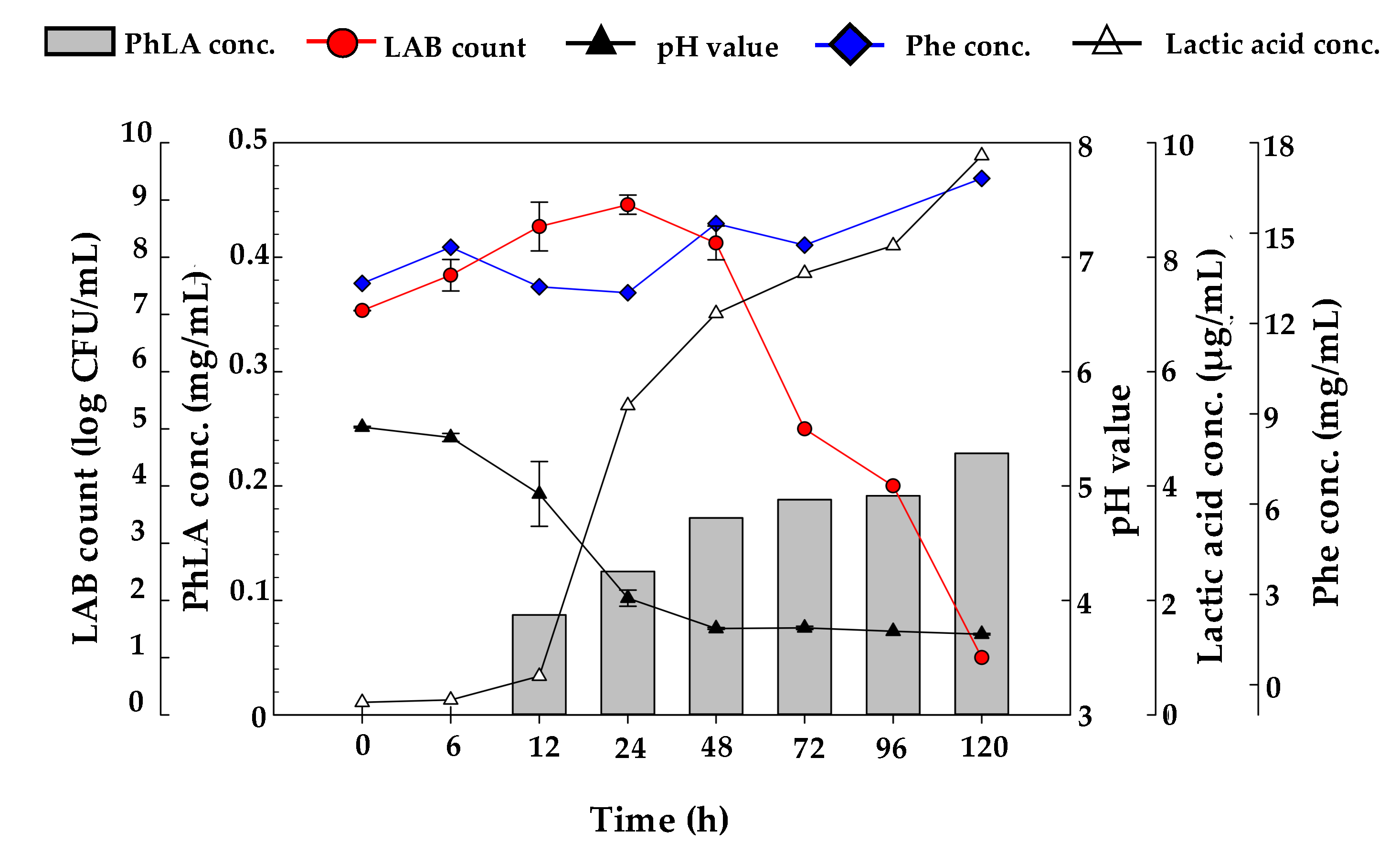
3.3. Pretreatment of Porphyra Residues with Cellulase for PhLA Production by LAB Fermentation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khoo, C.G.; Dasan, Y.K.; Lam, M.K.; Lee, K.T. Algae biorefinery: Review on a broad spectrum of downstream processes and products. Bioresour. Technol. 2019, 292, 121964. [Google Scholar] [CrossRef] [PubMed]
- Kuda, T.; Eda, M.; Kataoka, M.; Nemoto, M.; Kawahara, M.; Oshio, S.; Takahashi, H.; Kimura, B. Anti-glycation properties of the aqueous extract solutions of dried algae products and effect of lactic acid fermentation on the properties. Food Chem. 2016, 192, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, M.; Jegatheesan, A.; Poonam, C.; Perumal, K.; Arunkumar, K. Biosaccharification and ethanol production from spent seaweed biomass using marine bacteria and yeast. Renew. Energy 2017, 105, 133–139. [Google Scholar] [CrossRef]
- Ho, S.H.; Huang, S.W.; Chen, C.Y.; Hasunuma, T.; Kondo, A.; Chang, J.S. Bioethanol production using carbohydrate-rich microalgae biomass as feedstock. Bioresour. Technol. 2013, 135, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Günerken, E.; D’Hondt, E.; Eppink, M.; Garcia-Gonzalez, L.; Elst, K.; Wijffels, R.H. Cell disruption for microalgae biorefineries. Biotechnol. Adv. 2015, 33, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Chen, W.C.; Gao, Y.H.; Chen, G.W.; Lin, H.T.; Pan, C.L. Enzyme-assisted method for phycobiliproteins extraction from Porphyra and evaluation of their bioactivity. Processes 2021, 9, 560. [Google Scholar] [CrossRef]
- Guan, J.; Han, C.; Guan, Y.; Zhang, S.; Yun, J.; Yao, S. Optimizational production of phenyllactic acid by a Lactobacillus buchneri strain via uniform design with overlay sampling methodology. Chin. J. Chem. Eng. 2019, 27, 418–425. [Google Scholar] [CrossRef]
- Prema, P.; Smila, D.; Palavesam, A.; Immanuel, G. Production and characterization of an antifungal compound (3-phenyllactic acid) produced by Lactobacillus plantarum strain. Food Bioprocess Technol. 2010, 3, 379–386. [Google Scholar] [CrossRef]
- Ström, K.; Schnürer, J.; Melin, P. Co-cultivation of antifungal Lactobacillus plantarum MiLAB 393 and Aspergillus nidulans, evaluation of effects on fungal growth and protein expression. FEMS Microbiol. Lett. 2005, 246, 119–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, T.; Nguyen, H.D.; Ito, T.; Zhou, S.; Osada, L.; Tateyama, S.; Kaneko, T.; Takaya, N. Microbial monomers custom-synthesized to build true bio-derived aromatic polymers. Appl. Microbiol. Biotechnol. 2013, 97, 8887–8894. [Google Scholar] [CrossRef]
- Tiwari, B.K.; Troy, D.J. Seaweed sustainability–food and nonfood applications. In Seaweed Sustainability; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1–6. [Google Scholar]
- Lin, H.T.; Huang, M.Y.; Kao, T.Y.; Lu, W.J.; Lin, H.J.; Pan, C.L. Production of lactic acid from seaweed hydrolysates via lactic acid bacteria fermentation. Fermentation 2020, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Cortés-Zavaleta, O.; López-Malo, A.; Hernández-Mendoza, A.; García, H. Antifungal activity of lactobacilli and its relationship with 3-phenyllactic acid production. Int. J. Food Microbiol. 2014, 173, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, B.; Pan, B. Biotransformation of phenylpyruvic acid to phenyllactic acid by growing and resting cells of a Lactobacillus sp. Biotechnol. Lett. 2007, 29, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Bustos, A.Y.; de Valdez, G.F.; Gerez, C.L. Optimization of phenyllactic acid production by Pediococcus acidilactici CRL 1753. Application of the formulated bio-preserver culture in bread. Biol. Control 2018, 123, 137–143. [Google Scholar] [CrossRef]
- Valerio, F.; Lavermicocca, P.; Pascale, M.; Visconti, A. Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol. Lett. 2004, 233, 289–295. [Google Scholar] [CrossRef]
- Zhao, W.; Ding, H.; Lv, C.; Hu, S.; Huang, J.; Zheng, X.; Yao, S.; Mei, L. Two-step biocatalytic reaction using recombinant Escherichia coli cells for efficient production of phenyllactic acid from l-phenylalanine. Process Biochem. 2018, 64, 31–37. [Google Scholar] [CrossRef]
- Wu, S.C.; Wang, F.J.; Pan, C.L. The comparison of antioxidative properties of seaweed oligosaccharides fermented by two lactic acid bacteria. J. Mar. Sci. Technol. 2010, 18, 537–545. [Google Scholar]
- Kawaguchi, H.; Teramura, H.; Uematsu, H.; Hara, J.Y.; Hasunuma, T.; Hirano, K.; Sazuka, T.; Kitano, H.; Tsuge, Y.; Kahar, P.; et al. Phenyllactic acid production by simultaneous saccharification and fermentation of pretreated sorghum bagasse. Bioresour. Technol. 2015, 182, 169–178. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Sugar (mg/mL) | Reducing Sugar (mg/mL) | Lactic Acid (%) | PhLA(%) |
|---|---|---|---|---|
| Porphyra enzymatic hydrolysates 1 | 14.22 ± 1.27 | 1.12 ± 0.11 | - | - |
| Ultrafiltration (UF) > 100 kDa 2 | 14.02 ± 1.38 | 1.10 ± 0.10 | - | - |
| UF < 100 kDa 3 | 11.87 ± 1.20 | 0.86 ± 0.10 | - | - |
| Unfermented Porphyra residues 4 | 4.80 ± 0.19 | 0.42 ± 0.06 | - | - |
| KP4 fermented Porphyra 5 | 0.06 ± 0.00 | 0.04 ± 0.01 | 0.13% | N.D. 7 |
| KP3 fermented Porphyra 6 | 0.05 ± 0.00 | 0.03 ± 0.00 | 0.13% | N.D. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-H.; Chen, W.-C.; Gao, Y.-H.; Hsiao, H.-I.; Pan, C.-L. Production of Phenyllactic Acid from Porphyra Residues by Lactic Acid Bacterial Fermentation. Processes 2021, 9, 678. https://doi.org/10.3390/pr9040678
Huang C-H, Chen W-C, Gao Y-H, Hsiao H-I, Pan C-L. Production of Phenyllactic Acid from Porphyra Residues by Lactic Acid Bacterial Fermentation. Processes. 2021; 9(4):678. https://doi.org/10.3390/pr9040678
Chicago/Turabian StyleHuang, Chung-Hsiung, Wei-Chen Chen, Yu-Huei Gao, Hsin-I Hsiao, and Chorng-Liang Pan. 2021. "Production of Phenyllactic Acid from Porphyra Residues by Lactic Acid Bacterial Fermentation" Processes 9, no. 4: 678. https://doi.org/10.3390/pr9040678