
Improving the Thermo-Activity and -Stability of Pectate Lyase from Dickeya dadantii DCE-01 for Ramie Degumming


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Point Mutations Prediction
2.3. Mutation Sites and Primer
2.4. Construction and Expression of Mutant Recombinant Plasmid
2.5. Pectate Lyase Activity Determination and SDS-PAGE Analysis
2.6. Screening of Heat Stable Mutants
2.7. Optimal Ca2+ Concentration of Mutant
2.8. Enzymatic Degumming of Ramie
3. Results and Discussion
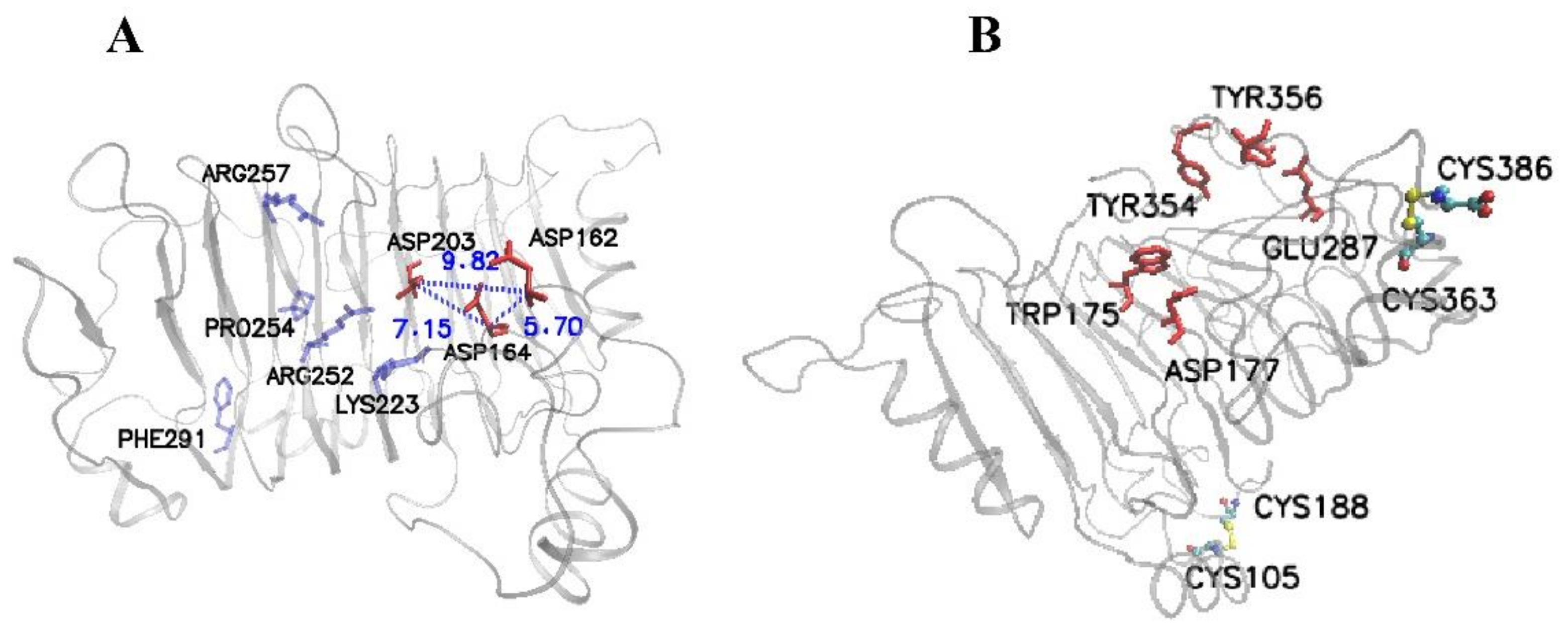
3.1. Determination of the Potential Mutant Sites
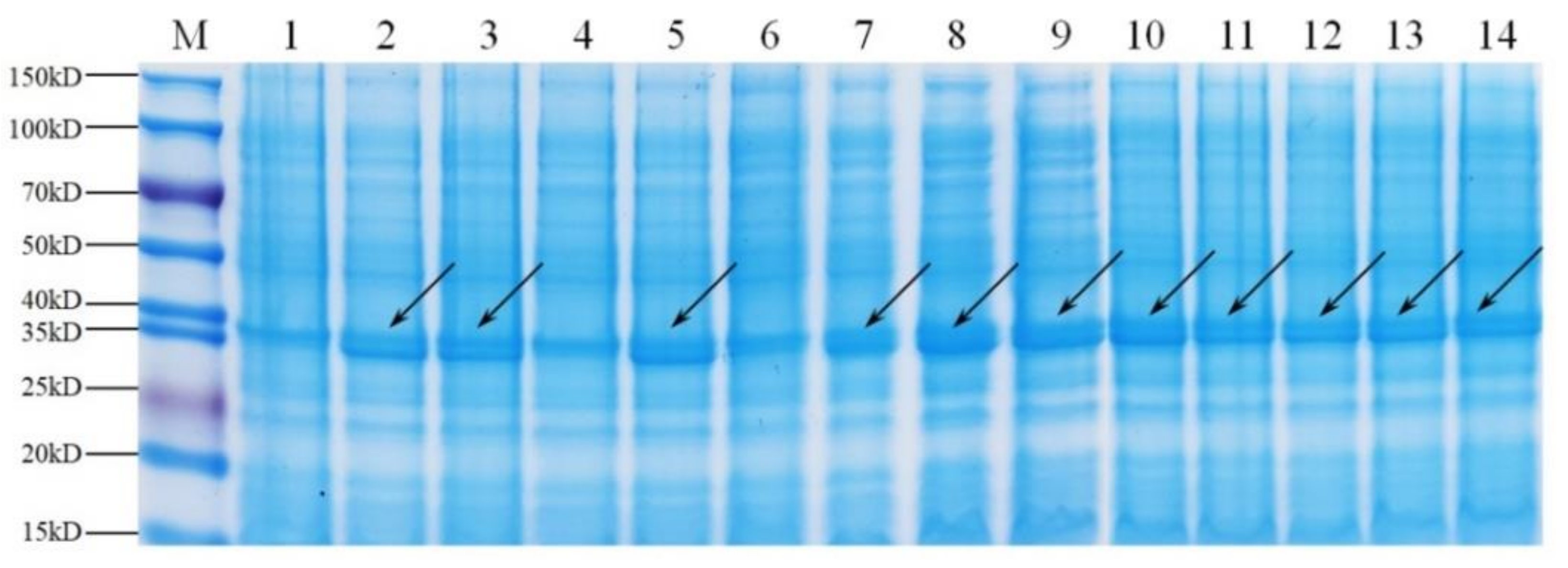
3.2. Analysis of Exogenous Expression of Mutant Pels
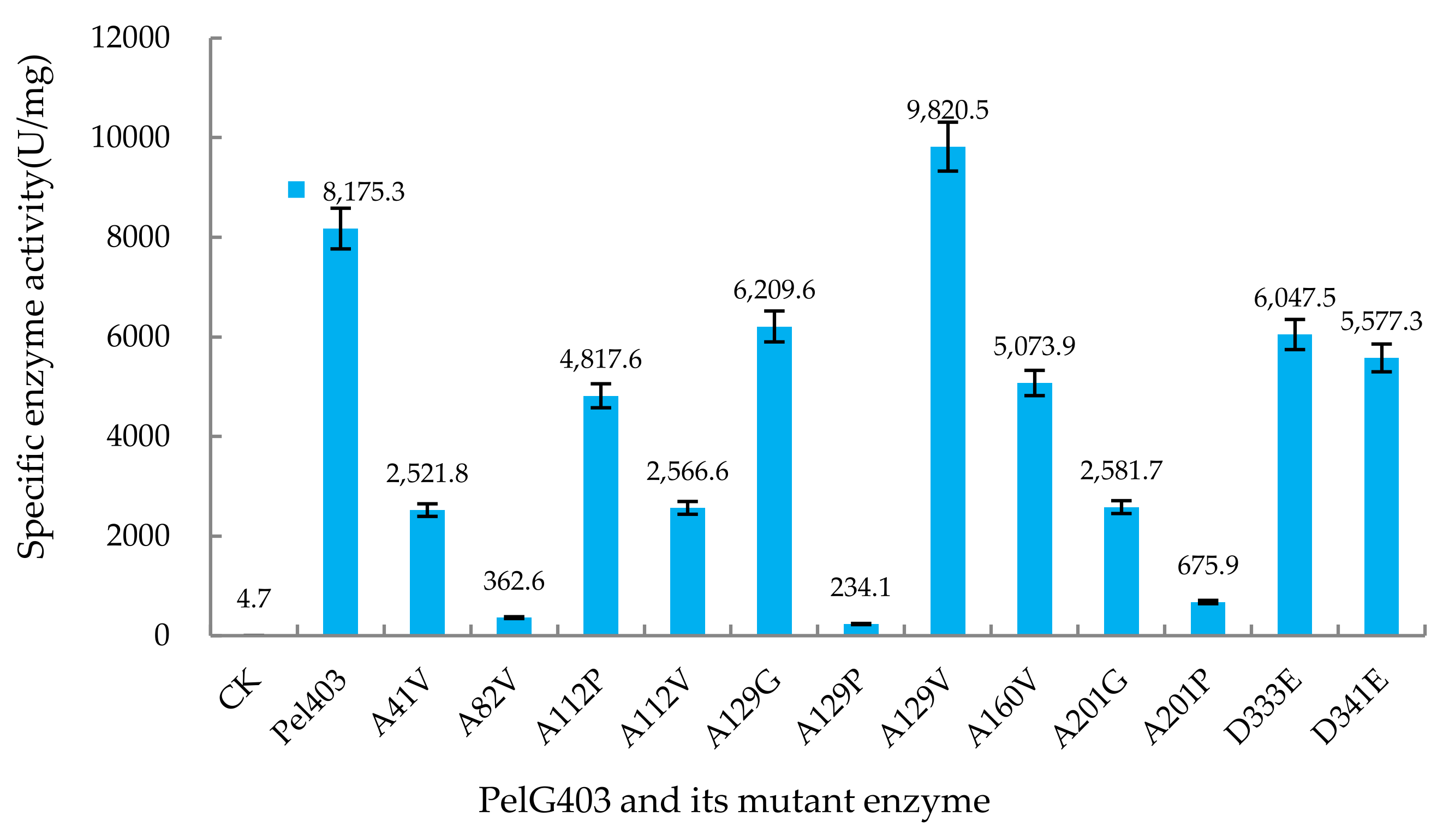
3.3. Analysis of Pectate Lyase Activity
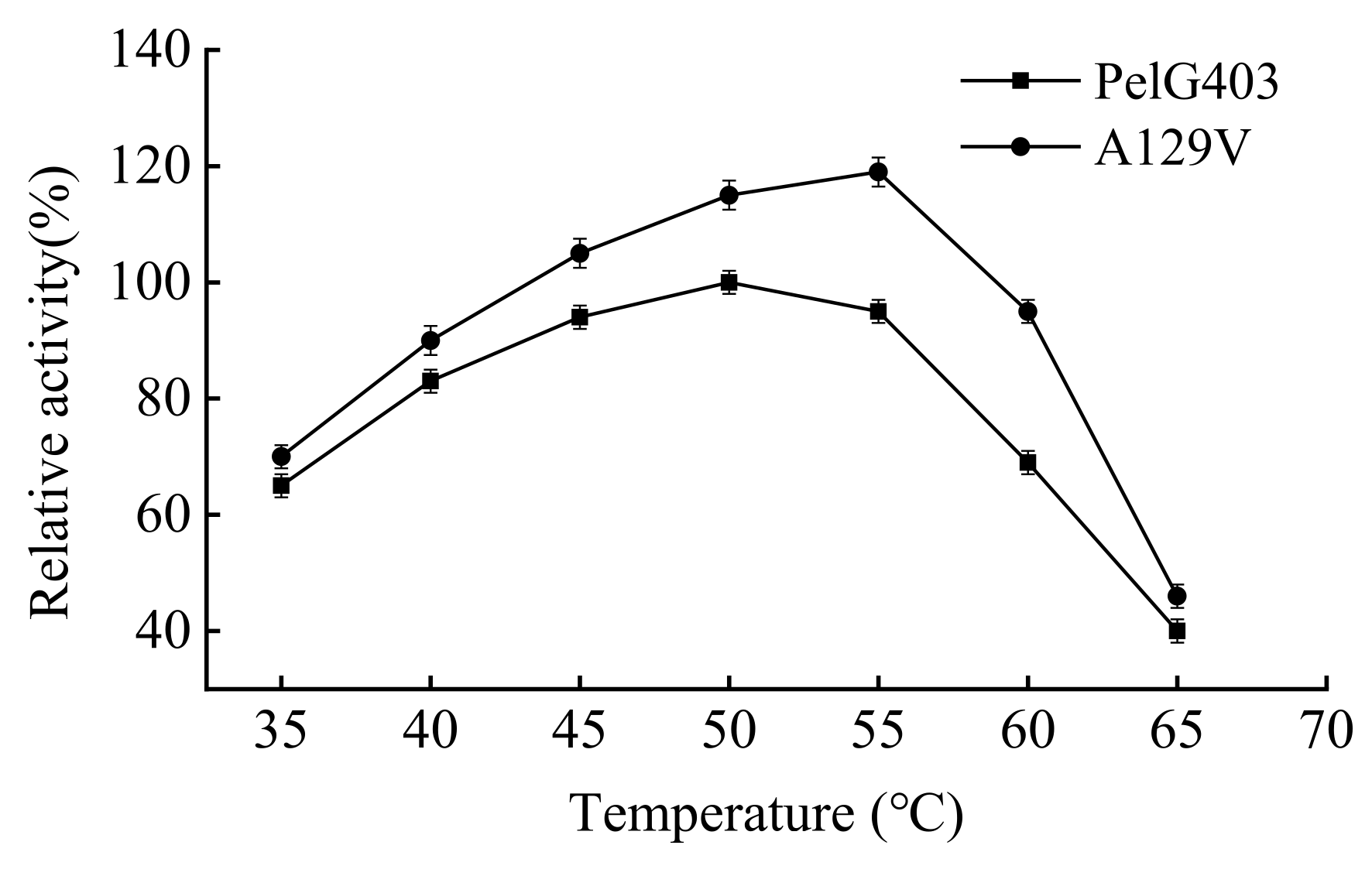
3.4. Optimal Reaction Temperature
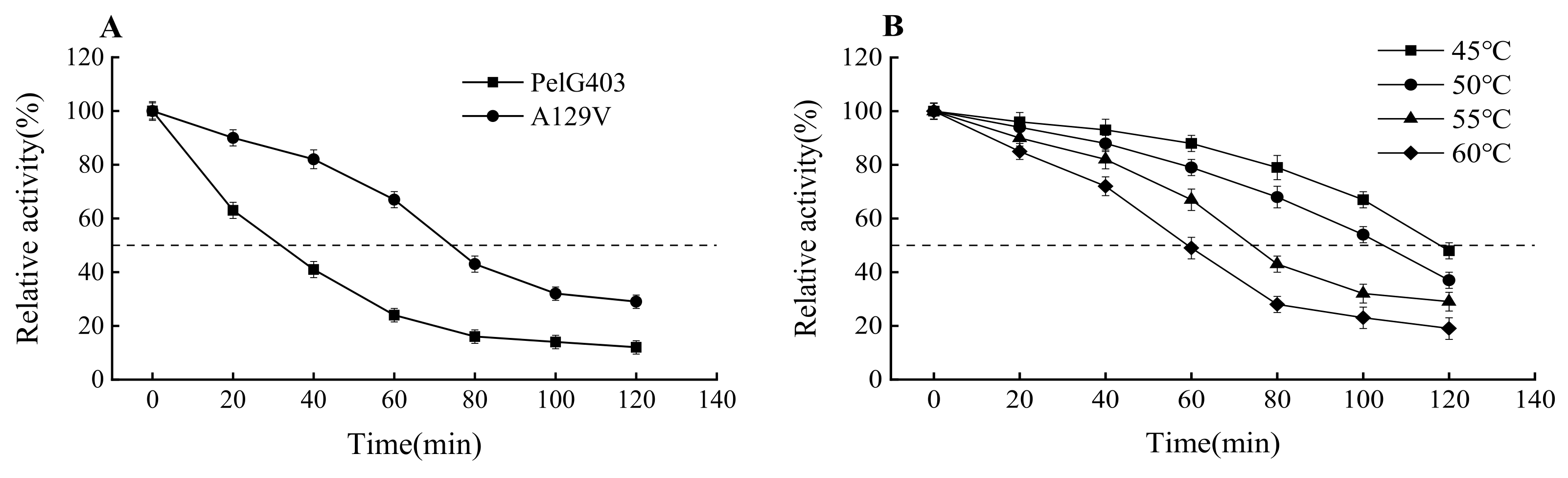
3.5. Thermal Stability
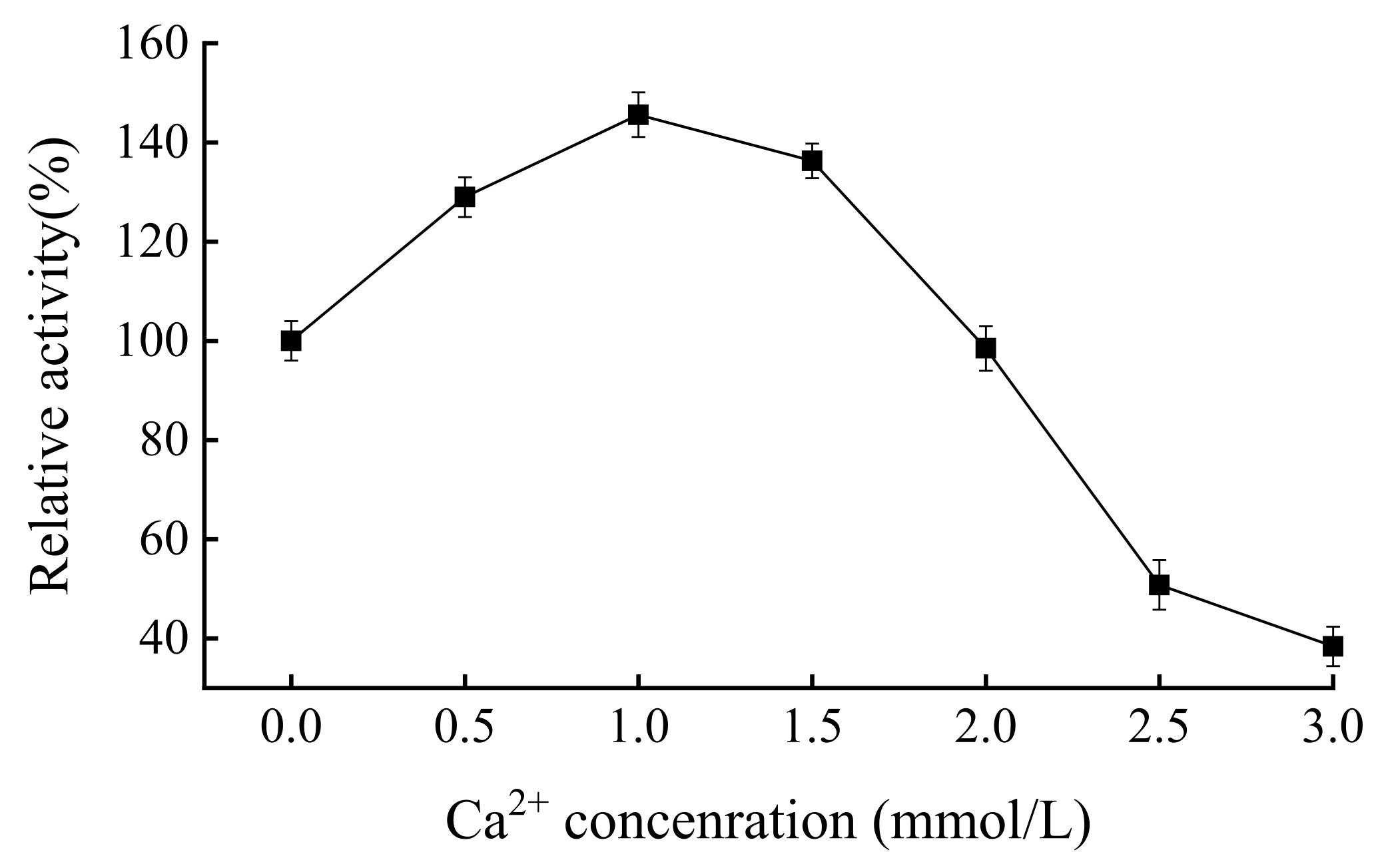
3.6. Analysis of the Optimal Ca2+ Concentration of Forward Mutant Enzyme
3.7. Ramie Degumming
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hugouvieux-Cotte-Pattat, N.; Condemine, G.; Shevchik, V.E. Bacterial pectate lyases, structural and functional diversity. Environ. Microbiol. Rep. 2014, 6, 427–440. [Google Scholar] [CrossRef]
- Tardy, F.; Nasser, W.; Robert-Baudouy, J.; Hugouvieux-Cotte-Pattat, N. Comparative analysis of the five major Erwinia chrysanthemi pectate lyases: Enzyme characteristics and potential inhibitors. J. Bacteriol. 1997, 179, 2503–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitri; Tawali, A.; Tawali, A.B.; Laga, A. The effect of soaking time on mucilage removal from the coffee bean using pectinase enzyme. IOP Conf. Ser. Earth Environ. Sci. 2021, 807, 022053. [Google Scholar] [CrossRef]
- Sonia, A.; Mandhan, R.P.; Sudha, D.S.; Rakesh, K.; Jitender, S. Potential application of alkaline pectinase from Bacillus subtilis SS in pulp and paper industry. Appl. Biochem. Biotechnol. 2008, 149, 287–293. [Google Scholar]
- Nwobi, A.; Cybulska, I.; Tesfai, W.; Shatilla, Y.; Rodríguez, J.; Thomsen, M.H. Simultaneous saccharification and fermentation of solid household waste following mild pretreatment using a mix of hydrolytic enzymes in combination with Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2015, 99, 929–938. [Google Scholar] [CrossRef]
- Neelima, M.; Kavita, S.; Mukty, S.; Archana, D.; Brajesh, P.; Hyeji, J.; Seorin, P.; Srinath, P.; Sunghun, C. Biotransformation of Citrus Waste-I: Production of Biofuel and Valuable Compounds by Fermentation. Processes 2021, 9, 220. [Google Scholar]
- Aggarwal, R.; Dutta, T.; Sheikh, J. Extraction of pectinase from Candida isolated from textile mill effluent and its application in bio-scouring of cotton. Sustain. Chem. Pharm. 2020, 17, 100291. [Google Scholar] [CrossRef]
- Li, Z.F.; Li, Z.L.; Ding, R.Y.; Yu, C.W. Composition of ramie hemicelluloses and effect of polysaccharides on fiber properties. Text. Res. J. 2016, 86, 451–460. [Google Scholar] [CrossRef]
- Fan, P.; He, F.; Yang, Y.; Ao, M.; Ouyang, J.; Liu, Y.; Yu, L. In-situ microbial degumming technology with Bacillus sp. HG-28 for industrial production of ramie fibers. Biochem. Eng. J. 2015, 97, 50–58. [Google Scholar] [CrossRef]
- Vardhini, V.K.J.; Murugan, R.; Surjit, R. Effect of alkali and enzymatic treatments of banana fibre on properties of banana/polypropylene composites. J. Ind. Text. 2018, 47, 1849–1864. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Dutta, N.; Chattopadhyay, D.; Chakrabarti, K. Degumming of ramie fiber and the production of reducing sugars from waste peels using nanoparticle supplemented pectate lyase. Bioresour. Technol. 2013, 137, 202–208. [Google Scholar] [CrossRef]
- Yang, Q.; Duan, S.; Cheng, L.; Feng, X.; Zheng, K.; Xie, C.; Liu, Z.; Pen, Y. Engineering of a Bacillus subtilis Strain Deficient in Cellulase: Application in Degumming of Ramie. FIBERS Polym. 2019, 20, 57–62. [Google Scholar] [CrossRef]
- Guo, F.; Zou, M.; Li, X.; Zhao, J.; Qu, Y.; Abd-Aziz, S. An effective degumming enzyme from Bacillus sp. Y1 and synergistic action of hydrogen peroxide and protease on enzymatic degumming of ramie fibers. BioMed Res. Int. 2013, 2013, 212315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, H.; Zhou, C.; Ma, Y.; Li, J.; Song, J. The alkaline pectate lyase PEL168 of Bacillus subtilis heterologously expressed in Pichia pastorisis more stable and efficient for degumming ramie fiber. BMC Biotechnol. 2013, 13, 26–35. [Google Scholar]
- Yadav, S.; Dubey, A.K.; Anand, G.; Kumar, R.; Yadav, D. Purification and biochemical characterization of an alkaline pectin lyase from Fusarium decemcellulareMTCC 2079 suitable for Crotolaria juncea fiber retting. J. Basic Microbiol. 2014, 54, 161–169. [Google Scholar] [CrossRef]
- Zhou, C.; Xue, Y.; Ma, Y. Cloning, evaluation, and high-level expression of a thermo-alkaline pectatelyase from alkaliphilic Bacillus clausii with potential in ramie degumming. Appl. Microbiol. Biotechnol. 2017, 101, 3663–3676. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Duan, S.; Zheng, K.; Feng, X.; Yang, Q.; Liu, Z.Y.; Liu, Z.; Peng, Y. An alkaline pectate lyase D from Dickeya dadantii DCE-01: Clone, expression, characterization, and potential application in ramie bio-degumming. Text. Res. J. 2019, 89, 2075–2083. [Google Scholar] [CrossRef]
- Shu, T.; Bai, Y.; Wang, Y.; Wang, H.; Li, P.; Xiang, M.; Yu, T.; Xu, H.; Yu, L. A high-efficiency and eco-friendly degumming process for ramie fibers. J. Clean. Prod. 2020, 276, 124271. [Google Scholar] [CrossRef]
- Wang, Y.; Bai, Y.; Shu, T.; Fan, P.; Zhang, H.; Turunen, O.; Xiong, H.; Yu, L. Characterization of a versatile glycoside hydrolase Cel5M from Pectobacterium carotovorum HG-49 for ramie degumming. Text. Res. J. 2020, 90, 1602–1615. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Zhou, C.; Ma, Y.; Li, L.; Song, J. Cloning, expression and characterization of a pectate lyase from Paenibacillus sp. 0602 in recombinant Escherichia coli. BMC Biotechnol. 2014, 14, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.; Li, X.; Zhao, J.; Qu, Y. Characteristics of polygalacturonate lyase C from Bacillus subtilis 7-3-3 and its synergistic action with pelA in enzymatic degumming. PLoS ONE 2013, 8, e79357. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Duan, S.; Feng, X.; Zheng, K.; Yang, Q.; Wang, Q.; Cheng, L.; Peng, Y. High level expression of pectate lyase gene from microbiology for ramie degumming and its bioinformatics analysis. Acta Agric. Boreali-Sin. 2021, 36, 18–23. (In Chinese) [Google Scholar]
- Zhou, Z.; Wang, X. Rational design and structure-based engineering of alkaline pectate lyase from Paenibacillus sp. 0602 to improve thermostability. BMC Biotechnol. 2021, 21, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Huang, S.; Wang, Z.; Fu, J.; Lv, P.; Miao, C.; Liu, T.; Yang, L.; Luo, W. Enhanced activity of Rhizomucor miehei lipase by directed saturation mutation of the propeptide. Enzyme. Microb. Technol. 2021, 150, 109870. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, L.; Wang, W.; Zou, M.; Ding, T.; Ye, X.; Liu, D. Synergistic effect and mechanisms of combining ultrasound and pectinase on pectin hydrolysis. Food Bioprocess Technol. 2016, 9, 1249–1257. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 277, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Liu, Z.; Duan, S.; Sun, Q.; Peng, Y.; Feng, X.; Zheng, K.; Hu, Z.; Zhang, Y. A rapid process of ramie bio-degumming by Pectobacterium sp. CXJZU-120. Text. Res. J. 2012, 82, 1–7. [Google Scholar] [CrossRef]
- Steven, H.R.; Robert, S.D.; Michael, G.; Margaret, L.; Frances, J. Characterization and implications of Ca2+ binding to pectate lyase C. J. Biol. Chem. 2003, 278, 12271. [Google Scholar]
- Kluskens, L.D.; Gert-Jan, M.A.; Alphons, G.J.V.; Willem, V.M.; Van der, O.J. Molecular and biochemical characterization of the thermoactive family 1 pectate lyase from the hyperthermophilic bacterium Thermotoga maritime. Biochem. J. 2003, 370, 651–659. [Google Scholar] [CrossRef] [Green Version]
- Seyedarabi, A.; Teng, T.T.; Ali, S.; Hussain, S.; Fries, M.; Madsen, R.; Clausen, M.H.; Teixteira, S.; Brocklehurst, K.; Pickersgill, R.W. Structural insights into substrate specificity and the anti beta-elimination mechanism of pectate lyase. Biochemistry 2010, 49, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Joyet, P.; Declerck, N.; Gaillardin, C. Hyperthermostable variants of a highly thermostable alpha-amylase. Bio/Technology 1992, 10, 1579–1583. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.N.; Tan, J.; Wang, Y.; Wang, J. Enhancing the thermostability of a mono- and diacylglycerol lipase from Malassizia globose by stabilizing a flexible loop in the catalytic pocket. Enzyme. Microb. Technol. 2021, 149, 109849. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Wang, D.; Lv, C.; Zhang, Y.; Gelbic, I.; Ye, X. Archives of microbiology: Screening of pectinase-producing bacteria from citrus peel and characterization of a recombinant pectate lyase with applied potential. Arch. Microbiol. 2020, 202, 1005–1013. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Y.; Chang, Z.; Wang, H.; Leier, A.; Marquez-Lago, T.T.; Ma, Y.; Li, J.; Song, J. Structure-based engineering of a pectate lyase with improved specific activity for ramie degumming. Appl. Microbiol. Biotechnol. 2017, 101, 2919–2929. [Google Scholar] [CrossRef]
- Li, G.; Fang, X.; Su, F.; Chen, Y.; Xu, L.; Yan, Y. Enhancing the thermostability of Rhizomucor miehei lipase with a limited screening library by rational-design point mutations and disulfide bonds. Appl. Environ. Microbiol. 2018, 84, e02129-17. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Gui, X.; Zhou, C.; Xue, Y.; Ma, Y.; Tang, S. Improving the thermoactivity and thermostability of pectate lyase from Bacillus pumilus for ramie degumming. Appl. Microbiol. Biotechnol. 2015, 99, 2673–2682. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Primer Sequences |
|---|---|
| A41V | F: 5’ CGGGCGGTTATGTGACCACTTCCGG 3’ R: 5’ CCGGAAGTGGTCACATAACCGCCCG 3’ |
| A82V | F: 5’ GGTGAAAGGCGGCGTATACCCGCTGGTCATC 3’ R: 5’ GATGACCAGCGGGTATACGCCGCCTTTCACC 3’ |
| A112V | F: 5’ CAGTGGAGCAAAGACGTACGCGGCGTGGAAATC 3’ R: 5’ GATTTCCACGCCGCGTACGTCTTTGCTCCACTG 3’ |
| A112P | F: 5’GGCCAGTGGAGCAAAGACCCACGCGGCGTGGAAATCAAAG 3’ R: 5’ CTTTGATTTCCACGCCGCGTGGGTCTTTGCTCCACTGGCC 3’ |
| A129V | F: 5’ CACCATCATCGGCGTGAACGGTTCTTCCGC 3’ R: 5’ GCGGAAGAACCGTTCACGCCGATGATGGTG 3’ |
| A129G | F: 5’ CCATCATCGGCGGCAACGGTTCTTC 3’ R: 5’ GAAGAACCGTTGCCGCCGATGATGG 3’ |
| A129P | F: 5’ CACCATCATCGGCCCCAACGGTTCTTC 3’ R: 5’ GAAGAACCGTTGGGGCCGATGATGGTG 3’ |
| A160V | F: 5’ CTACCTGCCGGGCGGCGTACAGGATGGCGATATGTTC 3’ R: 5’ GAACATATCGCCATCCTGTACGCCGCCCGGCAGGTAG 3 |
| A201G | F: 5’ CCACGTTCGAATCCGGATTTGACATCAAGAAAG 3’ R: 5’ CTTTCTTGATGTCAAATCCGGATTCGAACGTGG 3’ |
| A201P | F: 5’ CACCACGTTCGAATCCCCGTTTGACATCAAGAAAG 3’ R: 5’ CTTTCTTGATGTCAAACGGGGATTCGAACGTGGTG 3’ |
| D333E | F: 5’ CCTGGACGGCGGAAACCAAAGCTTACG 3’ R: 5’ CGTAAGCTTTGGTTTCCGCCGTCCAGG 3’ |
| D341E | F: 5’ CGTCAACGCCGAAAGCTGGACTTCC 3’ R: 5’ GGAAGTCCAGCTTTCGGCGTTGACG 3’ |
| CK | PelG403 | A129V | Commercial Enzyme | |
|---|---|---|---|---|
| Weightlessness rate (%) | 5.77 | 12.07 | 18.33 | 19.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Duan, S.; Feng, X.; Yang, Q.; Zheng, K.; Peng, Y.; Cheng, L. Improving the Thermo-Activity and -Stability of Pectate Lyase from Dickeya dadantii DCE-01 for Ramie Degumming. Processes 2021, 9, 2106. https://doi.org/10.3390/pr9122106
Xu H, Duan S, Feng X, Yang Q, Zheng K, Peng Y, Cheng L. Improving the Thermo-Activity and -Stability of Pectate Lyase from Dickeya dadantii DCE-01 for Ramie Degumming. Processes. 2021; 9(12):2106. https://doi.org/10.3390/pr9122106
Chicago/Turabian StyleXu, Huan, Shengwen Duan, Xiangyuan Feng, Qi Yang, Ke Zheng, Yuande Peng, and Lifeng Cheng. 2021. "Improving the Thermo-Activity and -Stability of Pectate Lyase from Dickeya dadantii DCE-01 for Ramie Degumming" Processes 9, no. 12: 2106. https://doi.org/10.3390/pr9122106