A Fructan Sucrase Secreted Extracellular and Purified in One-Step by Gram-Positive Enhancer Matrix Particles


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Conditions, and Plasmids
2.2. Cloning of the Fructan Sucrase Gene
2.3. Determination of Inducement Conditions
2.4. Expression and Purification of the Recombinant Enzyme
2.5. Determination of the Enzyme Activity
2.6. Effect of pH, Temperature, and Ions on the Activity of the BSO Recombinant Fructan Sucrase
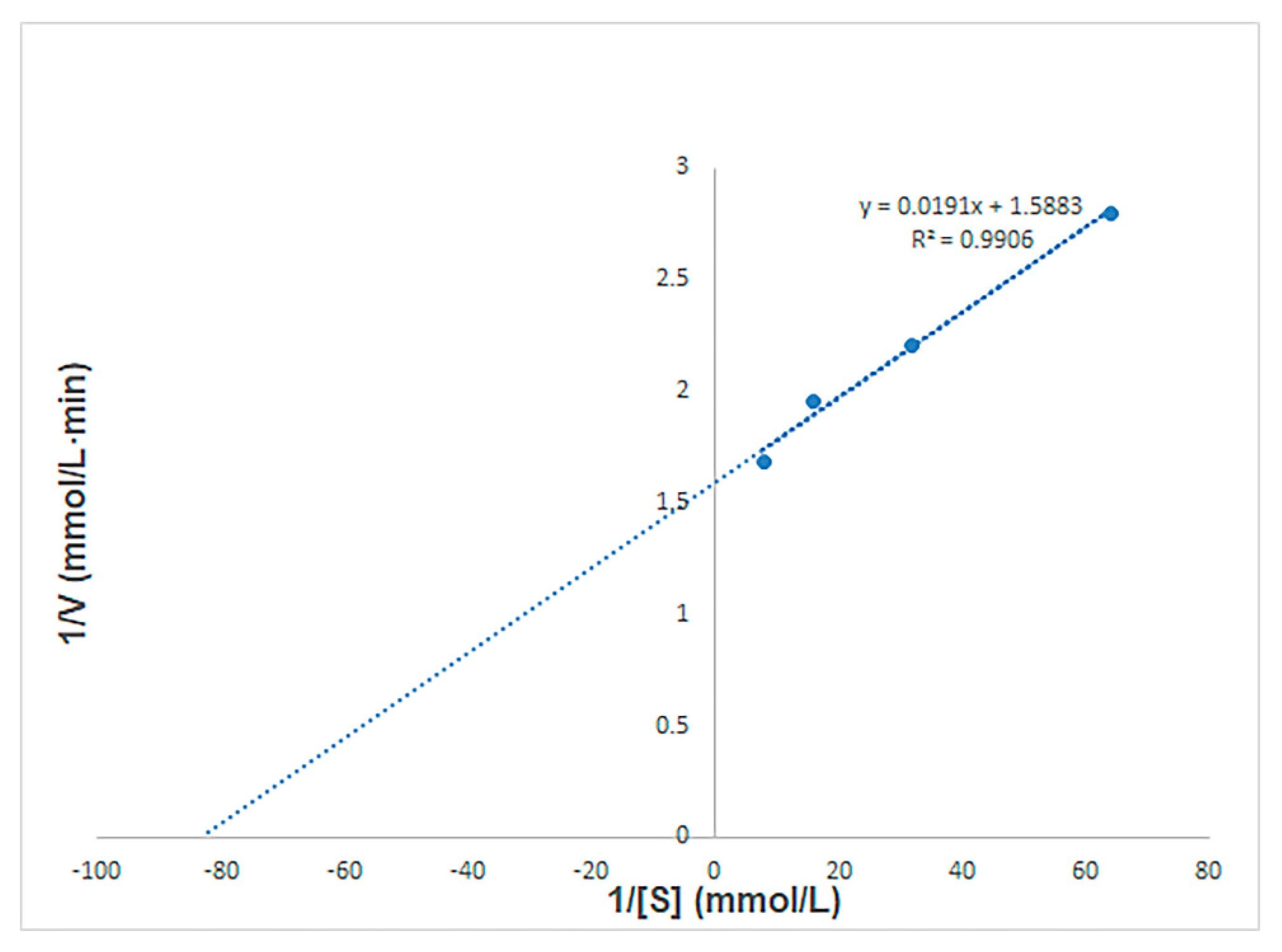
2.7. Determining the Kinetic Parameters of BSO Recombinant Fructan Sucrase
2.8. Data Analysis
3. Results and Discussion
3.1. Cloning of the Fructan Sucrase Gene
3.2. Effect of Temperature, Time, IPTG Concentration on Induced Expression
3.2.1. Effect of Temperature on Induction Expression
3.2.2. Effect of Time on Induced Expression
3.2.3. Effect of IPTG Concentration on Induced Expression
3.3. Expression and Purification of Recombinant Fructan Sucrase
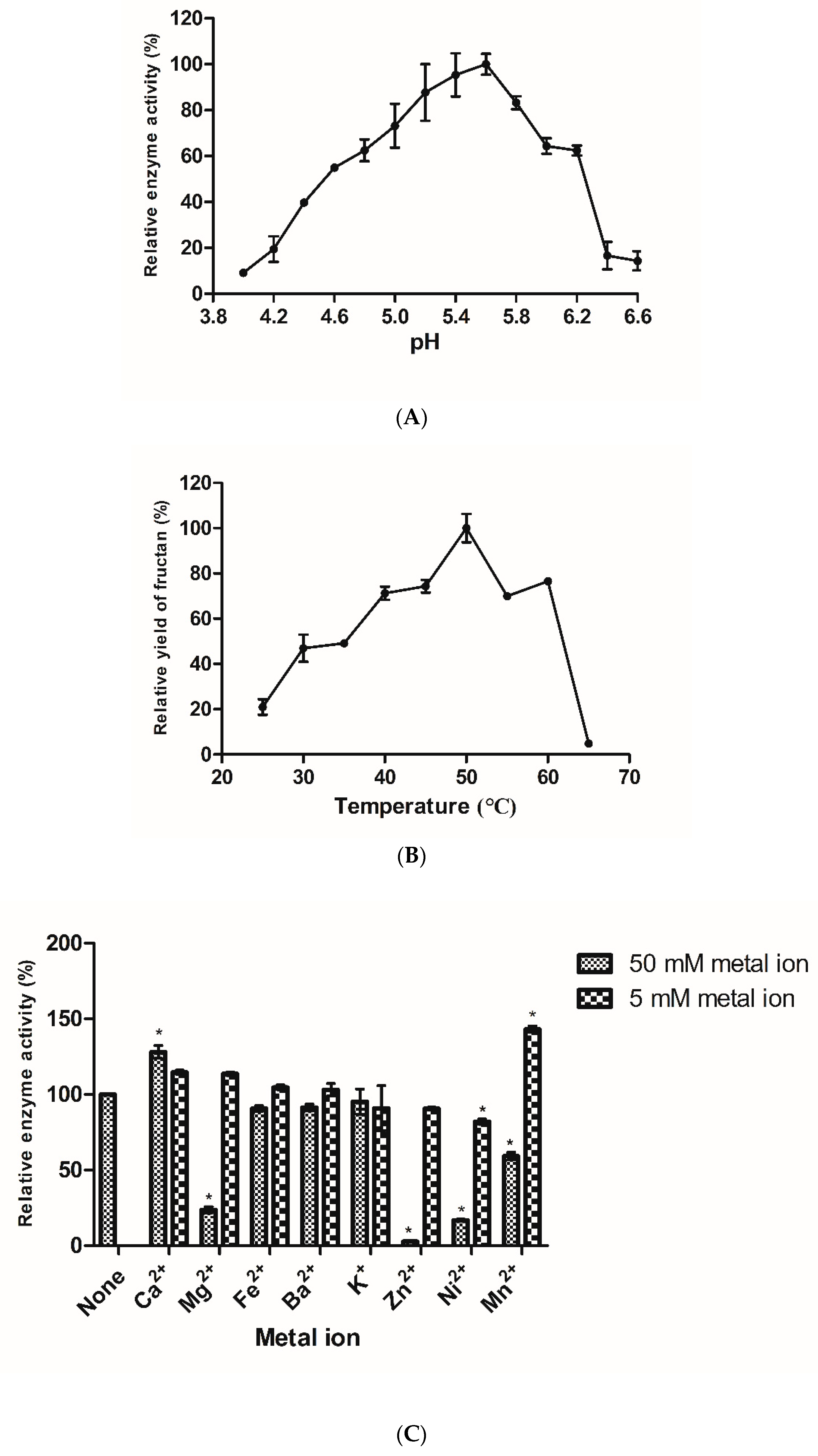
3.4. Effect of pH, Temperature, and Ions on the Activity of Recombinant Fructan Sucrase
3.4.1. Effect of pH on the Activity of Fructan Sucrase
3.4.2. Effect of Temperature on the Activity of Fructan Sucrase
3.4.3. Effect of Ions on the Activity of Recombinant Fructan Sucrase
3.5. The Kinetic Parameters of Recombinant Fructan Sucrase
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belghith, K.S.; Dahech, I.; Hamden, K.; Feki, A.; Mejdoub, H.; Belghith, H. Hypolipidemic effect of diet supplementation with bacterial levan in cholesterol-fed rats. Int. J. Biol. Macromol. 2012, 50, 1070–1074. [Google Scholar] [CrossRef]
- Byun, B.Y.; Lee, S.J.; Mah, J.H. Antipathogenic activity and preservative effect of levan (β-2, 6-fructan), a multifunctional polysaccharide. Int. J. Food Sci. Technol. 2014, 49, 238–245. [Google Scholar] [CrossRef]
- Dahech, I.; Harrabi, B.; Hamden, K.; Feki, A.; Mejdoub, H.; Belghith, H.; Belghith, K.S. Antioxidant effect of nondigestible levan and its impact on cardiovascular disease and atherosclerosis. Int. J. Biol. Macromol. 2013, 58, 281–286. [Google Scholar] [CrossRef]
- Srikanth, R.; Siddartha, G.; Reddy, C.H.; Harish, B.S.; Ramaiah, M.J.; Uppuluri, K.B. Antioxidant and anti-inflammatory levan produced from Acetobacter xylinum NCIM2526 and its statistical optimization. Carbohydr. Polym. 2015, 123, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Methus, K.; Rath, P.; Thanapon, C.; Karan, W.; Surasak, C. Computational design of oligosaccharide producing levansucrase from Bacillus licheniformis RN-01 to improve its thermostability for production of levan-type fructooligosaccharides from sucrose. Int. J. Biol. Macromol. 2020, 160, 252–263. [Google Scholar] [CrossRef]
- Srikanth, R.; Reddy, C.H.; Siddartha, G.; Ramaiah, M.J.; Uppuluri, K.B. Uppuluri review on production, characterization and applications of microbial levan. Carbohydr. Polym. 2015, 120, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Ragab, T.I.M.; Shalaby, A.S.G.; Awdan, S.A.E.; El-Bassyouni, G.T.; Salama, B.M.; Helmy, W.A.; Esawy, M.A. Role of levan extracted from bacterial honey isolates in curing peptic ulcer: In vivo. Int. J. Biol. Macromol 2020, 142, 64–573. [Google Scholar] [CrossRef] [PubMed]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.A. Pandey Applications of microbial enzymes in food industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Lalitagauri, R.; Sunita, P.; Debabrata, B. Enzymes—An existing and promising tool of food processing industry. Recent Pat. Biotechnol. 2016, 10, 58–71. [Google Scholar] [CrossRef]
- Trujilloa, L.E.; Arrietaa, J.G.; Dafhnisa, F.; Garcíab, J.; Valdésb, J.; Tambarac, Y.; Pérezb, M.; Hernándeza, L. Fructo-oligosaccharides production by the Gluconacetobacter diazotrophicus levansucrase expressed in the methylotrophic yeast Pichia pastoris. Enzym. Microb Technol. 2001, 28, 139–144. [Google Scholar] [CrossRef]
- Gao, S.; Qi, X.; Hart, D.J.; Gao, H.; An, Y. Expression and characterization of levansucrase from Clostridium acetobutylicum. J. Agric. Food Chem. 2017, 65, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Xu, W.; Bai, Y.; Zhang, W.; Zhang, T.; Mu, W. Biosynthesis of levan from sucrose using a thermostable levansucrase from Lactobacillus reuteri LTH5448. Int. J. Biol. Macromol. 2018, 113, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.R.; Ramana, K.V. Optimization of levan production by cold-active Bacillus licheniformis ANT 179 and fructooligosaccharide synthesis by its levansucrase. Appl. Biochem. Biotechnol. 2017, 181, 986–1006. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.; Vasileva, T.; Bivolarski, V.; Salim, A.; Morel, S.; Rabier, P.; Gabriel, V. Optimization of the expression of levansucrase L17 in recombinant E. coli. Biotechnol. Biotechnol. Equip. 2018, 32, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Ishida, R.; Sakaguchi, K.; Matsuzaki, C.; Katoh, T.; Ishida, N.; Yamamoto, K.; Hisa, K. Levansucrase from Leuconostoc mesenteroides NTM048 produces a levan exopolysaccharide with immunomodulating activity. Biotechnol. Lett. 2016, 38, 681–687. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, S.H.; Zhang, T.; Jing, B.; Mu, W.M. Efficient biosynthesis of levan from sucrose by a novel levansucrase from Brenneria goodwinii. Carbohydr. Polym. 2017, 157, 1732–1740. [Google Scholar] [CrossRef]
- Jong, W.S.; Saurí, A.; Luirink, J. Extracellular production of recombinant proteins using bacterial autotransporters. Curr. Opin. Biotechnol. 2010, 21, 646–652. [Google Scholar] [CrossRef]
- Choi, J.H.; Lee, S.Y. Secretory and extracellular production of recombinant proteins using Escherichia coli. Appl. Microbiol. Biotechnol. 2004, 64, 625–635. [Google Scholar] [CrossRef]
- Su, L.; Xu, C.; Woodard, R.W.; Chen, J.; Wu, J. A novel strategy for enhancing extracellular secretion of recombinant proteins in Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 6705–6713. [Google Scholar] [CrossRef]
- Sambasivam, G.; Nair, S.; Karthikeyan, G. Effect of codon-optimized E-coli signal peptides on recombinant Bacillus stearothermophilus maltogenic amylase periplasmic localization, yield and activity. J. Ind. Microbiol. Biotechnol. 2014, 41, 1435–1442. [Google Scholar] [CrossRef]
- Chen, J.; Gai, Y.; Fu, G.; Zhou, W.; Zhang, D.; Wen, J. Enhanced extracellular production of α-amylase in Bacillus subtilisby optimization of regulatory elements and over-expression of PrsA lipoprotein. Biotechnol. Lett. 2015, 37, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Angkawidjaja, C.; Kuwahara, K.; Omori, K.; Koga, Y.; Takano, K.; Kanaya, S. Extracellular secretion of Escherichia coli alkaline phosphatase with a C-terminal tag by type I secretion system: Purification and biochemical characterization. Protein. Eng. Des. Sel. 2006, 19, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.F.; Li, B.; Liu, Z.G.; Wang, M.; Gu, Z.B.; Du, G.C.; Wu, J.; Chen, J. Calcium leads to further increase in glycine-enhanced extracellular secretion of recombinant alpha-cyclodextrin glycosyltransferase in Escherichia coli. J. Agric. Food Chem. 2009, 57, 6231–6237. [Google Scholar] [CrossRef] [PubMed]
- Matos, C.F.R.O.; Branston, S.D.; Albiniak, A.; Dhanoya, A.; Robinson, C. High-yield export of a native heterologous protein to the periplasm by the Tat translocation pathway in Escherichia coli. Biotechnol. Bioeng. 2012, 109, 2533–2542. [Google Scholar] [CrossRef] [PubMed]
- Kujau, M.J.; Hoischen, C.; Riesenberg, D.; Gumpert, J. Expression and secretion of functional miniantibodies McPC603scFvDhlx in cell-wall-less L-form strains of Proteus mirabilisand Escherichia coli: A comparison of the synthesis capacities of L-form strains with an E. coli producer strain. Appl. Microbiol. Biotechnol. 1998, 49, 51–58. [Google Scholar] [CrossRef]
- Lech, M. Optimisation of protein-free waste whey supplementation used for the industrial microbiological production of lactic acid. Biochem. Eng. J. 2020, 157, 107531. [Google Scholar] [CrossRef]
- Wood, E.J. Molecular cloning, a laboratory manual. Biochem. Educ. 1983, 11, 182–183. [Google Scholar] [CrossRef]
- Liu, S.L.; Du, K.; Chen, W.Z.; Liu, G.; Xing, M. Effective approach to greatly enhancing selective secretion and expression of three cytoplasmic enzymes in Escherichia coli through synergistic effect of EDTA and lysozyme. J. Ind. Microbiol. Biotechnol. 2012, 39, 1301–1307. [Google Scholar] [CrossRef]
- Guo, Y.; Bian, W.; Zhang, Y. Expression in Escherichia coli, purification and characterization of LRSAM1, a LRR and RING domain E3 ubiquitin ligase. Protein Expres Purif. 2017, 129, 158–161. [Google Scholar] [CrossRef]
- Zhao, F.K.; Song, Q.Z.; Wang, B.B.; Du, R.P.; Han, Y.; Zhou, Z.J. Secretion of the recombination α-amylase in Escherichia coli and purification by the gram-positive enhancer matrix (GEM) particles. Int. J. Biol. Macromol. 2019, 123, 91–96. [Google Scholar] [CrossRef]
- Li, P.C.; Qiao, X.W.; Zheng, Q.S.; Hou, J.B. Immunogenicity and immunoprotection of porcine circovirus type 2 (PCV2) Cap protein displayed by Lactococcus lactis. Vaccine 2016, 34, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Saluja, V.; Visser, M.R.; Veer, W.T.; Roosmalen, M.L.V.; Leenhouts, K.; Hinrichs, W.L.J.; Huckriede, A.; Frijlink, H.W. Influenza antigen-sparing by immune stimulation with Gram-positive enhancer matrix (GEM) particles. Vaccine 2010, 28, 7963–7969. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.L.; Qin, W.T.; Xu, J.T.; Yan, Y.R.; Tian, J.; Wu, N.F.; Yao, B. Enhancing the soluble expression of an amylase in Escherichia coli by the mutations related to its domain interactions. Protein Expr. Purif. 2016, 120, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Sumitomo, N.; Ozaki, K.; Kawai, S.; Ito, S. Nucleotide sequence of the gene for an alkaline endoglucanase from an Alkalophilic Bacillus and its expression in Escherichia coli and Bacillus subtilis. Biosci. Biotechnol. Biochem. 1992, 6, 872–877. [Google Scholar] [CrossRef]
- Solingen, P.; Meijer, D.; Kleij, W.A.; Barnett, C.; Bolle, R.; Power, S.D.; Jones, B.E. Cloning and expression of an endocellulase gene from a novel Streptomycete isolated from an East African soda lake. Extremophiles 2001, 5, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Srensen, H.P.; Mortensen, K.K. Advanced genetic strategies for recombinant protein expression in Escherichia coli. J. Biotechnol. 2005, 115, 113–128. [Google Scholar] [CrossRef]
- Meng, F.Q.; Zhao, H.Z.; Zhang, C.; Lu, F.X.; Bie, X.M. Expression of a novel bacteriocin—The plantaricin Pln1—In Escherichia coli and its functional analysis. Protein Expr. Purif. 2016, 119, 85–93. [Google Scholar] [CrossRef]
- Laymon, R.A.; Adney, W.S.; Mohagheghi, A.; Himmel, M.E.; Thomas, S.R. Cloning and expression of full-length Trichoderma reesi cellobiohydrolase I cDNAs in Escherichia coli. Appl. Biochem. Biotechnol. 1996, 57–58, 389–397. [Google Scholar] [CrossRef]
- Tang, B.; Pan, H.; Zhang, Q.; Ding, L. Cloning and expression of cellulase gene EG1 from Rhizopus stolonifer var. reflexus TP-02 in Escherichia coli. Bioresour. Technol. 2009, 100, 6129–6132. [Google Scholar] [CrossRef]
- Zhao, B.; Du, R.; Wang, J.Y.; Xu, M.; Han, Y.; Zhou, Z.J. Purification and biochemical characterization of a novel glucansucrase from Leuconostoc citreum B-2. Biotechnol. Lett. 2020, 42, 1535–1545. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, L.; Guo, W.; Jia, L.B.; Fu, Y.; Gui, S.; Lu, F.P. Cloning, expression, and characterization of a thermostable and pH-stable laccase from Klebsiella pneumoniae and its application to dye decolorization. Process Biochem. 2016, 53, 125–134. [Google Scholar] [CrossRef]
- Malik, B.; Rashid, N.; Ahmad, N.; Akhtar, M. Escherichia coli signal peptidase recognizes and cleaves the signal sequence of α-amylase originating from Bacillus licheniformis. Biochemistry (Moscow) 2013, 78, 958–962. [Google Scholar] [CrossRef] [PubMed]
- Hövels, M.; Kosciow, K.; Kniewel, J.; Jakob, F.; Deppenmeier, U. High yield production of levan-type fructans by Gluconobacter japonicus LMG 1417. Int. J. Biol. Macromol. 2020, 164, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.N.D.; Oliveira, L.A.D.; Andrade, J.S. Partial characterization of amylases of two Indigenous Central Amazonian Rhizobia Strains. Braz. Arch. Biol. Techn. 2010, 53, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Ding, R.; Li, Z.; Chen, S.; Wu, D.; Wu, J.; Chen, J. Enhanced secretion of recombinant α-cyclodextrin glucosyltransferase from E. coli by medium additives. Process. Biochem. 2010, 45, 880–886. [Google Scholar] [CrossRef]
- Fuhrmann, G.F.; Völker, B. Misuse of graphical analysis in nonlinear sugar transport kinetics by Eadie-Hofstee plots. Biochim. Biophys. Acta 1993, 1145, 180–182. [Google Scholar] [CrossRef]
- Xu, W.; Ni, D.; Zhang, W.; Guang, C.; Zhang, T.; Mu, W. Recent advances in Levansucrase and Inulosucrase: Evolution, characteristics, and application. Crit. Rev. Food Sci. Nutr. 2019, 59, 3630–3647. [Google Scholar] [CrossRef]
- Hill, A.; Chen, L.; Mariage, A.; Petit, J.L.; Berardinis, V.D.; Karboune, S. Discovery of new levansucrase enzymes with interesting properties and improved catalytic activity to produce levan and fructooligosaccharides. Catal. Sci. Technol. 2019, 9, 2931–2944. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence |
|---|---|
| Primers of cloning of fructan sucrase gene from B. subtilis | |
| BS-F | 5′-CCGCGGATCCAAAGAAACGAACCAAAAG-3′ (containing BamHIsite) |
| BS-R | 5′-CCGCCTGCAGTTTGTTAACTGTTAATTG-3′ (containing PstIsite) |
| Primers of cloning of fructan sucrase and secretory signal peptide gene from B. subtilis | |
| BS-FF-F | 5′-CCGCGGATCCAACATCAAAAAGTTTGC-3′ (containing BamHIsite) |
| BS-FF-R | 5′-CCGCCTGCAGTTTGTTAACTGTTAATTG-3′ (containing PstIsite) |
| Primers of recombinant expression vectors of fructan sucrase from B. subtilis and signal peptide genes secreted by E. coli | |
| BSO-F | 5′-CGCGGATCCAAAAAAACCGCTATCGCTATCGCTGTTGCTCTGGCTGGTTTCGCTA CCGTTGCTCAGGCTAAAGAAACGAACCAAAAG-3′ (containing BamHIsite) |
| BSO-R | 5′-CCGCCTGCAGTTTGTTAACTGTTAATTG-3′ (containing PstIsite) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Xiao, H.; Zhao, F.; Zhao, B.; Xu, M.; Zhou, Z.; Han, Y. A Fructan Sucrase Secreted Extracellular and Purified in One-Step by Gram-Positive Enhancer Matrix Particles. Processes 2021, 9, 95. https://doi.org/10.3390/pr9010095
Wang J, Xiao H, Zhao F, Zhao B, Xu M, Zhou Z, Han Y. A Fructan Sucrase Secreted Extracellular and Purified in One-Step by Gram-Positive Enhancer Matrix Particles. Processes. 2021; 9(1):95. https://doi.org/10.3390/pr9010095
Chicago/Turabian StyleWang, Jingyue, Huazhi Xiao, Fangkun Zhao, Bo Zhao, Min Xu, Zhijiang Zhou, and Ye Han. 2021. "A Fructan Sucrase Secreted Extracellular and Purified in One-Step by Gram-Positive Enhancer Matrix Particles" Processes 9, no. 1: 95. https://doi.org/10.3390/pr9010095