Choline-Based Ionic Liquids as Media for the Growth of Saccharomyces cerevisiae

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Synthesis of ILs
2.2.2. Microorganism and Culture Conditions
2.2.3. Minimum Inhibitory Concentration (MIC)
2.2.4. Screening of ILs Concentration
3. Results and Discussion
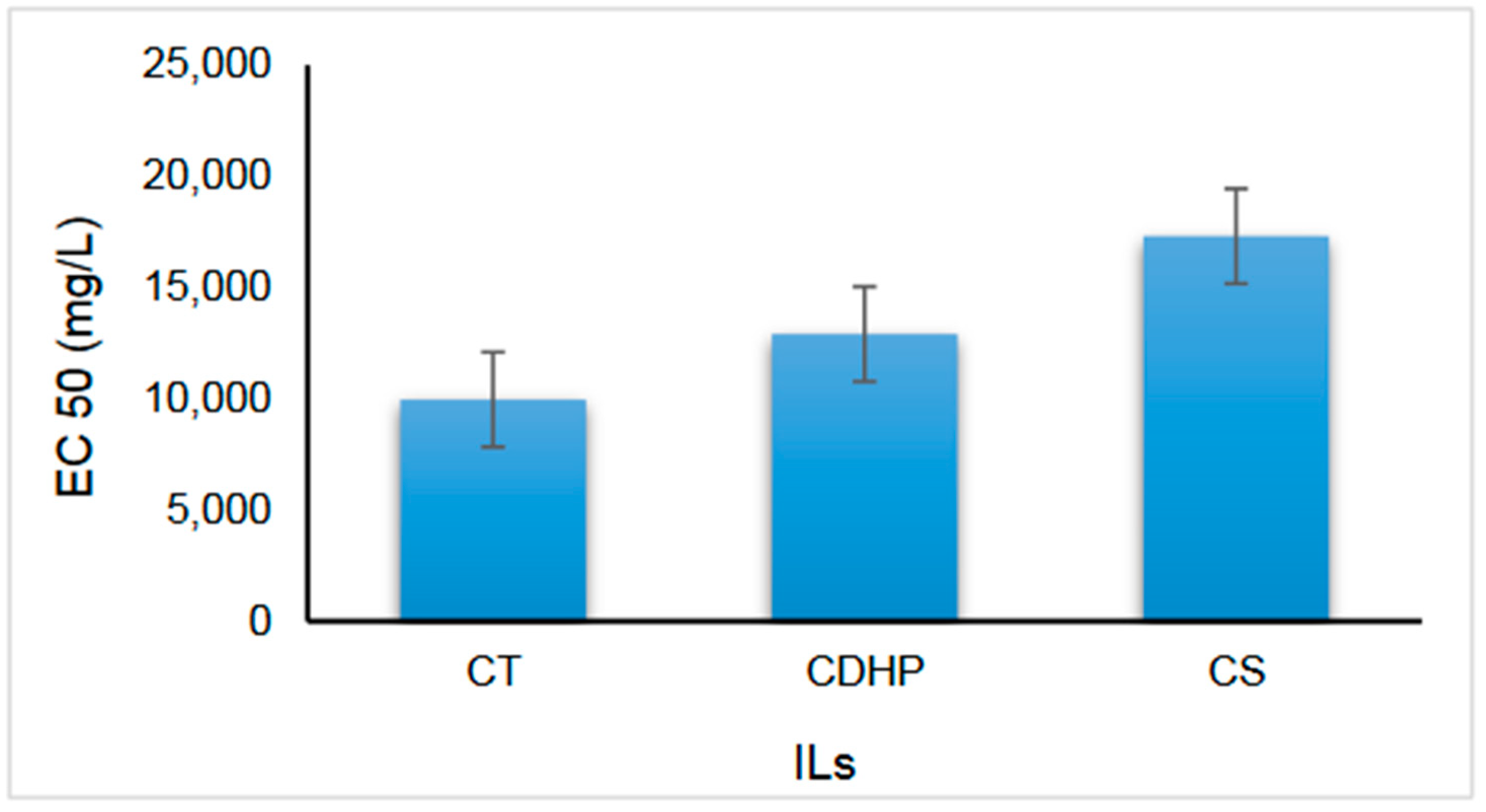
3.1. Toxicity of ILs
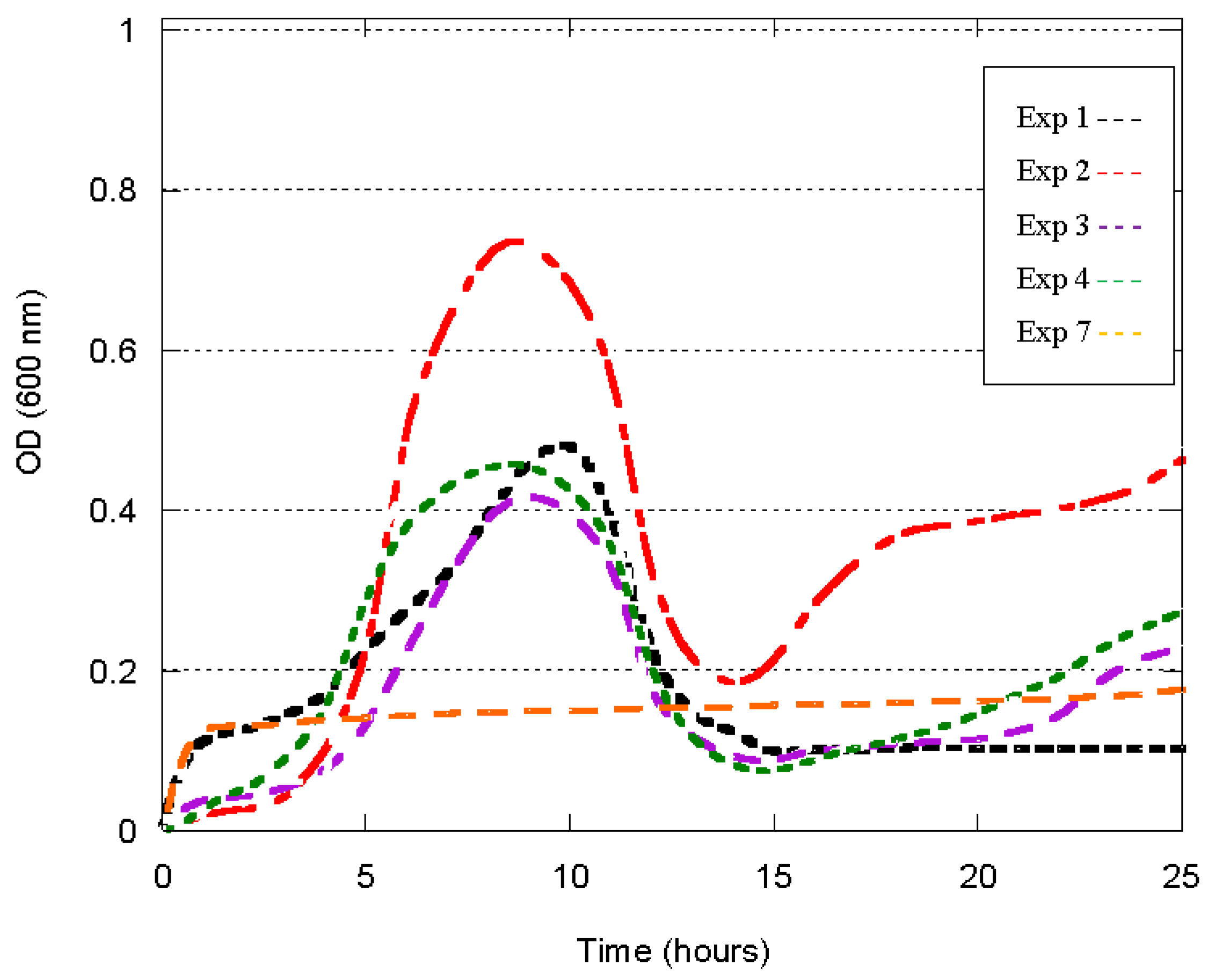
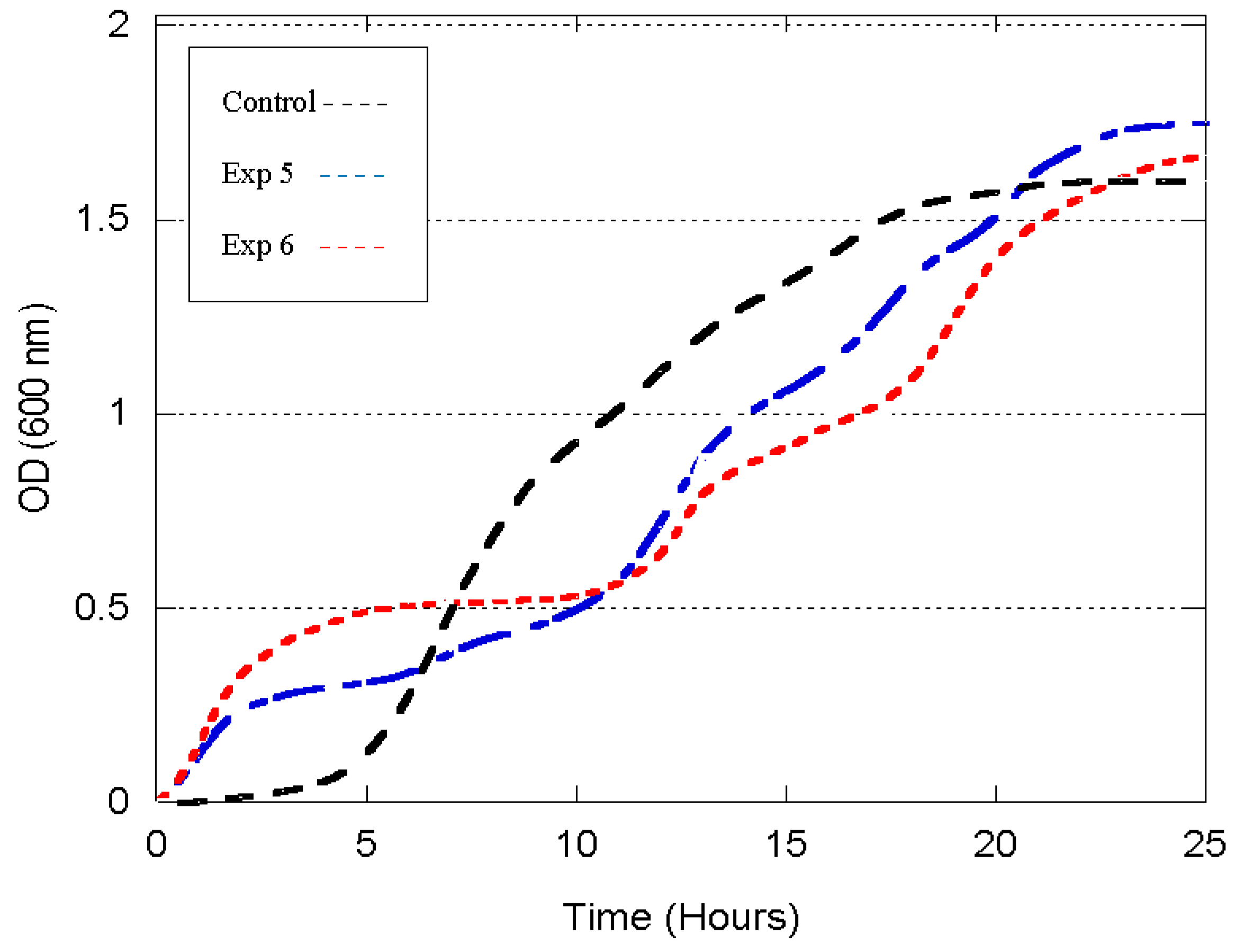
3.2. Growth Profile
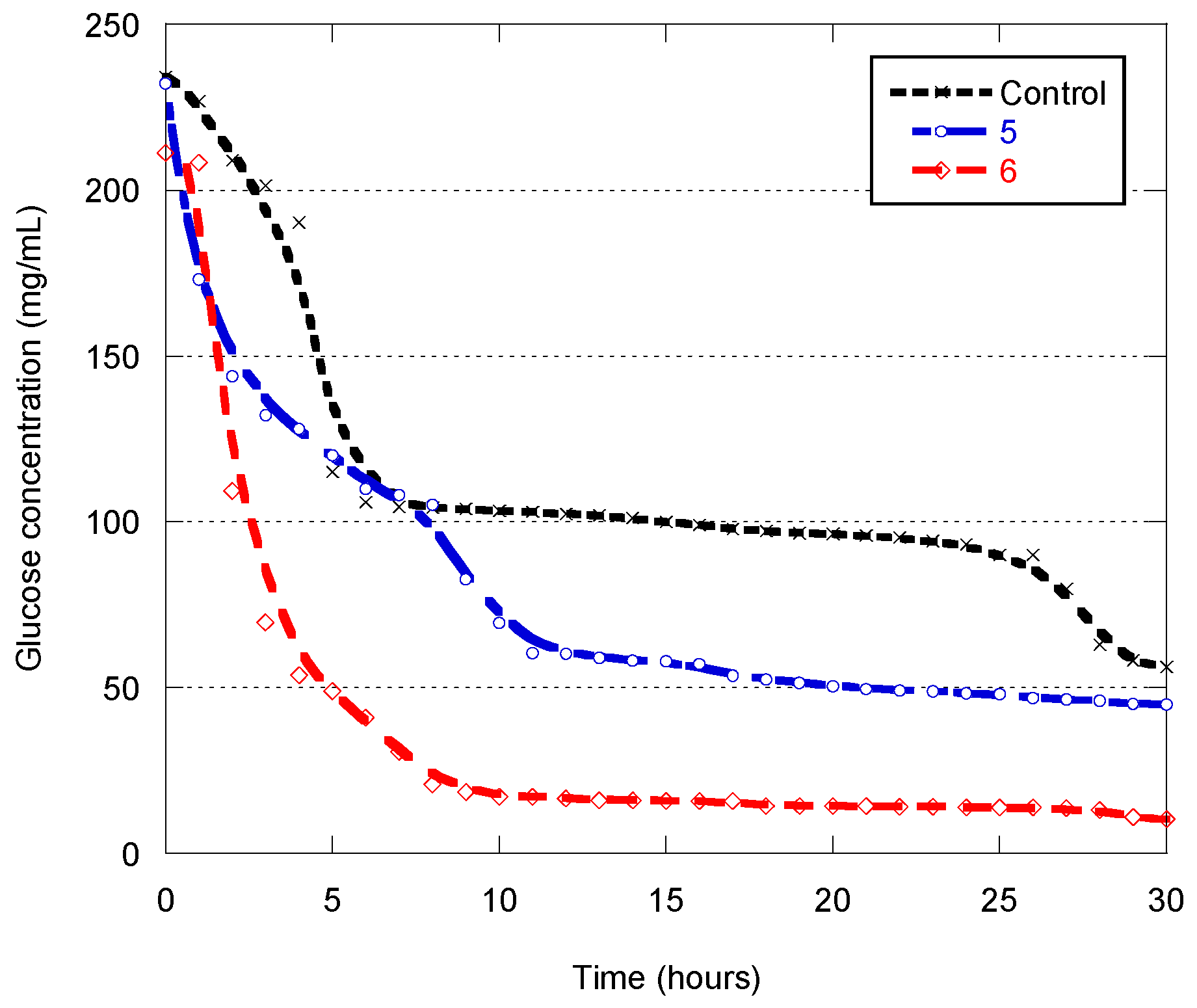
3.3. Glucose Utilization Profile
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviation
| emim[Cl] | 1-ethyl-3-methylimidazolium chloride |
| bmim[Cl] | 1-butyl-3-methylimidazolium chloride |
| emim[OAc] | 1-ethyl-3-methylimidazolium acetate |
| emim[CH3PO4] | 1-ethyl-3-methylimidazolium methylphosphonate |
| DMEAA | N,N-dimethylethanolammonium acetate |
| [EtOHNMe3][Me2PO4] | 2-hydroxyethyl trimethyl ammonium dimethyl phosphate |
| emim[MeO(H)PO2] | 1-ethyl-3-methylimidazolium methylphosphonate |
| omimPF6 | 1-methyl-3-octylimidazolium hexafluorophosphate |
| YPD | Yeast Extract-Peptone-Dextrose |
| FDA | Food and Drug Administration |
References
- Charoenrat, T.; Khumruaengsri, N.; Promdonkoy, P.; Rattanaphan, N.; Eurwilaichitr, L.; Tanapongpipat, S.; Roongsawang, N. Improvement of recombinant endoglucanase produced in Pichia pastoris KM71 through the use of synthetic medium for inoculum and pH control of proteolysis. J. Biosci. Bioeng. 2013, 116, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Ott, F.D. Synthetic media and techniques for the xenic cultivation of marine algae and flagellate. Va. J. Sci. 1965, 16, 205–218. [Google Scholar]
- Carvalho, A.V.; da Costa Lopes, A.M.; Bogel-Łukasik, R. Relevance of the acidic 1-butyl-3-methylimidazolium hydrogen sulphate ionic liquid in the selective catalysis of the biomass hemicellulose fraction. RSC Adv. 2015, 5, 47153–47164. [Google Scholar] [CrossRef] [Green Version]
- Monod, J. The phenomenon of enzymatic adaptation. In Selected Papers in Molecular Biology; Monod, J., Ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 68–134. [Google Scholar]
- Arzumanian, V. Synthetic media for cultivation of lipophilic yeast malassezia spp. Vestnik Rossiiskoi Akademii Meditsinskikh Nauk 1999, 11, 54–56. [Google Scholar]
- Aguilar-Uscanga, B.; Francois, J. A study of the yeast cell wall composition and structure in response to growth conditions and mode of cultivation. Lett. Appl. Microbiol. 2003, 37, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, A.; Sakaguchi, N.; Shigeto, S. Can cells maintain their bioactivity in ionic liquids? A novel single-cell assessment by Raman microspectroscopy. J. Raman Spectrosc. 2019. [Google Scholar] [CrossRef]
- Takaoka, T.; Katsuta, H. Long-term cultivation of mammalian cell strains in protein-and lipid-free chemically defined synthetic media. Exp. Cell Res. 1971, 67, 295–304. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Serrano-Amatriain, C.; Chen, Y.; Siewers, V.; Nielsen, J. Improving biobutanol production in engineered Saccharomyces cerevisiae by manipulation of acetyl-CoA metabolism. J. Ind. Microbiol. Biotechnol. 2013, 40, 1051–1056. [Google Scholar] [CrossRef]
- van Zutphen, T.; Todde, V.; de Boer, R.; Kreim, M.; Hofbauer, H.F.; Wolinski, H.; Veenhuis, M.; van der Klei, I.J.; Kohlwein, S.D. Lipid droplet autophagy in the yeast Saccharomyces cerevisiae. Mol. Biol. Cell 2014, 25, 290–301. [Google Scholar] [CrossRef]
- Schadeweg, V.; Boles, E. n-Butanol production in Saccharomyces cerevisiae is limited by the availability of coenzyme A and cytosolic acetyl-CoA. Biotechnol. Biofuels 2016, 9, 44. [Google Scholar] [CrossRef]
- Curran, K.A.; Leavitt, J.M.; Karim, A.S.; Alper, H.S. Metabolic engineering of muconic acid production in Saccharomyces cerevisiae. Metab. Eng. 2013, 15, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Borodina, I.; Kildegaard, K.R.; Jensen, N.B.; Blicher, T.H.; Maury, J.; Sherstyk, S.; Schneider, K.; Lamosa, P.; Herrgård, M.J.; Rosenstand, I. Establishing a synthetic pathway for high-level production of 3-hydroxypropionic acid in Saccharomyces cerevisiae via β-alanine. Metab. Eng. 2015, 27, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Jakočiūnas, T.; Bonde, I.; Herrgård, M.; Harrison, S.J.; Kristensen, M.; Pedersen, L.E.; Jensen, M.K.; Keasling, J.D. Multiplex metabolic pathway engineering using CRISPR/Cas9 in Saccharomyces cerevisiae. Metab. Eng. 2015, 28, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Dipeolu, O.; Green, E.; Stephens, G. Effects of water-miscible ionic liquids on cell growth and nitro reduction using Clostridium sporogenes. Green Chem. 2009, 11, 397–401. [Google Scholar] [CrossRef]
- Fan, L.-L.; Li, H.-J.; Chen, Q.-H. Applications and mechanisms of ionic liquids in whole-cell biotransformation. Int. J. Mol. Sci. 2014, 15, 12196–12216. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.; Ferguson, J.L.; Gunaratne, H.Q.N.; Seddon, K.R.; Goodacre, R.; Stephens, G.M. Screening ionic liquids for use in biotransformations with whole microbial cells. Green Chem. 2011, 13, 1843–1851. [Google Scholar] [CrossRef]
- Welton, T. Room-Temperature Ionic Liquids. Solvents for Synthesis and Catalysis. Chem. Rev. 1999, 99, 2071–2084. [Google Scholar] [CrossRef]
- Earle, M.J.; Seddon, K.R. Ionic liquids. Green solvents for the future. Pure Appl. Chem. 2000, 72, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Zhang, C.; Crittle, T.D. Choline-based deep eutectic solvents for enzymatic preparation of biodiesel from soybean oil. J. Mol. Catal. B Enzym. 2013, 85–86, 243–247. [Google Scholar] [CrossRef]
- Hayyan, M.; Hashim, M.A.; Hayyan, A.; Al-Saadi, M.A.; AlNashef, I.M.; Mirghani, M.E.; Saheed, O.K. Are deep eutectic solvents benign or toxic? Chemosphere 2013, 90, 2193–2195. [Google Scholar] [CrossRef]
- Vrikkis, R.M.; Fraser, K.J.; Fujita, K.; MacFarlane, D.R.; Elliott, G.D. Biocompatible Ionic Liquids: A New Approach for Stabilizing Proteins in Liquid Formulation. J. Biomech. Eng. 2009, 131, 074514. [Google Scholar] [CrossRef] [PubMed]
- Couling, D.J.; Bernot, R.J.; Docherty, K.M.; Dixon, J.K.; Maginn, E.J. Assessing the factors responsible for ionic liquid toxicity to aquatic organisms via quantitative structure–property relationship modeling. Green Chem. 2006, 8, 82–90. [Google Scholar] [CrossRef]
- Weaver, K.D.; Kim, H.J.; Sun, J.; MacFarlane, D.R.; Elliott, G.D. Cyto-toxicity and biocompatibility of a family of choline phosphate ionic liquids designed for pharmaceutical applications. Green Chem. 2010, 12, 507–513. [Google Scholar] [CrossRef]
- Petkovic, M.; Ferguson, J.L.; Gunaratne, H.N.; Ferreira, R.; Leitao, M.C.; Seddon, K.R.; Rebelo, L.P.N.; Pereira, C.S. Novel biocompatible cholinium-based ionic liquids—Toxicity and biodegradability. Green Chem. 2010, 12, 643–649. [Google Scholar] [CrossRef]
- Vasylkovska, R.; Burdylyuk, N.; Semchyshyn, H. Involvement of catalase in Saccharomyces cerevisiae hormetic response to hydrogen peroxide. J. Vasyl Stefanyk Precarpathian Natl. Univ. 2015, 2, 107–114. [Google Scholar] [CrossRef]
- Rattan, S.I. The future of aging interventions: Aging intervention, prevention, and therapy through hormesis. J. Gerontol. Ser. A Biol. Sci. Med Sci. 2004, 59, B705–B709. [Google Scholar] [CrossRef]
- Berry, D.B.; Gasch, A.P. Stress-activated genomic expression changes serve a preparative role for impending stress in yeast. Mol. Biol. Cell 2008, 19, 4580–4587. [Google Scholar] [CrossRef]
- Calabrese, V.; Cornelius, C.; Mancuso, C.; Pennisi, G.; Calafato, S.; Bellia, F.; Bates, T.E.; Stella, A.M.G.; Schapira, T.; Kostova, A.T.D. Cellular stress response: A novel target for chemoprevention and nutritional neuroprotection in aging, neurodegenerative disorders and longevity. Neurochem. Res. 2008, 33, 2444–2471. [Google Scholar] [CrossRef]
- Lushchak, V.I. Dissection of the hormetic curve: Analysis of components and mechanisms. Dose-Response 2014, 12, 466–479. [Google Scholar] [CrossRef]
- Zhu, S.; Yu, P.; Lei, M.; Tong, Y.; Zheng, L.; Zhang, R.; Ji, J.; Chen, Q.; Wu, Y. Influence of the ionic liquid 1-butyl-3-methylimidazolium chlorine on the ethanol fermentation of Saccharomyces cerevisiae AY93161 and its kinetics analysis. Energy Educ. Sci. Technol. Part A Energy Sci. Res. 2013, 30, 817–828. [Google Scholar]
- Ouellet, M.; Datta, S.; Dibble, D.C.; Tamrakar, P.R.; Benke, P.I.; Li, C.; Singh, S.; Sale, K.L.; Adams, P.D.; Keasling, J.D. Impact of ionic liquid pretreated plant biomass on Saccharomyces cerevisiae growth and biofuel production. Green Chem. 2011, 13, 2743–2749. [Google Scholar] [CrossRef]
- Nancharaiah, Y.; Francis, A. Hormetic effect of ionic liquid 1-ethyl-3-methylimidazolium acetate on bacteria. Chemosphere 2015, 128, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, N.; Husson, E.; Jacquard, C.; Wewetzer, S.; Büchs, J.; Sarazin, C.; Gosselin, I. Impact of two ionic liquids, 1-ethyl-3-methylimidazolium acetate and 1-ethyl-3-methylimidazolium methylphosphonate, on Saccharomyces cerevisiae: Metabolic, physiologic, and morphological investigations. Biotechnol. Biofuels 2015, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Sitepu, I.R.; Shi, S.; Simmons, B.A.; Singer, S.W.; Boundy-Mills, K.; Simmons, C.W. Yeast tolerance to the ionic liquid 1-ethyl-3-methylimidazolium acetate. Fems Yeast Res. 2014, 14, 1286–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, H.; Okayama, H. A kinase from fission yeast responsible for blocking mitosis in S phase. Nature 1995, 374, 817. [Google Scholar] [CrossRef] [PubMed]
- Miller-Fleming, L.; Giorgini, F.; Outeiro, T.F. Yeast as a model for studying human neurodegenerative disorders. Biotechnol. J. Healthc. Nutr. Technol. 2008, 3, 325–338. [Google Scholar]
- Dickinson, Q.; Bottoms, S.; Hinchman, L.; McIlwain, S.; Li, S.; Myers, C.L.; Boone, C.; Coon, J.J.; Hebert, A.; Sato, T.K. Mechanism of imidazolium ionic liquids toxicity in Saccharomyces cerevisiae and rational engineering of a tolerant, xylose-fermenting strain. Microb. Cell Factories 2016, 15, 17. [Google Scholar] [CrossRef]
- Sendovski, M.; Nir, N.; Fishman, A. Bioproduction of 2-phenylethanol in a biphasic ionic liquid aqueous system. J. Agric. Food Chem. 2010, 58, 2260–2265. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.-D.; Liu, Q.-P.; Smith, T.J.; Li, N.; Zong, M.-H. Evaluation of toxicity and biodegradability of cholinium amino acids ionic liquids. PLoS ONE 2013, 8, e59145. [Google Scholar] [CrossRef]
- Gouveia, W.; Jorge, T.; Martins, S.; Meireles, M.; Carolino, M.; Cruz, C.; Almeida, T.; Araújo, M. Toxicity of ionic liquids prepared from biomaterials. Chemosphere 2014, 104, 51–56. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi, K.; Taniguchi, N.; Arai, S.; Yamada, R.; Katahira, S.; Ishida, N.; Takahashi, H.; Ogino, C.; Kondo, A. Direct bioethanol production from cellulose by the combination of cellulase-displaying yeast and ionic liquid pretreatment. Green Chem. 2011, 13, 2948–2953. [Google Scholar] [CrossRef]
- Passino, D.R.M.; Smith, S.B. Acute bioassays and hazard evaluation of representative contaminants detected in Great Lakes fish. Environ. Toxicol. Chem. 1987, 6, 901–907. [Google Scholar] [CrossRef]
- Gross, B.; Ronen, N.; Honigman, S.; Livne, E. Tryptophan toxicity—Time and dose response in rats. In Tryptophan, Serotonin, and Melatonin; Springer: Berlin/Heidelberg, Germany, 1999; pp. 507–516. [Google Scholar]
- Nockemann, P.; Thijs, B.; Driesen, K.; Janssen, C.R.; Van Hecke, K.; Van Meervelt, L.; Kossmann, S.; Kirchner, B.; Binnemans, K. Choline saccharinate and choline acesulfamate: Ionic liquids with low toxicities. J. Phys. Chem. B 2007, 111, 5254–5263. [Google Scholar] [CrossRef]
- Foureau, D.M.; Vrikkis, R.M.; Jones, C.P.; Weaver, K.D.; MacFarlane, D.R.; Salo, J.C.; McKillop, I.H.; Elliott, G.D. In vitro assessment of choline dihydrogen phosphate (CDHP) as a vehicle for recombinant human interleukin-2 (rhIL-2). Cell. Mol. Bioeng. 2012, 5, 390–401. [Google Scholar] [CrossRef]
- Zhu, S.; Yu, P.; Tong, Y.; Chen, R.; Lv, Y.; Zhang, R.; Lei, M.; Ji, J.; Chen, Q.; Wu, Y. Effects of the ionic liquid 1-butyl-3-methylimidazolium chloride on the growth and ethanol fermentation of Saccharomyces cerevisiae AY92022. Chem. Biochem. Eng. Q. 2012, 26, 105–109. [Google Scholar]
- Yoo, B.; Jing, B.; Jones, S.E.; Lamberti, G.A.; Zhu, Y.; Shah, J.K.; Maginn, E.J. Molecular mechanisms of ionic liquid cytotoxicity probed by an integrated experimental and computational approach. Sci. Rep. 2016, 6, 19889. [Google Scholar] [CrossRef]
- Yazdani, A.; Sivapragasam, M.; Levêque, J.; Moniruzzaman, M. Microbial biocompatibility and biodegradability of choline-amino acid based ionic liquids. J. Microb. Biochem. Technol. 2016, 8, 415–421. [Google Scholar] [CrossRef]
- Sivapragasam, M.; Moniruzzaman, M.; Goto, M. Recent advances in exploiting ionic liquids for biomolecules: Solubility, stability and applications. Biotechnol. J. 2016, 11, 1000–1013. [Google Scholar] [CrossRef]
- Fujita, K.; Nakamura, N.; Murata, K.; Igarashi, K.; Samejima, M.; Ohno, H. Electrochemical analysis of electrode-immobilized dehydrogenases in hydrated choline dihydrogen phosphate-type ionic liquid. Electrochim. Acta 2011, 56, 7224–7227. [Google Scholar] [CrossRef]
- Ganske, F.; Bornscheuer, U.T. Growth of Escherichia coli, Pichia pastoris and Bacillus cereus in the presence of the ionic liquids [BMIM][BF 4] and [BMIM][PF 6] and organic solvents. Biotechnol. Lett. 2006, 28, 465–469. [Google Scholar] [CrossRef]
- Cho, C.-W.; Jeon, Y.-C.; Pham, T.P.T.; Vijayaraghavan, K.; Yun, Y.-S. The ecotoxicity of ionic liquids and traditional organic solvents on microalga Selenastrum capricornutum. Ecotoxicol. Environ. Saf. 2008, 71, 166–171. [Google Scholar] [CrossRef]
- Brock, M.; Nickel, A.-C.; Madziar, B.; Blusztajn, J.K.; Berse, B. Differential regulation of the high affinity choline transporter and the cholinergic locus by cAMP signaling pathways. Brain Res. 2007, 1145, 1–10. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ILs | Molecular Weight, (g mol−1) | Physical Appearance | Thermal Properties | |||
|---|---|---|---|---|---|---|
| Cation | Anion | Abbreviation | TGA Decomposition Temperature, Tonset (°C) | DSC Glass Transition Temperature, Tg (°C) | ||
 Choline Choline |  Tryptophanate | CT | 307.39 | Brown, viscous liquid | 187.55 | −26.88 |
 Saccharinate | CS | 286.35 | Light yellow, viscous liquid | 248.82 | −62.33 | |
 Phosphate | CDP | 201.16 | White, viscous liquid | 229.85 | −24.63 | |
| Experiment | Media Constituent | Ratio |
|---|---|---|
| 1 | CT + CDHP + CS | 2:1:2 |
| 2 | Yeast Extract + CDHP + CS | 1:1:2 |
| 3 | CT + Peptone + CS | 2:2:2 |
| 4 | CT + CDHP + Glucose | 2:1:2 |
| 5 | Yeast Extract + PEPTONE + CS | 1:2:2 |
| 6 | Yeast Extract + CDHP + Glucose | 1:1:2 |
| 7 | CT + Peptone + Glucose | 2:2:2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivapragasam, M.; Wilfred, C.D.; Jaganathan, J.R.; Krishnan, S.; Ab Wan Karim Ghani, W.A.W. Choline-Based Ionic Liquids as Media for the Growth of Saccharomyces cerevisiae. Processes 2019, 7, 471. https://doi.org/10.3390/pr7070471
Sivapragasam M, Wilfred CD, Jaganathan JR, Krishnan S, Ab Wan Karim Ghani WAW. Choline-Based Ionic Liquids as Media for the Growth of Saccharomyces cerevisiae. Processes. 2019; 7(7):471. https://doi.org/10.3390/pr7070471
Chicago/Turabian StyleSivapragasam, Magaret, Cecilia Devi Wilfred, Joshua Raj Jaganathan, Sooridarsan Krishnan, and Wan Azlina Wan Ab Wan Karim Ghani. 2019. "Choline-Based Ionic Liquids as Media for the Growth of Saccharomyces cerevisiae" Processes 7, no. 7: 471. https://doi.org/10.3390/pr7070471