Lipid Isolation Process and Study on Some Molecular Species of Polar Lipid Isolated from Seed of Madhuca ellitica

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of Total Lipids (TL)
2.3. Lipid Fractionation
2.4. Analysis of Lipid Class Compositions
2.5. Analysis of Lipid Class Compositions
2.6. Analysis of Molecular Species of Polar Lipids
3. Results
3.1. Total Lipid Composition
3.2. Fatty Acid Composition of Total Lipids and Lipid Fractions
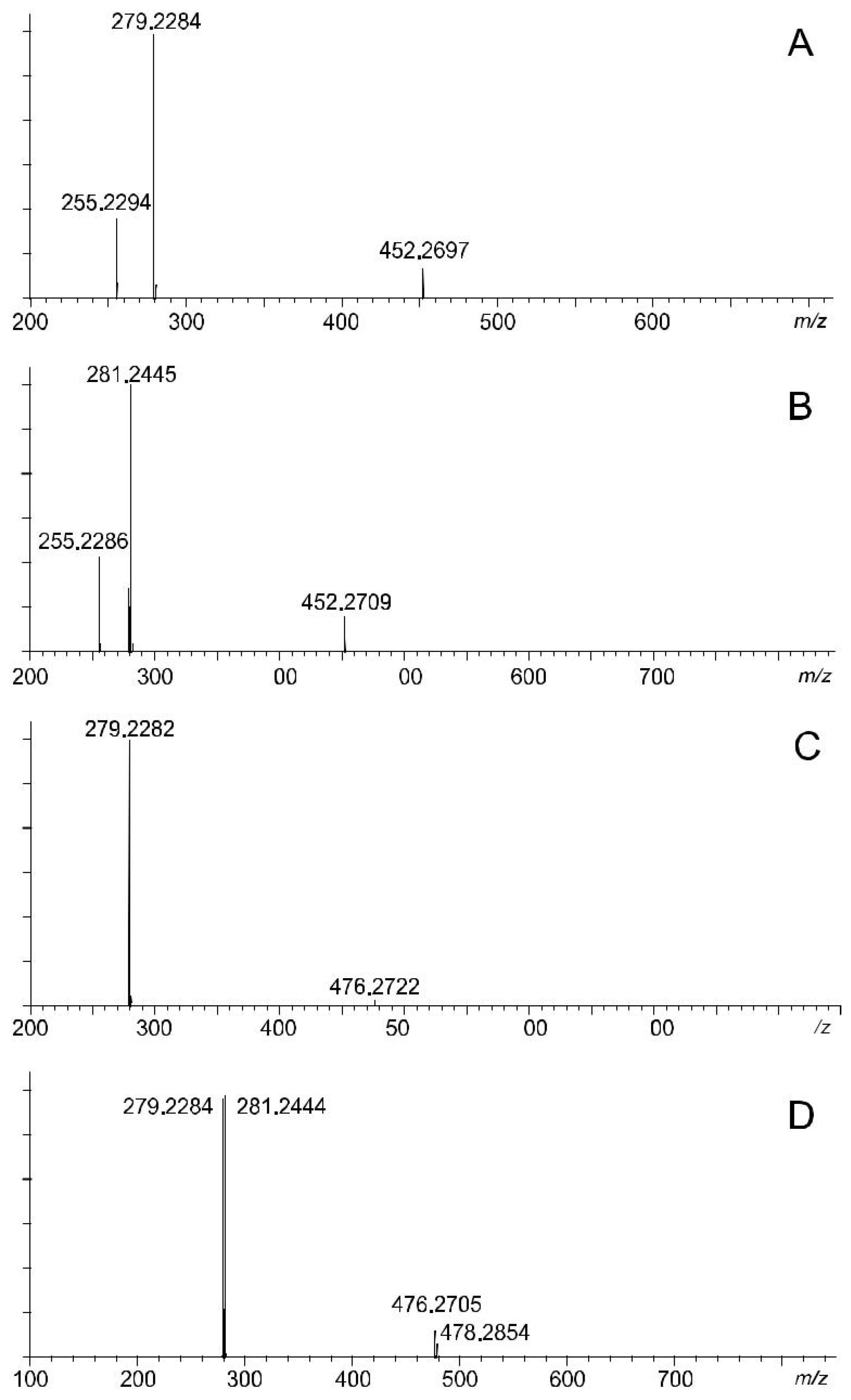
3.3. Chemical Structure of Molecular Species of Polar Lipids
3.4. Composition of the Molecular Species of the Phospholipids
3.5. Sulfoquinovosyldiacylglycerol (SQDG)
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kundu, M. Madhuca longifolia (Koenig) J. F. Macb. Seed Leaflets 2012, 156. [Google Scholar]
- Chopra, R.N.; Nayar, S.L.; Chopra, I.C. Glossary of Indian Medicinal Plant; National Institute of Science communication and Information resource (CSIR): New Delhi, India, 1985. [Google Scholar]
- Benerji, D.S.N.; Rajini, K.; Rao, B.S. Studies on physico-chemical and nutritional parameter for the production of ethanol from Mahua flower (madhuca indica) using Saccharomyces cerevisiae-3090 through submerged fermentation (smf). J. Microb. Biochem. Technol. 2010, 2, 46–50. [Google Scholar] [CrossRef]
- Siddiqui, S.B.; Khan, S.; Kardar, M.N. A New Isoflavone from the Madhuca latifolia. Nat. Prod. Res. 2010, 24, 76–80. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Tanaka, M.; Arihara, S.; Pal, B.C.; Roy, S.K.; Matsumura, E.; Katayama, S. New oleanene triterpenoid saponins from Madhuca longifolia. J. Nat. Prod. 2000, 63, 1679–1681. [Google Scholar] [CrossRef]
- Yosioka, I.; Inada, A.; Kitagawa, I. Soil bacterial hydrolysis leading to genuine aglycon. VIII. Structures of a genuine sapogenol protobassic acid and a prosapogenol of seed kernels of Madhuca longifoli. Tetrahedron 1974, 30, 707–714. [Google Scholar] [CrossRef]
- Siddiqui, S.B.; Khan, S.; Kardar, M.N.; Aslam, H. Chemical constituents from the fruits of Madhuca latifolia. Helventica Chim. Acta. 2004, 87, 1194–1201. [Google Scholar] [CrossRef]
- Ruckmini, C. Reproductive toxicology and nutritional studies on mahua oil (Madhuca latifolia). Food Chem. Toxicol. 1990, 28, 601–605. [Google Scholar] [CrossRef]
- Manorama, R.; Chinnasamy, N.; Rukmini, C. Multigeneration studies on red palm oil, and on hydrogenated vegetable oil containing mahua oil. Food Chem. Toxicol. 1993, 31, 369–375. [Google Scholar] [CrossRef]
- Ramadan, M.F.; Sharanabasappa, G.; Parmjyothi, S.; Seshagiri, M.; Moersel, J.T. Profile and levels of fatty acids and bioactive constituents in mahua butter from fruit-seeds of buttercup tree [Madhuca longifolia (Koenig)]. Eur. Food Res. Technol. 2006, 222, 710–718. [Google Scholar] [CrossRef]
- Singh, A.; Singh, I.S. Chemical evaluation of Mahua (Madhuca indica) seed. Food Chem. 1991, 40, 221–228. [Google Scholar] [CrossRef]
- Ayyanar, M.; Ignacimuthu, S. Traditional knowledge of kani tribals in Kouthalai of Tirunelveli hills, Tamil Nadu. India. J. Ethnopharmacol. 2005, 102, 246–255. [Google Scholar] [CrossRef]
- Yadav, S.; Suneja, P.; Hussain, Z.; Abraham, Z.; Mishra, S.K. Prospects and potential of Madhuca longifolia (Koenig) J.F. Macbride for nutritional and industrial purpose. Biomass Bioenergy 2011, 35, 1539–1544. [Google Scholar] [CrossRef]
- Parrota, J.A. Healing Plants of Peninsular India; CABI Publishing: London, UK, 2001. [Google Scholar]
- Gunstone, F.D. Phospholipid Technology and Applications; The Oily Press: Bridgwater, UK, 2008. [Google Scholar]
- Schwudke, D.; Schuhmann, K.; Herzog, R.; Bornstein, S.R.; Shevchenko, A. Shotgun lipidomics on high resolution mass spectrometers. Cold Spring Harb. Perspect. Biol. 2001, 3, a004614. [Google Scholar] [CrossRef]
- Frega, N.G.; Pacetti, D.; Boselli, E. Characterization of phospholipid molecular species by means of HPLC-tandem mass spectrometry. In Tandem Mass Spectrometry—Applications and Principles; Prasain, J., Ed.; InTech: London, UK, 2012; pp. 637–672. [Google Scholar]
- Gray, C.G.; Lasiter, A.D.; Li, C.; Leblond, J.D. Mono- and digalactosyldiacylglycerol composition of dinoflagellates. I. Peridinin-containing taxa. Eur. J. Phycol. 2009, 44, 191–197. [Google Scholar] [CrossRef]
- Kadri, N.; Khettal, B.; Yahiaoui-Zaidi, R.; Barragan-Montero, V.; Montero, J.L. Analysis of polar lipid fraction of Pinus halepensis Mill. seeds from North Algeria. Ind. Crops Prod. 2013, 51, 116–122. [Google Scholar] [CrossRef]
- Dodson, V.J.; Mouget, J.L.; Dahmen, J.L.; Leblond, J.D. The long and short of it: Temperature dependent modifications of fatty acid chain length and unsaturation in the galactolipid profiles of the diatoms Haslea ostrearia and Phaeodactylum tricornutum. Hydrobiologia 2014, 727, 95–107. [Google Scholar] [CrossRef]
- Marikkar, J.M.N.; Ghazali, H.M.; Long, K. Composition and thermal characteristics of Madhuca longifolia seed fat and its solid and liquid fractions. J. Oleo Sci. 2010, 59, 7–14. [Google Scholar] [CrossRef]
- Peterson, B.L.; Cummings, B.S. A review of chromatographic methods for the assessment of phospholipids in biological samples. Biomed. Chromatogr. 2006, 20, 227–243. [Google Scholar] [CrossRef]
- Imbs, A.B.; Dang, L.P.; Rybin, V.G.; Nguyen, N.T.; Pham, L.Q. Distribution of very-long-chain fatty acids between molecular species of different phospholipid classes of two soft corals. Biochem. Anal. Biochem. 2015, 4, 1–3. [Google Scholar]
- Carter, J.D.; Leblond, J.D. Red (hot) algae: Modulation of mono-and digalactosyldiacylglycerol-associated fatty acids of Polysiphonia sp. and Porphyridium sp. in response to growth temperature. Eur. J. Phycol. 2018, 53, 460–470. [Google Scholar] [CrossRef]
- Honda, M.; Ishimaru, T.; Itabashi, Y.; Vyssotski, M. Glycerolipid composition of the red macroalga Agarophyton Chilensis and comparison to the closely related Agarophyton vermiculophyllum producing different types of eicosanoids. Mar. Drugs 2019, 17, 96. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.F.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissue. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Hamoutene, D.; Puestow, T.; Miller-Banoub, J.; Wareham, V. Main lipid classes in some species of deep-sea corals in the Newfoundland and Labrador region (Northwest Atlantic Ocean). Coral Reefs 2008, 27, 237–246. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Kulikova, I.V. Lipids of different parts of the lamina of Laminaria japonica Aresch. Bot. Mar. 2000, 43, 87–91. [Google Scholar] [CrossRef]
- Vaskovsky, V.E.; Kostetsky, E.Y.; Vasendin, I.M. A universal reagent for phospholipids analysis. J. Chromatogr. 1974, 114, 129–141. [Google Scholar] [CrossRef]
- Svetashev, V.I. Mild method for preparation of 4,4-dimethyloxazoline derivatives of polyunsaturated fatty acids for GC–MS. Lipids. 2011, 46, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Stransky, K.; Jursik, T.; Vitek, A. Standard equivalent chain length values of monoenic and polyenic (methylene interrupted) fatty acids. J. High Resolut. Chromatogr. 1997, 20, 143–158. [Google Scholar] [CrossRef]
- The AOCS Lipid Library—Mass Spectrometry of Fatty Acid Derivatives. Available online: http://lipidlibrary.aocs.org/ms/masspec.html (accessed on 16 August 2014).
- Postle, A.D.; Wilton, D.C.; Hunt, A.N.; Attard, G.S. Probing phospholipid dynamics by electrospray ionisation mass spectrometry. Prog. Lipid Res. 2007, 46, 200–224. [Google Scholar] [CrossRef]
- Imbs, A.B.; Dang, L.P.T.; Rybin, V.G.; Svetashev, V.I. Fatty acid, lipid class, and phospholipid molecular species composition of the soft coral Xenia sp. (Nha Trang Bay, the South China Sea, Vietnam). Lipids 2015, 60, 575–589. [Google Scholar] [CrossRef]
- Mohamed, A.; Bhardwaj, H.; Hamama, A.; Webber, C.L., III. Chemical composition of kenaf (Hibiscus cannabinus L.) seed oil. Ind. Corp. Prod. 1995, 4, 157–165. [Google Scholar] [CrossRef]
- Dowhan, W. Molecular basis for membrane phospholipid diversity: Why are there so many lipids? Annu. Rev. Biochem. 1997, 66, 199–232. [Google Scholar] [CrossRef] [PubMed]
- Postle, A.D. Phospholipid lipidomics in health and disease. Eur. J. Lipid Sci. Tech. 2009, 111, 2–13. [Google Scholar] [CrossRef]
- Hermansson, M.; Hokynar, K.; Somerharju, P. Mechanisms of glycerophospholipid homeostasis in mammalian cells. Prog. Lipid Res. 2011, 50, 240–257. [Google Scholar] [CrossRef]
- Astudillo, A.M.; Balgoma, D.; Balboa, M.A.; Balsinde, J. Dynamics of arachidonic acid mobilization by inflammatory cells. Biochim. Biophys. Acta 2012, 1821, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacetti, D.; Boselli, E.; Lucci, P.; Frega, N.G. Simultaneous analysis of glycolipids and phospholids molecular species in avocado (Persea americana Mill.) fruit. J. Chromatogr. A 2007, 1150, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Postle, A.D. Lipidomics. Curr. Opin. Clin. Nut. Metab. Care. 2012, 15, 127–133. [Google Scholar] [CrossRef]
- Brouwers, J.F. Liquid chromatographic-mass spectrometric analysis of phospholipids. Chromatography, ionization and quantification. Biochim. Biophys. Acta 2011, 1811, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.S.; Shiva, S.; Roth, M.R.; Tamura, P.; Zheng, L.Q.; Sarowar, S.; Li, M.; Honey, S.; McEllhiney, D.; Hinkes, P.; et al. Lipid changes after leaf wounding in Arabidopsis thaliana: Expanded lipidomic data form the basis for lipid co-occurrence analysis. Plant J. 2014, 80, 728–743. [Google Scholar] [CrossRef] [PubMed]
- Schmid, K.M.; Ohlrogge, J.B. Lipid metabolism in plants. In New Comprehensive Biochemistry, Biochemistry of Lipids, Lipoproteins and Membranes, 4th ed.; Vance, D.E., Vance, J.E., Eds.; Elsevier: New York, NY, USA, 2002; Volume 36, pp. 93–126. [Google Scholar]
- Frentzen, M. Phosphatidylglycerol and sulfoquinovosyldiacylglycerol: Anionic membrane lipids and phosphate regulation. Curr. Opin. Plant. Biol. 2004, 7, 270–276. [Google Scholar] [CrossRef]
- Basconcillo, L.S.; Zaheer, R.; Finan, T.M.; McCarry, B.E. A shotgun lipidomics approach in Sinorhizobium meliloti as a tool in functional genomics. J. Lipid. Res. 2009, 50, 1120–1132. [Google Scholar] [CrossRef]
- Kim, Y.H.; Yoo, J.S.; Kim, M.S. Structural characterization of sulfoquinovosyl, monogalactosyl and digalactosyl diacylglycerols by FAB-CID-MS/MS. J. Mass Spect. 1997, 32, 968–977. [Google Scholar] [CrossRef]
- Xue, C.H.; Hu, Y.Q.; Saito, H.; Zhang, Z.H.; Li, Z.J.; Cai, Y.P.; Ou, C.R.; Lin, H.; Imbs, A.B. Molecular species composition of glycolipids from Sprirulina platensis. Food Chem. 2002, 77, 9–13. [Google Scholar] [CrossRef]
- Li, H.Y.; Yan, X.J.; Xu, J.L.; Zhou, C.X. Precise identification of photosynthetic glycerolipids in microalga Tetraselmis chuii by UPLC-ESI-Q-TOF-MS. Sci. China. 2008, 51, 1101–1107. [Google Scholar] [CrossRef]
- Plouguerné, E.; da Gama, B.A.P.; Pereira, R.C.; Barreto-Bergter, E. Glycolipids from seaweeds and their potential biotechnological applications. Front. Cell Infect. Microbiol. 2014, 4, 174. [Google Scholar] [CrossRef] [PubMed]
- Melo, T.; Alves, E.; Azevedo, V.; Martins, A.S.; Neves, B.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomics as a new approach for the bioprospecting of marine macroalgae—Unraveling the polar lipid and fatty acid composition of Chondrus crispus. Algal Res. 2015, 8, 181–191. [Google Scholar] [CrossRef]
- Logvinov, S.V.; Denisenko, V.A.; Dmitrenok, P.S.; Moiseenko, O.P. Sulfoquinovosyldiacylglycerines from Scaphechinus mirabilis. Chem. Nat. Comp. 2012, 48, 175–179. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Lipid Class | Content (%) | Phospholipid (PL) Class | Content (%) |
|---|---|---|---|
| Waxes | 5.28 ± 0.1 | PC | 30.7 ± 0.4 |
| Triacylglycerols | 63.2 ± 1.5 | PE | 29.6 ± 0.3 |
| Diacylglycerols | 5.17 ± 0.9 | PI | 17.2 ± 0.1 |
| Free fatty acids | 6.14 ± 0.6 | PG | 10.9 ± 0.2 |
| Glycolipids | 5.78 ± 0.2 | PA | 6.8 ± 0.1 |
| Phospholipids | 14.43 ± 1.3 | LPE | 4.8± 0.1 |
| Fatty Acids | Total Lipids | Neutral Lipids | Polar Lipids |
|---|---|---|---|
| 14:0 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.1 ± 0.1 |
| 16:0 | 24.4 ± 1.2 | 25.7 ± 1.4 | 28.5 ± 1.7 |
| 16:1n-7 | 0.1 ± 0.1 | 0.1 ± 0.1 | nd |
| 18:0 | 11.5 ± 0.5 | 13.5 ± 2.3 | 12.5 ± 1.8 |
| 18:1n-9 | 49.5 ± 3.5 | 46.1 ± 2.8 | 44.8 ± 2.9 |
| 18:2n-6 | 13.6 ± 1.1 | 13.6 ± 0.9 | 13.2 ± 1.3 |
| 18:3n-3 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.3 ± 0.1 |
| 20:0 | 0.4 ± 0.2 | 0.4 ± 0.2 | 0.3 ± 0.1 |
| Σ SFA | 36.4 ± 0.4 | 39.8 ± 0.6 | 41.4 ± 0.5 |
| Σ UFA | 63.3 ± 1.7 | 60.0 ± 1.1 | 58.3 ± 1.3 |
| SFA/UFA | 0.58 | 0.66 | 0.71 |
| No. | Molecular Species | ESI-MS | Monoisotopic Molecular Mass | Molecular Formula | MS2 | ||||
|---|---|---|---|---|---|---|---|---|---|
| [M + H + Et3N]+ m/z | [M − H]– m/z | Measured | Calculated | Fragment Ion [M − H − X]– * | X | ||||
| m/z | Composition | ||||||||
| 1 | 16:0/18:2 PE | 817.6365 | 714.4988 | 715.5061 | 715.5152 | C39H74NO8P | 452.2697 | C21H44NO7P | C18H30O |
| 279.2284 | C18H31O2 | C21H43NO6P | |||||||
| 255.2294 | C16H31O2 | C23H43NO6P | |||||||
| 2 | 16:0/18:1 PE | 819.6499 | 716.5163 | 717.5236 | 717.5309 | C39H76NO8P | 452.2713 | C21H44NO7P | C18H32O |
| 281.2443 | C18H33O2 | C21H43NO6P | |||||||
| 255.2294 | C16H31O2 | C23H45NO6P | |||||||
| 3 | 18:2/18:2 PE | 841.6331 | 738.4975 | 739.5048 | 739.5152 | C41H74NO8P | 476.2778 | C23H44NO7P | C18H30O |
| 279.2283 | C18H31O2 | C23H43NO6P | |||||||
| 4 | 18:1/18:2 PE | 843.6484 | 740.5140 | 741.5213 | 741.5309 | C41H76NO8P | 478.2854 | C23H46NO7P | C18H30O |
| 476.2705 | C23H44NO7P | C18H32O | |||||||
| 281.2444 | C18H33O2 | C23H43NO6P | |||||||
| 279.2284 | C18H31O2 | C23H45NO6P | |||||||
| 5 | 18:1/18:1 PE (4.4) 18:0/18:2 PE (1) | 845.6627 | 742.5296 | 743.5369 | 743.5465 | C41H78NO8P | 478.2866 | C23H46NO7P | C18H32O |
| 476.2707 | C23H44NO7P | C18H34O | |||||||
| 460.2808 | C23H44NO6P | C18H34O2 | |||||||
| 283.2545 | C18H35O2 | C23H43NO6P | |||||||
| 281.2443 | C18H33O2 | C23H45NO6P | |||||||
| 279.2285 | C18H31O2 | C23H47NO6P | |||||||
| 6 | 18:0/18:1 PE | 847.6749 | 744.5436 | 745.5509 | 745.5622 | C41H80NO8P | 480.3046 | C23H48NO7P | C18H32O |
| 283.2577 | C18H35O2 | C23H45NO6P | |||||||
| 281.2441 | C18H33O2 | C23H47NO6P | |||||||
| No. | Molecular Species | ESI-MS | Monoisotopic Molecular Mass | Molecular Formula | MS2 | MS3 | ||
|---|---|---|---|---|---|---|---|---|
| [M + CH3COO]− m/z | Measured | Calculated | Fragment Ion * [M + CH3COO − C3H6O2]− m/z | Fragmentation ** | ||||
| m/z | Composition | |||||||
| 1 | 16:0/18:2 PC | 816.5760 | 757.5621 | 757.5622 | C42H80NO8P | 742.5389 | 279.2274 | C18H31O2 |
| 2 | 16:0/18:1 PC | 818.5889 | 759.5750 | 759.5778 | C42H82NO8P | 744.5532 | 281.2462 | C18H33O2 |
| 255.2320 | C16H31O2 | |||||||
| 3 | 16:0/18:0 PC | 820.5943 | 761.5804 | 761.5935 | C42H84NO8P | 746.5599 | ND | - |
| 4 | 18:2/18:2 PC | 840.5712 | 781.5573 | 781.5622 | C44H80NO8P | 766.5309 | ND | - |
| 5 | 18:1/18:2 PC | 842.5888 | 783.5749 | 783.5778 | C44H82NO8P | 768.5511 | 281.2432 | C18H33O2 |
| 279.2352 | C18H31O2 | |||||||
| 6 | 18:1/18:1 PC (1.58) 18:0/18:2 PC (1) | 844.6059 | 785.5920 | 785.5935 | C44H84NO8P | 770.5674 | 283.2602 | C18H35O2 |
| 281.2474 | C18H33O2 | |||||||
| 279.2332 | C18H31O2 | |||||||
| No. | Molecular Species | ESI-MS | Monoisotopic Molecular Mass | Molecular Formula | MS2 | ||
|---|---|---|---|---|---|---|---|
| [M − H]– m/z | Measured | Calculated | Fragment Ion * [M − H − X]− m/z | X | |||
| 1 | 16:1/18:1 PI 16:0/18:2 PI | 833.5188 | 834.5256 | 834.5258 | C43H79O13P | 595.2769 | C16H30O |
| 579.2901 | C16H30O2 | ||||||
| 577.2784 | C16H32O2 | ||||||
| 571.2904 | C18H30O | ||||||
| 553.2813 | C18H30O2 | ||||||
| 417.2418 | C22H40O7 (C6H10O5 + C16H30O2) | ||||||
| 415.2268 | C22H42O7 (C6H10O5 + C16H32O2) | ||||||
| 409.2352 | C24H40O6 (C6H10O5 + C18H30O) | ||||||
| 391.2253 | C24H42O7 (C6H10O5 + C18H32O2) | ||||||
| 297.0388 | C34H64O4 (C16H32O2 + C18H32O2) | ||||||
| 281.2474 | C25H45O11P | ||||||
| 279.2307 | C25H47O11P | ||||||
| 255.2346 | C27H47O11P | ||||||
| 2 | 16:0/18:1 PI | 835.5328 | 836.5401 | 836.5415 | C43H81O13P | 597.3036 | C16H30O |
| 579.2939 | C16H32O2 | ||||||
| 571.2876 | C18H32O | ||||||
| 553.2805 | C18H34O2 | ||||||
| 435.2438 | C22H40O6 (C6H10O5 + C16H30O) | ||||||
| 417.2389 | C22H42O7 (C6H10O5 + C16H32O2) | ||||||
| 409.2351 | C24H42O6 (C6H10O5 + C18H32O) | ||||||
| 391.2250 | C24H44O7 (C6H10O5 + C18H34O2) | ||||||
| 297.0373 | C34H64O4 (C16H32O2 + C18H34O2) | ||||||
| 281.2484 | C25H47O11P | ||||||
| 255.2337 | C27H49O11P | ||||||
| 3 | 18:2/18:2 PI | 857.5155 | 858.5228 | 858.5258 | C45H79O13P | 577.2805 | C18H32O2 |
| 415.2264 | C24H42O7 (C6H10O5 + C18H32O2) | ||||||
| 279.2337 | C27H47O11P | ||||||
| 4 | 18:0/18:2 PI 18:1/18:1 PI | 861.5463 | 862.5536 | 862.5571 | C45H81O13P | 599.3202 | C18H30O |
| 597.3037 | C18H32O | ||||||
| 595.2894 | C18H34O | ||||||
| 581.3095 | C18H32O2 | ||||||
| 579.2945 | C18H34O2 | ||||||
| 577.2775 | C18H36O2 | ||||||
| 437.2665 | C24H40O6 (C6H10O5 + C18H30O) | ||||||
| 435.2511 | C24H42O6 (C6H10O5 + C18H32O) | ||||||
| 433.2306 | C24H44O6 (C6H10O5 + C18H34O) | ||||||
| 419.2571 | C24H42O7 (C6H10O5 + C18H32O2) | ||||||
| 417.2394 | C24H44O7 (C6H10O5 + C18H34O2) | ||||||
| 415.2241 | C24H46O7 (C6H10O5 + C18H36O2) | ||||||
| 297.0370 | C36H68O4 | ||||||
| 283.2622 | C27H45O11P | ||||||
| 281.2484 | C27H47O11P | ||||||
| 279.2328 | C27H49O11P | ||||||
| 5 | 18:0/18:1 PI | 863.5635 | 864.5708 | 864.5728 | C45H83O13P | 701.5027 | C6H10O5 |
| 599.3203 | C18H32O | ||||||
| 597.3032 | C18H34O | ||||||
| 581.3097 | C18H34O2 | ||||||
| 579.2943 | C18H36O2 | ||||||
| 419.2573 | C24H44O7 (C6H10O5 + C18H34O2) | ||||||
| 417.2393 | C24H46O7 (C6H10O5 + C18H36O2) | ||||||
| 297.0376 | C36H70O4 | ||||||
| 283.2627 | C27H47O11P | ||||||
| 281.2485 | C27H49O11P | ||||||
| No. | Molecular Species | ESI-MS | Monoisotopic Molecular Mass | Molecular Formula | MS2 | ||
|---|---|---|---|---|---|---|---|
| [M − H]– m/z | Measured | Calculated | Fragment Ion * [M − H − X]− m/z | X | |||
| 1 | 16:0/18:2 PG | 745.4944 | 746.5017 | 746.5098 | C40H75O10P | 391.2224 | C21H38O4 (C3H6O2 + C18H32O2) |
| 279.2303 | C22H43O8P | ||||||
| 255.2336 | C24H43O8P | ||||||
| 2 | 16:0/18:1 PG | 747.5092 | 748.5164 | 748.5254 | C40H77O10P | 281.2482 | C22H43O8P |
| 255.2323 | C24H45O8P | ||||||
| 3 | 16:0/18:0 PG | 749.5212 | 750.5285 | 750.5411 | C40H79O10P | 283.2598 | C22H43O8P |
| 255.2321 | C24H47O8P | ||||||
| 4 | 18:1/18:2 PG | 771.5092 | 772.5164 | 772.5254 | C42H77O10P | 491.2788 | C18H32O2 |
| 281.2477 | C24H43O8P | ||||||
| 279.2325 | C24H45O8P | ||||||
| 5 | 18:1/18:1 PG 18:0/18:2 PG | 773.5253 | 774.5326 | 774.5411 | C42H79O10P | 283.2598 | C24H43O8P |
| 281.2478 | C24H45O8P | ||||||
| 279.2328 | C24H47O8P | ||||||
| 6 | 18:0/18:1 PG | 775.5380 | 776.5524 | 776.5567 | C42H81O10P | 493.2940 | C18H34O2 |
| 491.2773 | C18H36O2 | ||||||
| 419.2554 | C21H40O4 (C3H6O2 + C18H34O2) | ||||||
| 417.2406 | C21H42O4 (C3H6O2 + C18H36O2) | ||||||
| 283.2608 | C24H45O8P | ||||||
| 281.2478 | C24H47O8P | ||||||
| No. | Molecular Species | ESI-MS | Monoisotopic Molecular Mass | Molecular Formula | MS2 | ||
|---|---|---|---|---|---|---|---|
| [M − H]– m/z | Measured | Calculated | Fragment ion * [M − H − X]− m/z | X | |||
| 1 | 16:0/18:2 PA | 671.4643 | 672.4716 | 672.4730 | C37H69O8P | 409.2375 | C18H30O |
| 391.2271 | C18H32O2 | ||||||
| 279.2274 | C19H37O6P | ||||||
| 255.2313 | C21H37O6P | ||||||
| 2 | 16:0/18:1 PA | 673.4798 | 674.4871 | 674.4887 | C37H71O8P | 417.2394 | C16H32O2 |
| 409.2332 | C18H32O | ||||||
| 391.2242 | C18H34O2 | ||||||
| 281.2469 | C19H37O6P | ||||||
| 255.2318 | C21H39O6P | ||||||
| 3 | 18:2/18:2 PA | 695.4660 | 696.4727 | 696.4730 | C39H69O8P | 433.2324 | C18H30O |
| 415.2228 | C18H32O2 | ||||||
| 279.2319 | C21H37O6P | ||||||
| 4 | 18:1/18:2 PA | 697.4803 | 698.4876 | 698.4887 | C39H71O8P | 435.2511 | C18H30O |
| 433.2351 | C18H32O | ||||||
| 417.2405 | C18H32O2 | ||||||
| 415.2241 | C18H34O2 | ||||||
| 281.2458 | C21H37O6P | ||||||
| 279.2316 | C21H39O6P | ||||||
| 5 | 18:0/18:2 PA 18:1/18:1 PA | 699.4949 | 700.5022 | 700.5043 | C39H73O8P | 437.2641 | C18H30O |
| 435.2511 | C18H32O | ||||||
| 419.256 | C18H32O2 | ||||||
| 417.2382 | C18H34O2 | ||||||
| 415.2223 | C18H36O2 | ||||||
| 283.2608 | C21H37O6P | ||||||
| 281.2479 | C21H39O6P | ||||||
| 279.2322 | C21H41O6P | ||||||
| 6 | 18:0/18:1 PA | 701.5098 | 702.5171 | 702.5200 | C39H75O8P | 437.2633 | C18H32O |
| 435.2503 | C18H34O | ||||||
| 419.2546 | C18H34O2 | ||||||
| 417.2385 | C18H36O2 | ||||||
| 283.2621 | C21H39O6P | ||||||
| 281.247 | C21H41O6P | ||||||
| Phospholipid Class | Molecular Species | Concentration | ||
|---|---|---|---|---|
| % of Each Phospholipid Class | % of Total Phospholipids | mg/kg of Total Lipids | ||
| PE | 16:0/18:1 PE | 19.21 | 5.77 | 2567 |
| 16:0/18:2 PE | 16.13 | 4.85 | 2158 | |
| 18:0/18:1 PE | 4.64 | 1.39 | 620 | |
| 18:0/18:2 PE | 3.96 | 1.19 | 529 | |
| 18:1/18:1 PE | 17.43 | 5.23 | 2329 | |
| 18:1/18:2 PE | 27.18 | 8.16 | 3632 | |
| 18:2/18:2 PE | 11.44 | 3.44 | 1529 | |
| PC | 16:0/18:0 PC | 3.96 | 1.19 | 530 |
| 16:0/18:1 PC | 30.11 | 9.05 | 4026 | |
| 16:0/18:2 PC | 13.27 | 3.99 | 1774 | |
| 18:0/18:2 PC | 17.10 | 5.14 | 2287 | |
| 18:1/18:1 PC | 10.79 | 3.24 | 1443 | |
| 18:1/18:2 PC | 19.78 | 5.94 | 2645 | |
| 18:2/18:2 PC | 4.98 | 1.50 | 666 | |
| PI | 16:0/18:1 PI | 44.18 | 7.70 | 3427 |
| 16:0/18:2 PI | 1.01 | 0.18 | 78 | |
| 16:1/18:1 PI | 20.92 | 3.65 | 1623 | |
| 18:0/18:2 PI | 23.08 | 4.02 | 1790 | |
| 18:1/18:1 PI | 5.25 | 0.92 | 407 | |
| 18:2/18:2 PI | 2.02 | 0.35 | 157 | |
| Other PI | 3.54 | 0.62 | 275 | |
| PG | 16:0/18:0 PG | 9.42 | 1.06 | 469 |
| 16:0/18:1 PG | 43.47 | 4.87 | 2167 | |
| 16:0/18:2 PG | 22.05 | 2.47 | 1099 | |
| 18:0/18:1 PG | 7.93 | 0.89 | 395 | |
| 18:0/18:2 PG | 2.77 | 0.31 | 138 | |
| 18:1/18:1 PG | 8.32 | 0.93 | 415 | |
| 18:1/18:2 PG | 5.14 | 0.58 | 256 | |
| Other PG | 0.90 | 0.10 | 45 | |
| PA | 16:0/18:1 PA | 29.01 | 1.91 | 852 |
| 16:0/18:2 PA | 15.98 | 1.05 | 469 | |
| 18:0/18:1 PA | 7.05 | 0.47 | 207 | |
| 18:0/18:2 PA | 4.74 | 0.31 | 139 | |
| 18:1/18:1 PA | 18.19 | 1.20 | 534 | |
| 18:1/18:2 PA | 19.62 | 1.29 | 576 | |
| 18:2/18:2 PA | 5.41 | 0.36 | 159 | |
| No. | Molecular Species | Content, mol. % | ESI–MS | Monoisotopic Molecular Mass | Molecular Formula | MS2 | ||
|---|---|---|---|---|---|---|---|---|
| Measured | Calculated | Fragment Ion a [M − H − X]− m/z | X | |||||
| 1 | 14:0/16:0 | 0.36 | 765.4858 | 766.4901 | 672.4730 | C39H74O12S | 537.2731 | C14H28O2 |
| 509.2461 | C16H32O2 | |||||||
| 2 | 16:0/16:0 | 7.15 | 793.5157 | 794.5198 | 794.5214 | C41H78O12S | 537.2723 | C16H32O2 |
| 3 | 16:0/18:3 | 2.04 | 815.4939 | 816.5011 | 816.5057 | C43H76O12S | 559.2593 | C16H32O2 |
| 4 | 16:0/18:2 + | 12.65 | 817.5106 | 818.5179 | 818.5214 | C43H78O12S | 563.2875 | C16H30O2 |
| 16:1/18:1 | 4.72 | 561.2734 | C16H32O2 | |||||
| 537.2730 | C18H32O2 | |||||||
| 5 | 16:0/18:1 | 28.09 | 819.5253 | 820.5325 | 820.537 | C43H80O12S | 563.2892 | C16H32O2 |
| 537.2747 | C18H34O2 | |||||||
| 6 | 18:2/18:3 | 0.77 | 839.4893 | 840.4965 | 840.5057 | C45H76O12S | 561.2694 | C18H30O2 |
| 559.2581 | C18H32O2 | |||||||
| 7 | 18:2/18:2 + | 2.44 | 841.5061 | 842.5134 | 842.5214 | C45H78O12S | 563.2841 | C18H30O2 |
| 18:1/18:3 | 0.75 | 561.2733 | C18H32O2 | |||||
| 559.2638 | C18H34O2 | |||||||
| 8 | 18:0/18:3 + | 0.99 | 843.5215 | 844.5287 | 844.537 | C45H80O12S | 565.3002 | C18H30O2 |
| 18:1/18:2 | 5.76 | 563.2883 | C18H32O2 | |||||
| 561.2728 | C18H34O2 | |||||||
| 559.2573 | C18H36O2 | |||||||
| 9 | 18:0/18:2 + | 7.12 | 845.5368 | 846.5441 | 846.5527 | C45H82O12S | 565.3005 | C18H32O2 |
| 18:1/18:1 | 8.75 | 563.2894 | C18H34O2 | |||||
| 561.2738 | C18H36O2 | |||||||
| 10 | 18:0/18:1 | 17.97 | 847.5527 | 848.5599 | 848.5683 | C45H84O12S | 565.3009 | C18H34O2 |
| 563.2903 | C18H36O2 | |||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phuong, D.L.; Toan, T.Q.; Dang, L.P.T.; Imbs, A.B.; Long, P.Q.; Thang, T.D.; Matthaeus, B.; Bach, L.G.; Bui, L.M. Lipid Isolation Process and Study on Some Molecular Species of Polar Lipid Isolated from Seed of Madhuca ellitica. Processes 2019, 7, 375. https://doi.org/10.3390/pr7060375
Phuong DL, Toan TQ, Dang LPT, Imbs AB, Long PQ, Thang TD, Matthaeus B, Bach LG, Bui LM. Lipid Isolation Process and Study on Some Molecular Species of Polar Lipid Isolated from Seed of Madhuca ellitica. Processes. 2019; 7(6):375. https://doi.org/10.3390/pr7060375
Chicago/Turabian StylePhuong, Doan Lan, Tran Quoc Toan, Ly P. T. Dang, Andrey B. Imbs, Pham Quoc Long, Tran Dinh Thang, Bertrand Matthaeus, Long Giang Bach, and Le Minh Bui. 2019. "Lipid Isolation Process and Study on Some Molecular Species of Polar Lipid Isolated from Seed of Madhuca ellitica" Processes 7, no. 6: 375. https://doi.org/10.3390/pr7060375