
Alternative Electron Sources for Cytochrome P450s Catalytic Cycle: Biosensing and Biosynthetic Application
 , ,
, ,
Abstract
:1. Introduction
2. Active Metals as Electron Donor for the Reduction in Heme Iron of CYPs
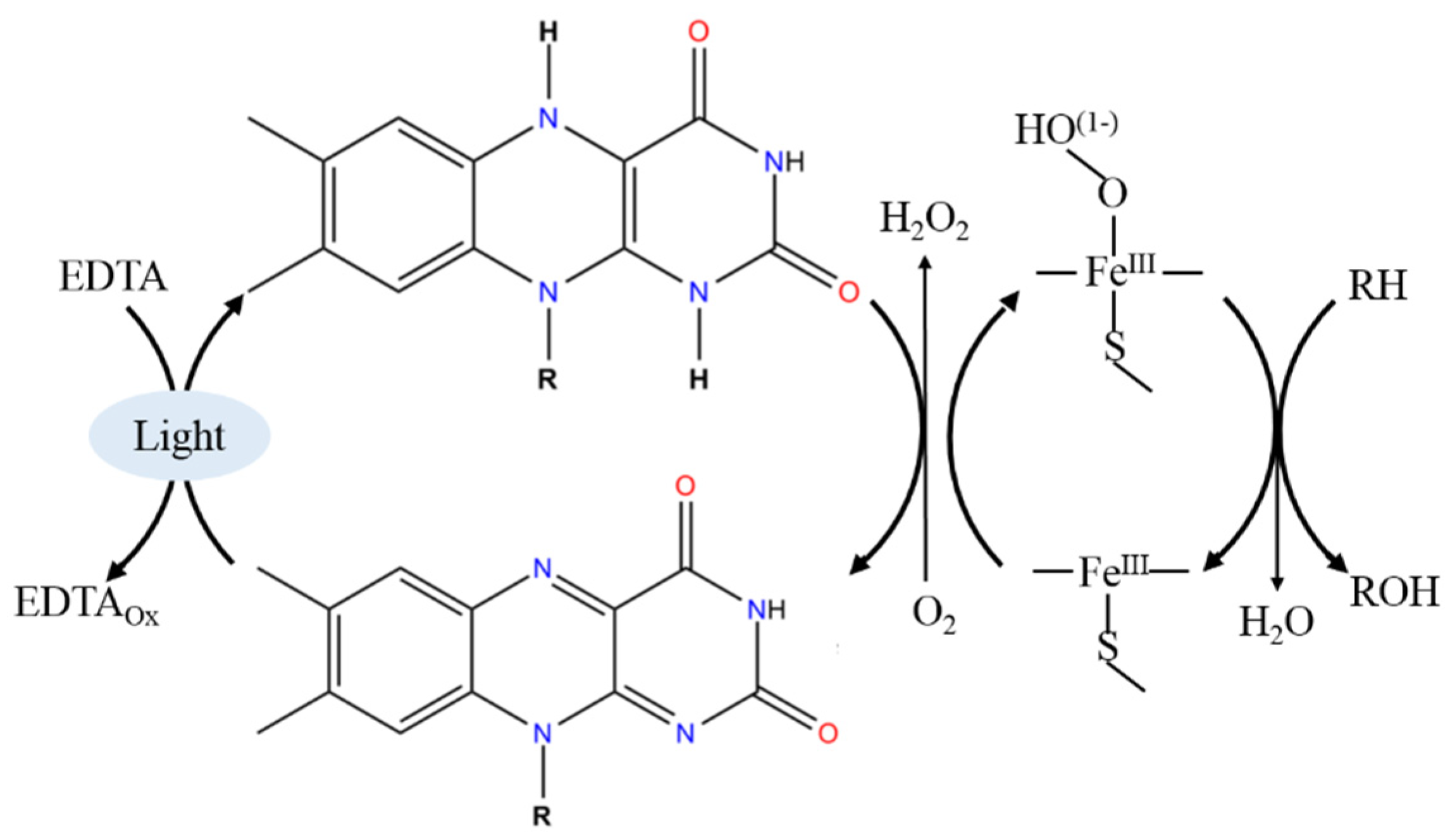
3. Light-Driven CYP Catalysis
4. Electrochemical Technology for Effective CYP Catalysis
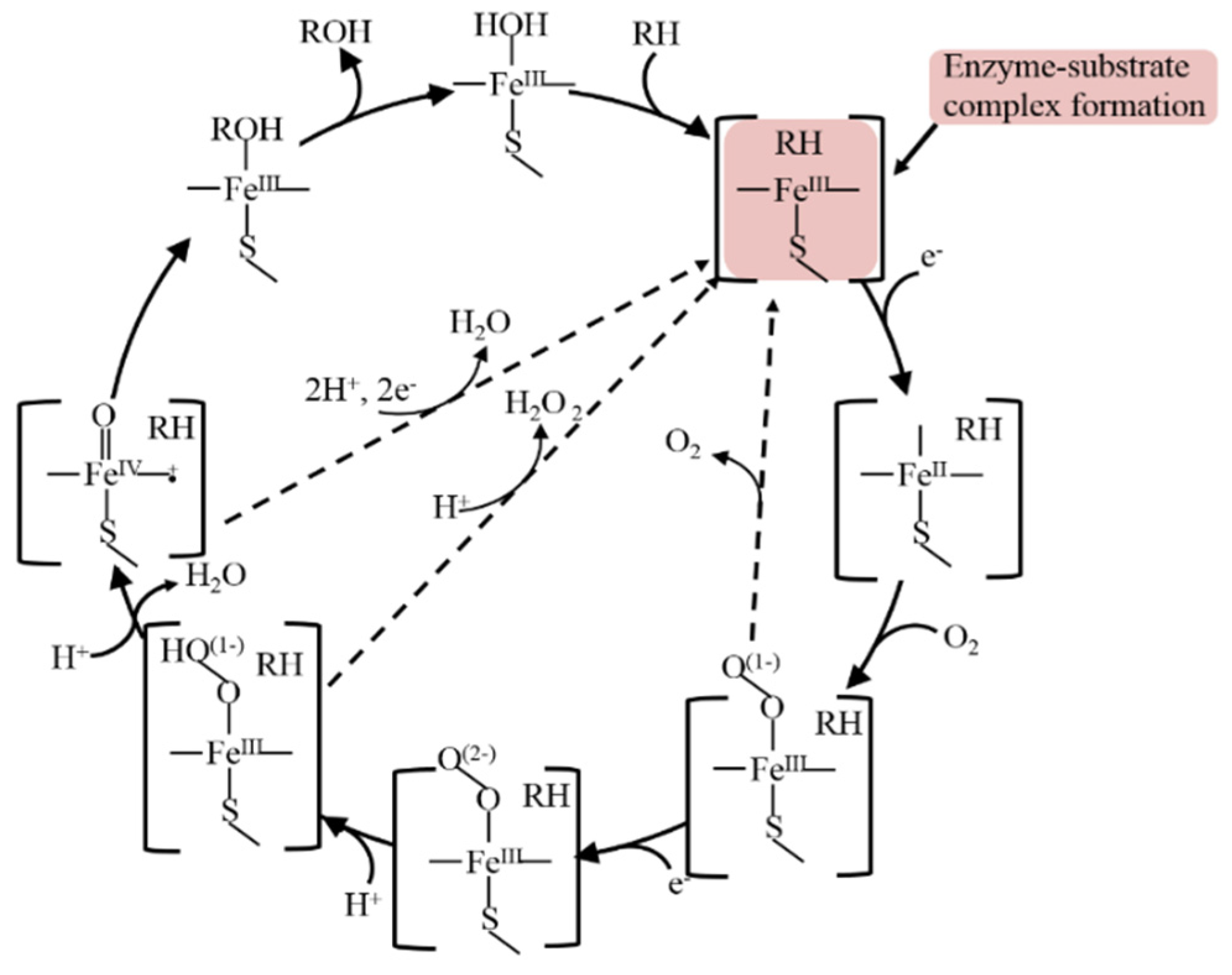
4.1. Modelling on Electrode the Catalytic Cycle of CYP3A4
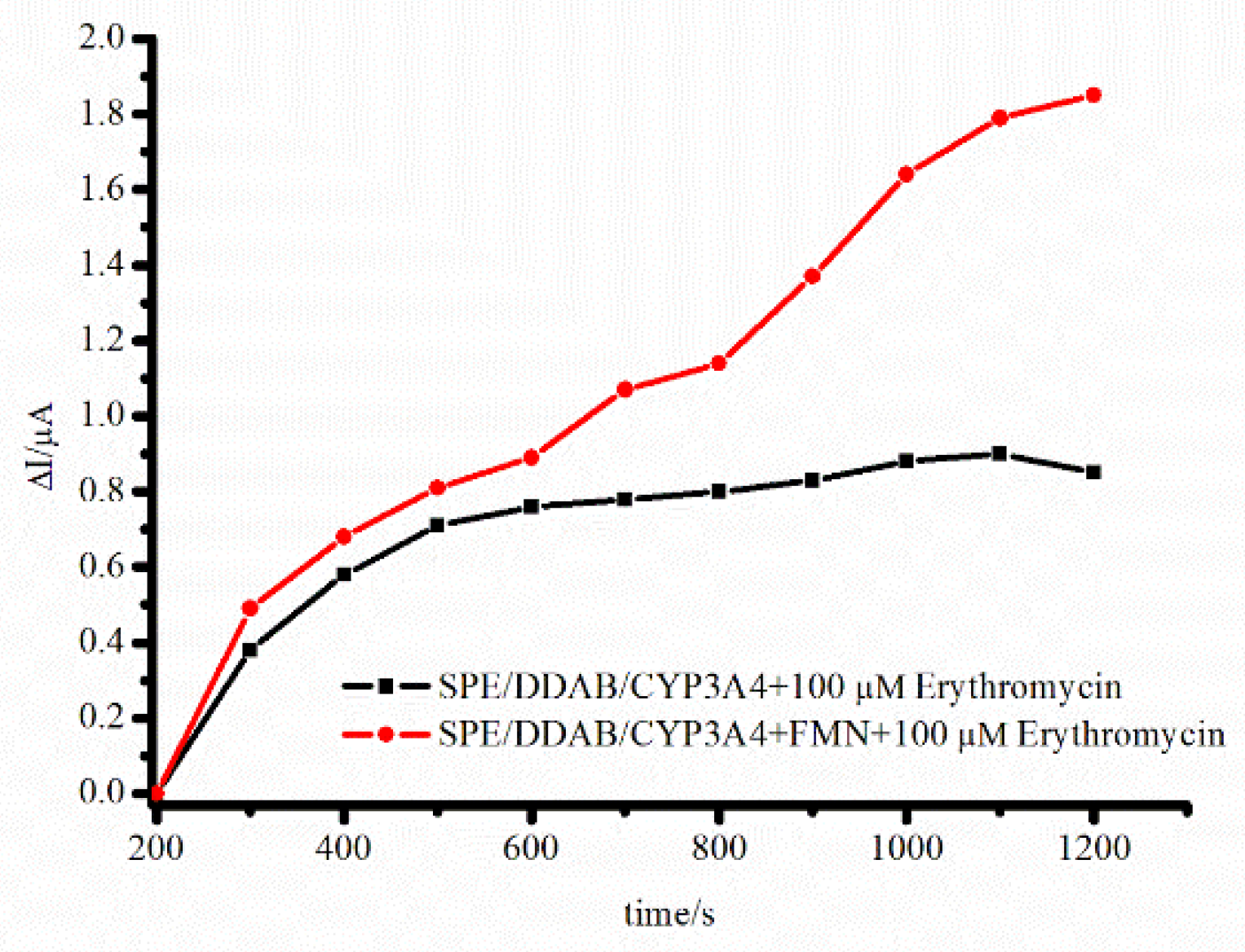
4.2. Electron Transfer Chain Optimization on CYP-Electrode
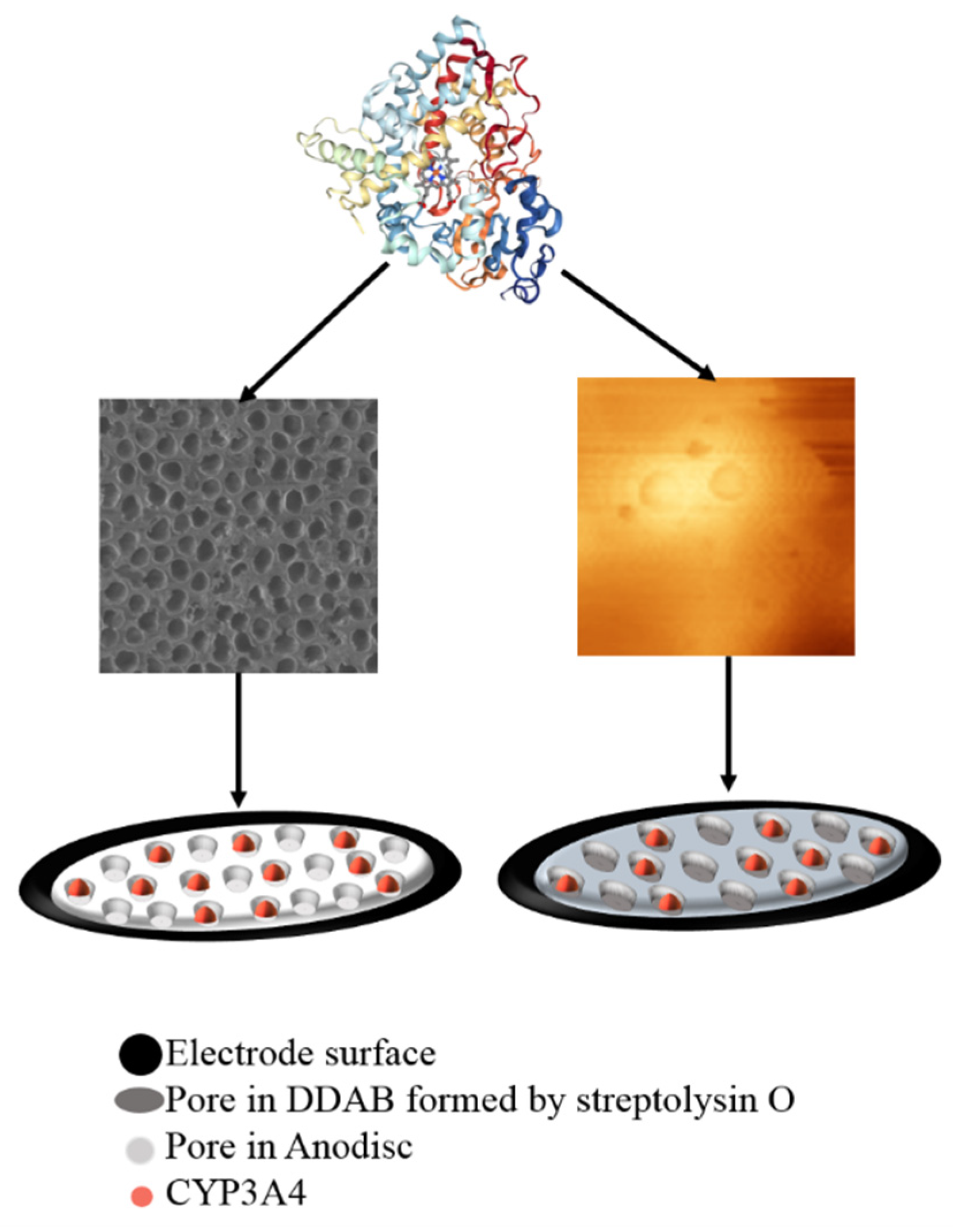
4.3. Modification of Electrode Surface for the 2D → 3D Transition
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bernhardt, R. Cytochromes P450 as Versatile Biocatalysts. J. Biotechnol. 2006, 124, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, R.; Urlacher, V.B. Cytochromes P450 as Promising Catalysts for Biotechnological Application: Chances and Limitations. Appl. Microbiol. Biotechnol. 2014, 98, 6185–6203. [Google Scholar] [CrossRef] [PubMed]
- Urlacher, V.B.; Girhard, M. Cytochrome P450 Monooxygenases in Biotechnology and Synthetic Biology. Trends Biotechnol. 2019, 37, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Paine, M.J.; Scrutton, N.S.; Munro, A.W.; Gutierrez, A.; Roberts, G.C.; Wolf, C.R. Electron transfer partners of cytochrome P450. In Cytochrome P450-Structure, Mechanism, and Biochemistry; Springer: Boston, MA, USA, 2005; pp. 115–148. [Google Scholar] [CrossRef]
- de Montellano, P.R.O. Cytochrome P450: Structure, Mechanism, and Biochemistry; Springer: New York, NY, USA, 2015. [Google Scholar] [CrossRef]
- Rendic, S.; Guengerich, F.P. Survey of human oxidoreductases and cytochrome P450 enzymes involved in the metabolism of xenobiotic and natural chemicals. Chem. Res. Toxicol. 2015, 28, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Mi, L.; Wang, Z.; Yang, W.; Huang, C.; Zhou, B.; Hu, Y.; Liu, S. Cytochromes P450 in biosensing and biosynthesis applications: Recent progress and future perspectives. Trends Anal. Chem. 2023, 158, 116791. [Google Scholar] [CrossRef]
- van Liempda, S.M.; Kool, J.; Reinen, J.; Schenk, T.; Meermana, J.H.N.; Irth, H.; Vermeulen, N.P.E. Development and validation of a microsomal online cytochrome P450 bioreactor coupled to solid-phase extraction and reversed-phase liquid chromatography. J. Chromatogr. A 2005, 1075, 205–212. [Google Scholar] [CrossRef]
- Guengerich, F.P. Intersection of the Roles of Cytochrome P450 Enzymes with Xenobiotic and Endogenous Substrates: Relevance to Toxicity and Drug Interactions. Chem. Res. Toxicol. 2017, 30, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P. 1.19—Drug Metabolism: Cytochrome P450. In Comprehensive Pharmacology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 470–508. [Google Scholar] [CrossRef]
- Winkler, M.; Geier, M.; Hanlon, S.P.; Nidetzky, B.; Glieder, A. Human Enzymes for Organic Synthesis. Angew. Chem. Int. Ed. 2018, 57, 13406–13423. [Google Scholar] [CrossRef] [Green Version]
- Lamb, D.C.; Waterman, M.R.; Kelly, S.L.; Guengerich, F.P. Cytochromes P450 and drug discovery. Curr. Opin. Biotechnol. 2007, 18, 504–512. [Google Scholar] [CrossRef]
- Brett, C.M.A.; Oliveira Brett, A.M. Electrochemistry: Principles, Methods, and Applications, 1st ed.; Oxford Science University Publications: Oxford, UK, 1993. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Kuzikov, A.V.; Masamrekh, R.A.; Filippova, T.A.; Koroleva, P.I.; Agafonova, L.E.; Bulko, T.V.; Archakov, A.I. Enzymology on an electrode and in a nanopore: Analysis algorithms, enzyme kinetics, and perspectives. BioNanoScience 2022, 12, 1341–1355. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Bulko, T.V.; Alexandrova, S.A.; Sokolov, N.N.; Schmid, R.D.; Bachmann, T.; Archakov, A.I. N-Terminal truncated cytochrome P450 2B4: Catalytic activities and reduction with alternative electron sources. Biochem. Biophys. Res. Commun. 1999, 263, 678–680. [Google Scholar] [CrossRef] [PubMed]
- Shumyantseva, V.V.; Bulko, T.V.; Archakov, A.I. Regulation of cytochrome P450 activity by physicochemical methods. Uspekhi Khimii 1999, 68, 967–975. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Bulko, T.V.; Zimin, A.G.; Uvarov, V.Y.; Archakov, A.I. Design of an artificial hemoprotein based on human serum albumin. Biochem. Mol. Biol. Int. 1996, 39, 503. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Güven, G.; Li, Y.; Schwaneberg, U. First steps towards a Zn/Co(III)sep-driven P450 BM3 reactor. Appl. Microbiol. Biotechnol. 2011, 91, 989–999. [Google Scholar] [CrossRef]
- Zheng, D.; Zhang, Y.; Liu, X.; Wang, J. Coupling natural systems with synthetic chemistry for light driven enzymatic biocatalysis. Photosynth. Res. 2020, 143, 221–231. [Google Scholar] [CrossRef]
- Zilly, F.E.; Taglieber, A.; Schulz, F.; Hollmann, F.; Reetz, M.T. Deazaflavins as mediators in light-driven cytochrome P450 catalyzed hydroxylations. Chem. Commun. 2009, 46, 7152–7154. [Google Scholar] [CrossRef]
- Jensen, K.; Jensen, P.E.; Møller, B.L. Light-driven chemical synthesis. Trends Plant Sci. 2012, 17, 60–63. [Google Scholar] [CrossRef]
- Nielsen, A.Z. Redirecting Photosynthetic Reducing Power toward Bioactive Natural Product Synthesis. ACS Synth. Biol. 2013, 2, 308–315. [Google Scholar] [CrossRef]
- Le, T.-K.; Park, J.H.; Choi, D.S.; Lee, G.-Y.; Choi, W.S.; Jeong, K.J.; Park, C.B.; Yun, C.-H. Solar-driven biocatalytic C-hydroxylation through direct transfer of photoinduced electrons. Green Chem. 2019, 21, 515. [Google Scholar] [CrossRef]
- Lassen, L.M.; Nielsen, A.Z.; Olsen, C.E.; Bialek, W.; Jensen, K.; Møller, B.L.; Jensen, P.E. Anchoring a plant cytochrome P450 via PsaM to the thylakoids in Synechococcus sp. PCC 7002: Evidence for light-driven biosynthesis. PLoS ONE 2014, 9, 102184. [Google Scholar] [CrossRef] [Green Version]
- Shalan, H.; Kato, M.; Cheruzel, L. Keeping the spotlight on cytochrome P450. BBA-Proteins Proteom. 2018, 1866, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Guo, J.; Cheng, F.; Gao, Z.; Du, L.; Meng, C.; Li, S.; Zhang, X. Cytochrome P450s in algae: Bioactive natural product biosynthesis and light-driven bioproduction. Acta Pharm. Sin. B 2022, 12, 2832–2844. [Google Scholar] [CrossRef] [PubMed]
- Eidenschenk, C.; Cheruzel, L. Ru(II)-diimine complexes and cytochrome P450 working hand-in-hand. J. Inorg. Biochem. 2020, 213, 111254. [Google Scholar] [CrossRef] [PubMed]
- Le, T.-K.; Kim, J.; Nguyen, N.A.; Nguyen, T.H.H.; Sun, E.-G.; Yee, S.-M.; Kang, H.-S.; Yeom, S.-J.; Park, C.B.; Yun, C.-H. Solar-Powered Whole-Cell P450 Catalytic Platform for C-Hydroxylation Reactions. ChemSusChem 2021, 14, 3054–3058. [Google Scholar] [CrossRef]
- Lee, J.H.Z.; Podgorski, M.N.; Moir, M.; Gee, A.R.; Bell, S.G. Selective Oxidations Using a Cytochrome P450 Enzyme Variant Driven with Surrogate Oxygen Donors and Light. Chem. Eur. J. 2022, 28, 202201366. [Google Scholar] [CrossRef] [PubMed]
- Shumyantseva, V.V.; Agafonova, L.E.; Bulko, T.V.; Kuzikov, A.V.; Masamrekh, R.A.; Yuan, J.; Pergushov, D.V.; Sigolaeva, L.V. Electroanalysis of Biomolecules: Rational Selection of Sensor Construction. Biochem. (Mosc.) 2021, 86, 140–151. [Google Scholar] [CrossRef]
- Masamrekh, R.A.; Kuzikov, A.V.; Haurychenka, Y.I.; Shcherbakov, K.A.; Veselovsky, A.V.; Filimonov, D.A.; Dmitriev, A.V.; Zavialova, M.G.; Gilep, A.A.; Shkel, T.V.; et al. In vitro interactions of abiraterone, erythromycin, and CYP3A4: Implications for drug-drug interactions. Fundam. Clinic. Pharmacol. 2020, 34, 120–130. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Bulko, T.V.; Kuzikov, A.V.; Masamrekh, R.A.; Konyakhina, A.Y.; Romanenko, I.; Max, J.B.; Köhler, M.; Gilep, A.A.; Usanov, S.A.; et al. All-electrochemical nanocomposite two-electrode setup for quantification of drugs and study of their electrocatalytical conversion by cytochromes P450. Electrochim. Acta 2020, 336, 135579. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Kuzikov, A.V.; Masamrekh, R.A.; Bulko, T.V.; Archakov, A.I. From electrochemistry to enzyme kinetics of cytochrome P450. Biosens. Bioelectron. 2018, 121, 192–204. [Google Scholar] [CrossRef]
- Masamrekh, R.; Filippova, T.; Haurychenka, Y.; Shumyantseva, V.; Kuzikov, A.; Shcherbakov, K.; Veselovsky, A.; Strushkevich, N.; Shkel, T.; Gilep, A.; et al. Estimation of the inhibiting impact of abiraterone D4A metabolite on human steroid 21-monooxygenase (CYP21A2). Steroids 2020, 154, 108528. [Google Scholar] [CrossRef]
- Kostin, V.A.; Zolottsev, V.A.; Kuzikov, A.V.; Masamrekh, R.A.; Shumyantseva, V.V.; Veselovsky, A.V.; Stulov, S.V.; Novikov, R.A.; Timofeev, V.P.; Misharin, A.Y. Oxazolinyl derivatives of [17(20)E]-21-norpregnene differing in the structure of A and B rings. Facile synthesis and inhibition of CYP17A1 catalytic activity. Steroids 2016, 115, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S. Bioelectrodes for evaluating molecular therapeutic and toxicity properties. Curr. Opin. Electrochem. 2019, 19, 20–26. [Google Scholar] [CrossRef]
- Di Nardo, G.; Gilardi, G. Natural Compounds as Pharmaceuticals: The Key Role of Cytochromes P450 Reactivity. Trends Biochem. Sci. 2020, 45, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Sakaki, T. Practical application of cytochrome P450. Biol. Pharm. Bull. 2012, 35, 844–849. [Google Scholar] [CrossRef] [Green Version]
- Kuzikov, A.; Masamrekh, R.; Shkel, T.; Strushkevich, N.; Gilep, A.; Usanov, S.; Archakov, A.; Shumyantseva, V. Assessment of electrocatalytic hydroxylase activity of cytochrome P450 3A4 (CYP3A4) by means of derivatization of 6β-hydroxycortisol by sulfuric acid for fluorimetric assay. Talanta 2019, 196, 231–236. [Google Scholar] [CrossRef]
- Kuzikov, A.; Filippova, T.; Masamrekh, R.; Shumyantseva, V. Biotransformation of phenytoin in electrochemically-driven CYP2C19 system. Biophys. Chem. 2022, 291, 106894. [Google Scholar] [CrossRef]
- Kuzikov, A.; Masamrekh, R.; Filippova, T.; Shumyantseva, V. Electrochemical Analysis of Metabolites as a Method for Cytochromes P450 Activity Determination. Biomed. Chem. Res. Methods 2022, 5, e00176. [Google Scholar] [CrossRef]
- Kuzikov, A.; Filippova, T.; Masamrekh, R.; Shumyantseva, V. Electroanalysis of 4′-Hydroxydiclofenac for CYP2C9 Enzymatic Assay. Electrocatalysis 2022, 13, 630–640. [Google Scholar] [CrossRef]
- Schneider, E.; Clark, D.S. Cytochrome P450 (CYP) enzymes and the development of CYP biosensors. Biosens. Bioelectron. 2013, 39, 1–13. [Google Scholar] [CrossRef]
- Panicco, P.; Castrignanò, S.; Sadeghi, S.J.; Di Nardo, G.; Gilardi, G. Engineered human CYP2C9 and its main polymorphic variants for bioelectrochemical measurements of catalytic response. Bioelectrochemistry 2021, 138, 107729. [Google Scholar] [CrossRef]
- Mi, L.; He, F.; Jiang, L.; Shangguan, L.; Zhang, X.; Ding, T.; Liu, A.; Zhang, Y.; Liu, S. Electrochemically-driven benzo [a] pyrene metabolism via human cytochrome P450 1A1 with reductase coated nitrogen-doped graphene nano-composites. J. Electroanal. Chem. 2017, 804, 23–28. [Google Scholar] [CrossRef]
- Avila-Sierra, A.; Moreno, J.A.; Goode, K.; Zhu, T.; Fryer, P.J.; Taylor, A.; Zhang, Z.J. Effects of structural and chemical properties of surface coatings on the adsorption characteristics of proteins. Surf. Coat. Technol. 2023, 452, 129054. [Google Scholar] [CrossRef]
- Martínez-Negro, M.; Oberländer, J.; Simon, J.; Mailänder, V.; Morsbach, S.; Landfester, K. A new methodology combining QCM-D and proteomic profiling enables characterization of protein adsorption on 2D surfaces. J. Colloid Interface Sci. 2023, 630, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Drug Metabolism: Cytochrome P450. Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Koroleva, P.I.; Bulko, T.V.; Shkel, T.V.; Gilep, A.A.; Veselovsky, A.V. Approaches for increasing the electrocatalitic efficiency of cytochrome P450 3A4. Bioelectrochemistry 2023, 149, 108277. [Google Scholar] [CrossRef]
- Nerimetla, R.; Krishnan, S. Electrocatalysis by subcellular liver fractions bound to carbon nanostructures for stereoselective green drug metabolite synthesis. Chem. Commun. 2015, 51, 11681–11684. [Google Scholar] [CrossRef] [Green Version]
- Walgama, C.; Nerimetla, R.; Materer, N.F.; Schildkraut, D.; Elman, J.F.; Krishnan, S. A Simple Construction of Electrochemical Liver Microsomal Bioreactor for Rapid Drug Metabolism and Inhibition Assays. Anal. Chem. 2015, 87, 4712–4718. [Google Scholar] [CrossRef]
- Sun, X.; Sun, J.; Ye, Y.; Ji, J.; Sheng, L.; Yang, D.; Sun, X. Metabolic pathway-based self-assembled Au@MXene liver microsome electrochemical biosensor for rapid screening of aflatoxin B1. Bioelectrochemistry 2023, 151, 108378. [Google Scholar] [CrossRef]
- Faulkner, K.M.; Shet, M.S.; Fisher, C.W.; Estabrook, R.W. Electrocatalytically driven w-hydroxylation of fatty acids using cytochrome P450 4A1. Proc. Natl. Acad. Sci. USA 1995, 92, 7705–7709. [Google Scholar] [CrossRef] [Green Version]
- Estabrook, R.W.; Faulkner, K.M.; Seth, M.S.; Fisher, C.W. Application of electrochemistry for P450-catalyzed reactions. Methods Enzymol. 1996, 272, 44–51. [Google Scholar] [CrossRef]
- Glass, S.M.; Guengerich, F.P. Cellular retinoid-binding proteins transfer retinoids to human cytochrome P450 27C1 for desaturation. J. Biol. Chem. 2021, 297, 101142. [Google Scholar] [CrossRef]
- Ducharme, J.; Auclair, K. Use of bioconjugation with cytochrome P450 enzymes. BBA-Proteins Proteom. 2018, 1866, 32–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuriato, D.; Correddu, D.; Catucci, G.; Di Nardo, G.; Bolchi, C.; Pallavicini, M.; Gilardi, G. Design of a H2O2-generating P450SPα fusion protein for high yield fatty acid conversion. Protein Sci. 2022, 31, e4501. [Google Scholar] [CrossRef] [PubMed]
- Gilardi, G.; Meharenna, Y.T.; Tsotsou, G.E.; Sadeghi, S.J.; Fairhead, M.; Giannini, S. Molecular Lego: Design of molecular assemblies of P450 enzymes for nanobiotechnology. Biosens. Bioelectron. 2002, 17, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Rua, F.; Sadeghi, S.J.; Castrignanò, S.; Di Nardo, G.; Gilardi, G. Engineering Macaca fascicularis cytochrome P450 2C20 to reduce animal testing for new drugs. J. Inorg. Biochem. 2012, 117, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Dodhia, V.; Sassone, C.; Fantuzzi, A.; Di Nardo, G.; Sadeghi, S.J.; Gilardi, G. Modulating the coupling efficiency of human cytochrome P450 CYP3A4 at electrode surfaces through protein engineering. Electrochem. Commun. 2008, 10, 1744–1747. [Google Scholar] [CrossRef]
- Degregorio, D.; Sadeghi, S.J.; Di Nardo, G.; Gilardi, G.; Solinas, S.P. Understanding uncoupling in the multiredox centre P450 3A4-BMR model system. J. Biol. Inorg. Chem. 2011, 16, 109–116. [Google Scholar] [CrossRef]
- Castrignanò, S.; Di Nardo, G.; Sadeghi, S.J.; Gilardi, G. Influence of inter-domain dynamics and surrounding environment flexibility on the direct electrochemistry and electrocatalysis of selfsufficient cytochrome P450 3A4-BMR chimeras. J. Inorg. Biochem. 2018, 188, 9–17. [Google Scholar] [CrossRef]
- Castrignanò, S.; D’Avino, S.; Di Nardo, G.; Catucci, G.; Sadeghi, S.J.; Gilardi, G. Modulation of the interaction between human P450 3A4 and B. megaterium reductase via engineered loops. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 116–125. [Google Scholar] [CrossRef]
- Degregorio, D.; D’Avino, S.; Castrignanò, S.; Di Nardo, G.; Catucci, G.; Sadeghi, S.J.; Gilardi, G. Human Cytochrome P450 3A4 as a Biocatalyst: Effects of the Engineered Linker in Modulation of Coupling Efficiency in 3A4-BMR Chimeras. Front. Pharmacol. 2017, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Fantuzzi, A.; Meharenna, Y.T.; Briscoe, P.B.; Sassone, C.; Borgia, B.; Gilardi, G. Improving catalytic properties of P450 BM3 haem domain electrodes by molecular Lego. Chem. Commun. 2006, 12, 1289–1291. [Google Scholar] [CrossRef]
- Correddu, D.; Catucci, G.; Giuriato, D.; Di Nardo, G.; Ciaramella, A.; Gilardi, G. Catalytically self-sufficient CYP116B5: Domain switch for improved peroxygenase activity. Biotechnol. J. 2023, 18, 2200622. [Google Scholar] [CrossRef] [PubMed]
- Shumyantseva, V.V.; Bulko, T.V.; Lisitsyna, V.B.; Urlacher, V.B.; Kuzikov, A.V.; Suprun, E.V.; Archakov, A.I. Electrochemical Measurement of Intraprotein and Interprotein Electron Transfer. Biophysics 2013, 58, 349–354. [Google Scholar] [CrossRef]
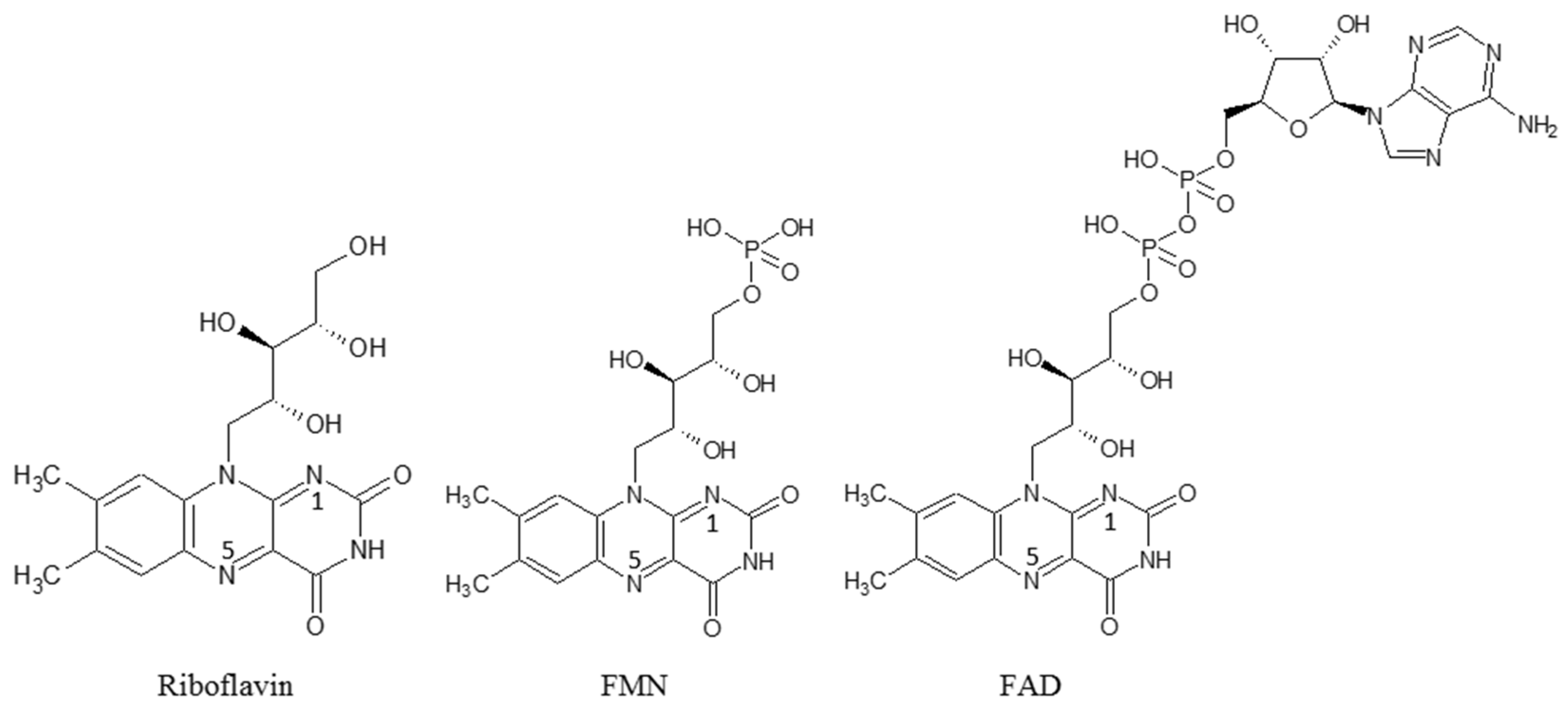
- Zhang, C.; Lu, M.; Lin, L.; Huang, Z.; Zhang, R.; Wu, X.; Chen, Y. Riboflavin Is Directly Involved in N-Dealkylation Catalyzed by Bacterial Cytochrome P450 Monooxygenases. Chembiochem 2020, 21, 2297–2305. [Google Scholar] [CrossRef] [PubMed]
- Shumyantseva, V.; Uvarov, V.; Byakova, O.; Archakov, A. Semisynthetic Flavocytochromes Based on Cytochrome P450 2B4: Reductase and Oxygenase Activities. Arch. Biochem. Biophys. 1998, 354, 133–138. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Bulko, T.V.; Bachmann, T.T.; Bilitewski, U.; Schmid, R.D.; Archakov, A.I. Electrochemical Reduction of Flavocytochromes 2B4 and 1A2 and Their Catalytic Activity. Arch Biochem. Biophys. 2000, 377, 43–48. [Google Scholar] [CrossRef]
- Roux, Y.; Ricoux, R.; Avenier, F.; Mahy, J.-P. Bio-inspired electron-delivering system for reductive activation of dioxygen at metal centres towards artificial flavoenzymes. Nat. Commun. 2015, 6, 8509. [Google Scholar] [CrossRef] [Green Version]
- Sevrioukova, I.F.; Li, H.; Zhang, H.; Peterson, J.A.; Poulos, T.L. Structure of a cytochrome P450–redox partner electron-transfer complex. Proc. Natl. Acad. Sci. USA 1999, 96, 1863–1868. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Shen, Y.; Liu, S. Enhanced light-driven catalytic performance of cytochrome P450 confined in macroporous silica. Chem. Commun. 2016, 52, 7703–7706. [Google Scholar] [CrossRef]
- Fang, X.; Qiao, L.; Yan, G.; Yang, P.; Liu, B. Multifunctional nanoreactor for comprehensive characterization of membrane proteins based on surface functionalized mesoporous foams. Anal. Chem. 2015, 87, 9360–9367. [Google Scholar] [CrossRef]
- Dai, Q.; Yang, L.; Wang, Y.; Cao, X.; Yao, C.; Xu, X. Surface charge-controlled electron transfer and catalytic behavior of immobilized cytochrome P450 BM3 inside dendritic mesoporous silica nanoparticles. Anal. Bioanal. Chem. 2020, 412, 4703–4712. [Google Scholar] [CrossRef]
- Küchler, A.; Yoshimoto, M.; Luginbühl, S.; Mavelli, F.; Walde, P. Enzymatic reactions in confined environments. Nat. Nanotechnol. 2016, 11, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Shumyantseva, V.V.; Koroleva, P.I.; Gilep, A.A.; Napolskii, K.S.; Ivanov, Y.D.; Kanashenko, S.L.; Archakov, A.I. Increasing the Efficiency of Cytochrome P450 3A4 Electrocatalysis Using Electrode Modification with Spatially Ordered Anodic Aluminum Oxide-Based Nanostructures for Investigation of Metabolic Transformations of Drugs. Dokl. Biochem. Biophys. 2022, 506, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, P.I.; Gilep, A.A.; Kraevskiy, S.V.; Tsybruk, T.V.; Shumyantseva, V.V. Improving the Efficiency of Electrocatalysis of Cytochrome P450 3A4 by Modifying the Electrode with Membrane Protein Streptolysin O for Studying the Metabolic Transformations of Drugs. Biosensors 2023, 13, 457. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Chen, Y.; Wang, Y.; Cai, Z.; Xiao, J.; Muhmood, T.; Hu, X. Three-Dimensional Ordered Porous Nanostructures for Lithium−Selenium Battery Cathodes That Confer Superior Energy-Storage Performance. ACS Appl. Mater. Interfaces 2021, 13, 9955–9964. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
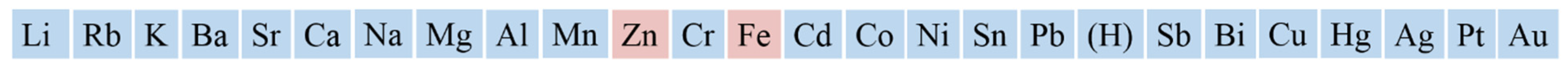{kind=link}
{kind=link}
| Electron Source | Advantages | Disadvantages |
|---|---|---|
| Active metals as electron donor | Label-free approach | Inactivation of enzymes during electro catalysis may occur The lack of commercial standardized sensor devices |
| Light-driven catalysis | Does not need additional electron sources Can be used for broad line of enzymes | Needs additional modification of enzymes with photosensitizers and sacrificial electron donors [25] The lack of commercial devices |
| Electrochemical technology | Additional proteins or redox-active molecules are not necessary Broad spectrum of measurement parameters (catalytic current, Icat; potential start of catalysis, Eonset; potential of catalysis, Ecat; potential of reduction, Ered; potential of oxidation, Eox) [14] Commercial availability of different types of electrodes as 2D sensor | Inactivation of enzymes during electro catalysis may occur |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shumyantseva, V.V.; Koroleva, P.I.; Bulko, T.V.; Agafonova, L.E. Alternative Electron Sources for Cytochrome P450s Catalytic Cycle: Biosensing and Biosynthetic Application. Processes 2023, 11, 1801. https://doi.org/10.3390/pr11061801
Shumyantseva VV, Koroleva PI, Bulko TV, Agafonova LE. Alternative Electron Sources for Cytochrome P450s Catalytic Cycle: Biosensing and Biosynthetic Application. Processes. 2023; 11(6):1801. https://doi.org/10.3390/pr11061801
Chicago/Turabian StyleShumyantseva, Victoria V., Polina I. Koroleva, Tatiana V. Bulko, and Lyubov E. Agafonova. 2023. "Alternative Electron Sources for Cytochrome P450s Catalytic Cycle: Biosensing and Biosynthetic Application" Processes 11, no. 6: 1801. https://doi.org/10.3390/pr11061801