
Novel Stimulants of Medicinal Basidiomycetes Growth Based on Nanoparticles of N-monosubstituted Amino Acid Derivatives of Fullerene C60
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Preparation of Fullerene C60 Water Dispersion
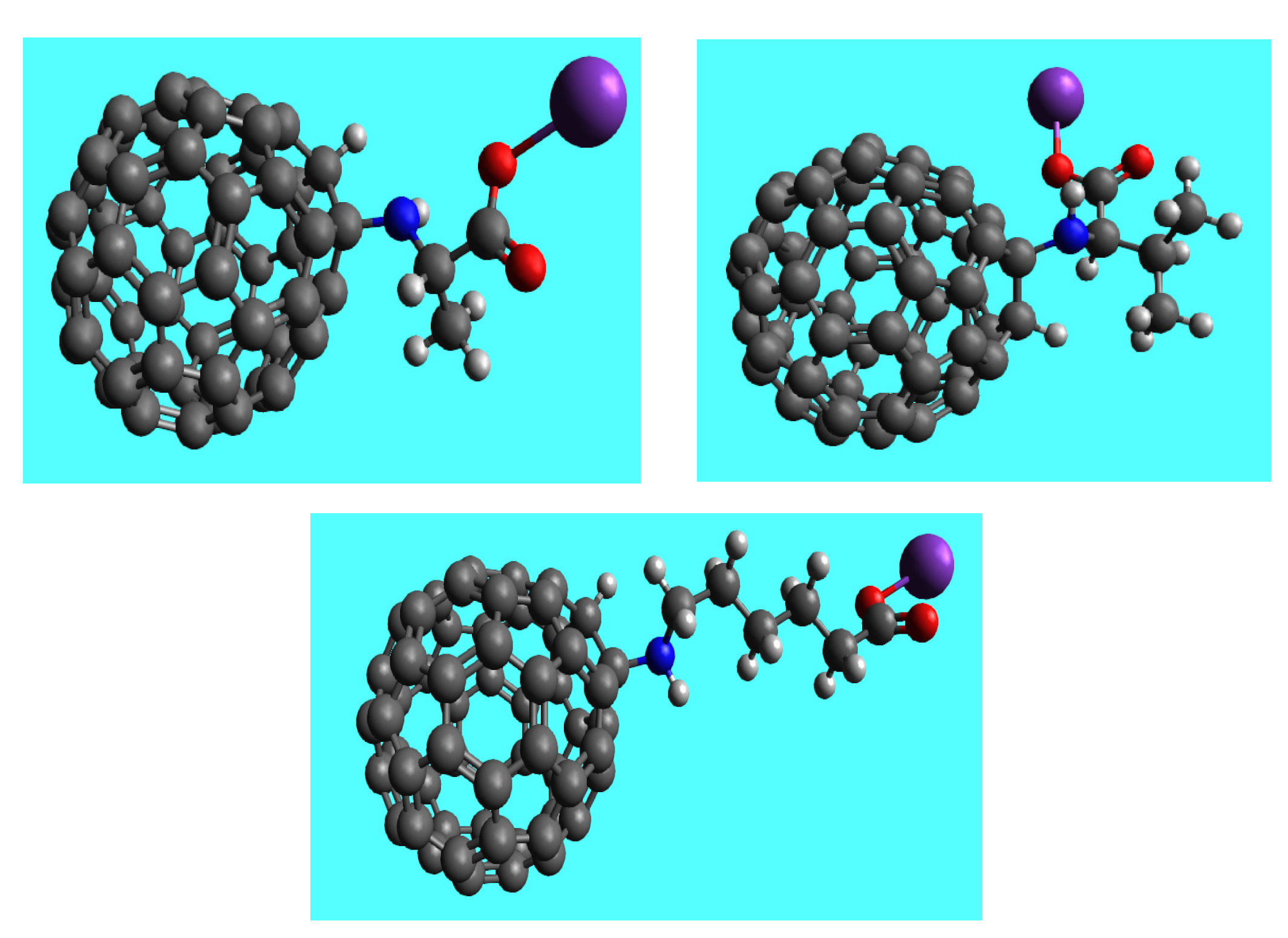
2.1.2. Amino Acid Fullerene Derivatives (AAFD) Synthesis
2.1.3. Strains and Cultivation Conditions
2.2. Methods
2.2.1. Mushroom Mycelium Biomass
2.2.2. Hydrodynamic Radius of Particles
2.2.3. DPPH Scavenging Activity
2.2.4. Lipid Peroxidation
2.2.5. Statistical Processing of Results
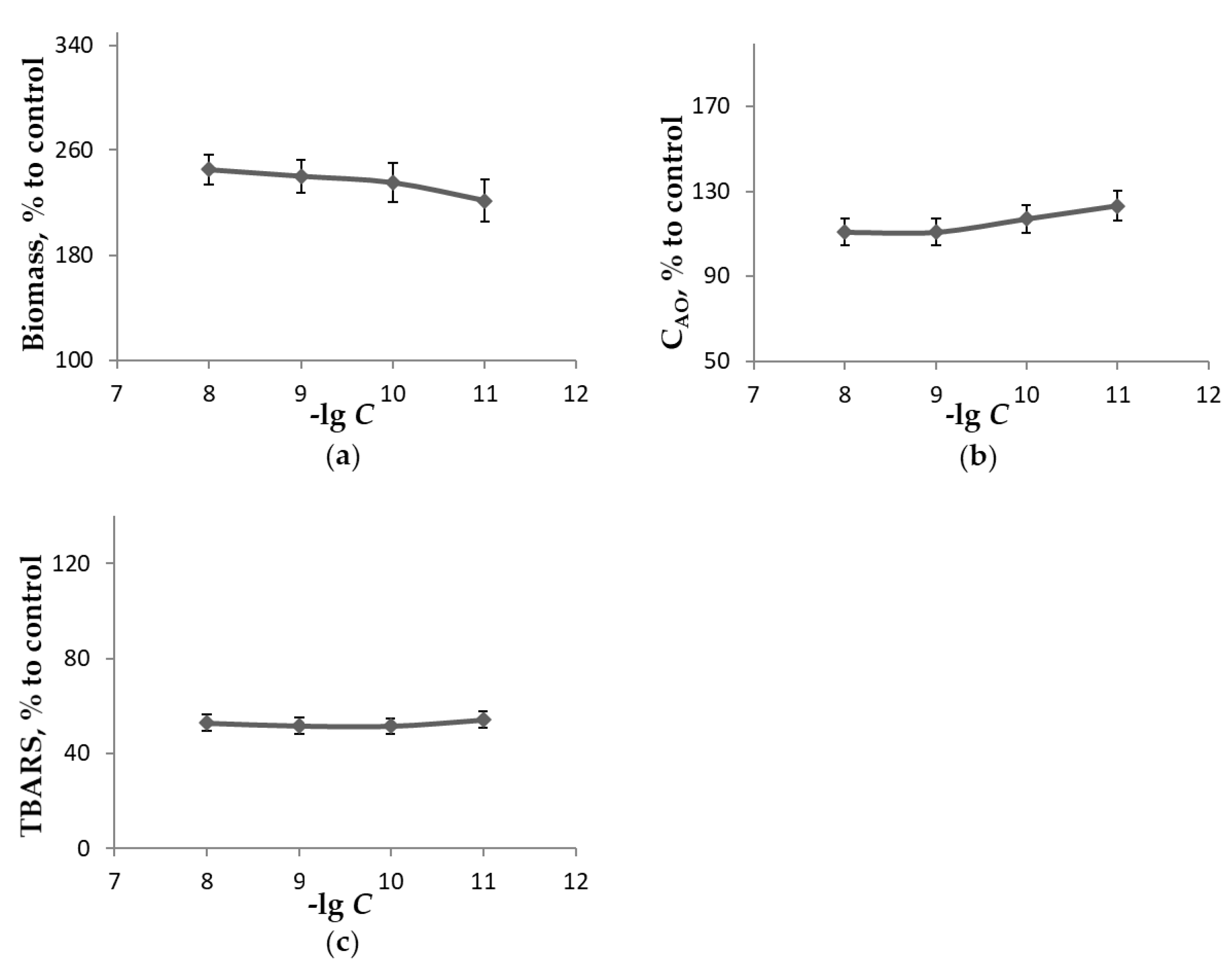
3. Results and Discussion
4. Conclusions
5. Patents
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dai, Y.C.; Yang, Z.L.; Ui, B.K.; Yu, C.J.; Zhou, L.W. Species diversity and utilization of medicinal mushrooms and fungi in China (review). Int. J. Med. Mushr. 2009, 11, 287–302. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, S.; Huang, M.; Xu, A. Antibacterial and antiviral value of the genus Ganoderma P. Karst. species (Aphyllophoromycetideae):A review. Int. J. Med. Mushrooms 2003, 5, 235–246. [Google Scholar] [CrossRef]
- Zhang, M.; Cui, S.W.; Cheung, P.C.K.; Wang, Q. Antitumor polysaccharides from mushrooms: A review on their isolation process, structural characteristics and antitumor activity. Trends Food Sci. Technol. 2007, 18, 4–19. [Google Scholar] [CrossRef]
- Hayakawa, K.; Mitsuhashi, N.; Saito, Y.; Takahashi, M.; Katano, S.; Shiojima, K.; Furuta, M.; Niibe, H. Effect of Krestin (PSK) as adjuvant treatment on the prognosis after radical radiotherapy in patients with non-small cell lung cancer. Anticancer Res. 1993, 13, 1815–1820. [Google Scholar] [PubMed]
- He, Z.; Lin, J.; He, Y.; Liu, S. Polysaccharide-Peptide from Trametes versicolor: The Potential Medicine for Colorectal Cancer Treatment. Biomedicines. 2022, 10, 2841. [Google Scholar] [CrossRef]
- Fernandes, A.; Nair, A.; Kulkarni, N.; Todewale, N.; Jobby, R. Exploring Mushroom Polysaccharides for the Development of Novel Prebiotics: A Review. Int. J. Med. Mushrooms. 2023, 25, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nakao, I.; Uchino, H.; Orita, K.; Kaido, I.; Kimura, T.; Goto, Y.; Kondo, T.; Takino, T.; Taguchi, T.; Nakajima, T.; et al. Clinical evaluation of schizophyllan (SPG) in advanced gastric cancer—A randomized comparative study by an envelope method. GanTo Kagaku Ryoho. 1983, 10, 1146–1159. [Google Scholar]
- Garcia, J.; Rodrigues, F.; Saavedra, M.J.; Nunes, F.M.; Marques, G. Bioactive polysaccharides from medicinal mushrooms: A review on their isolation, structural characteristics and antitumor activity. Food Biosci. 2022, 49, 101955. [Google Scholar] [CrossRef]
- Teplyakova, T.V.; Pyankov, O.V.; Safatov, A.S.; Ovchinnikova, A.S.; Kosogova, T.A.; Skarnovich, M.O.; Filippova, E.I.; Poteshkina, A.L. Water Extract of the Chaga Medicinal Mushroom, Inonotus obliquus (Agaricomycetes), Inhibits SARS-CoV-2 Replication in Vero E6 and Vero Cell Culture Experiments. Int. J. Med. Mushrooms 2022, 24, 23–30. [Google Scholar] [CrossRef]
- Wu, M.; Luo, X.; Xu, X.; Wei, W.; Yu, M.; Jiang, N.; Ye, L.; Yang, Z.; Fei, X. Antioxidant and immunomodulatory activities of a polysaccharide from Flammulina velutipes. J. Tradit. Chines Med. 2014, 34, 733–740. [Google Scholar] [CrossRef]
- Zhang, Z. Inhibitory effect of medicinal fungi on hepatitis B virus in vitro and in vivo. Bull. Beijing Med. Univ. 1989, 21, 455–459. [Google Scholar]
- Liu, J.; Zhorabek, F.; Dai, X.; Huang, J.; Chau, Y. Minimalist design of an intrinsically disordered protein-mimicking scaffold for an artificial membraneless organelle. ACS Cent. Sci. 2022, 8, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Ooi, V.E.; Liu, F. A review of pharmacological activities of mushroom polysaccharides. Int. J. Med. Mushrooms 1999, 1, 196–206. [Google Scholar] [CrossRef]
- Sivanesan, I.; Muthu, M.; Gopal, J.; Oh, J.W. Mushroom polysaccharide-assisted anticarcinogenic mycotherapy: Reviewing its clinical trials. Molecules 2022, 27, 4090. [Google Scholar] [CrossRef]
- Smania, A.; Monache, F.D.; Smania, E.F.; Cuneo, R.S. Antibacterial activity of steroidal compounds isolated from Ganoderma applanatum (Pers.) Pat. (Aphyllophoromycetideae) fruit body. Int. J. Med. Mushr. 1999, 1, 325–330. [Google Scholar] [CrossRef]
- Su, H.G.; Wang, Q.; Zhou, L.; Peng, X.R.; Xiong, W.Y.; Qiu, M.H. Functional triterpenoids from medicinal fungi Ganoderma applanatum: A continuous search for antiadipogenic agents. Bioorganic. Chem. 2021, 112, 104977. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.R.; Wang, Q.; Su, H.G.; Zhou, L.; Xiong, W.Y.; Qiu, M.H. Anti-adipogenic lanostane-type triterpenoids from the edible and medicinal mushroom Ganoderma applanatum. J. Fungi. 2022, 8, 331. [Google Scholar] [CrossRef]
- Li, X.-Y.; Li, S.-Y.; Yin, F.; Chen, H.-M.; Yang, D.-F.; Liu, X.-Q.; Jin, Q.-H.; Lv, X.-M.; Mans, D.; Zhang, X.-D.; et al. Antioxidative and Cytoprotective Effects of Ganoderma applanatum and Fomitopsis pinicola in PC12 Adrenal Phaeochromocytoma Cells. Int. J. Med. Mushrooms 2022, 24, 15–29. [Google Scholar] [CrossRef]
- Hossen, S.M.; Islam, M.J.; Hossain, M.R.; Barua, A.; Uddin, M.G.; Emon, N.U. CNS anti-depressant, anxiolytic and analgesic effects of Ganoderma applanatum (mushroom) along with ligand-receptor binding screening provide new insights: Multi-disciplinary approaches. Biochem. Biophys. Rep. 2021, 27, 101062. [Google Scholar] [CrossRef]
- Wang, J.; Sun, W.; Luo, H.; He, H.; Deng, W.Q.; Zou, K.; Liu, C.; Song, J.; Huang, W. Protective effect of eburicoic acid of the chicken of the woods mushroom, Laetiporus sulphureus (higher Basidiomycetes), against gastric ulcers in mice. Int. J. Med. Mushrooms 2015, 17, 619–626. [Google Scholar] [CrossRef]
- Khatua, S.; Ghosh, S.; Acharya, K. Laetiporus sulphureus (Bull.: Fr.) Murr. as food as medicine. Pharmacogn. J. 2017, 9, s1–s15. [Google Scholar] [CrossRef]
- Patocka, J. Will the sulphur polypore (Laetiporus sulphureus) become a new functional food. Glob. J. Med. Clin. Case Rep. 2019, 6, 006–009. [Google Scholar] [CrossRef]
- Zhang, Y.; Poon, K.; Masonsong, G.S.P.; Ramaswamy, Y.; Singh, G. Sustainable Nanomaterials for Bio-medical Applications. Pharmaceutics 2023, 15, 922. [Google Scholar] [CrossRef]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S.A.; Rehman, H.; Imran Ashraf, I.; Sanaullah, M. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 2022, 721, 137778. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Maragani, S.; Huang, L.; Jeon, S.; Canteenwala, T.; Hamblin, M.R.; Chiang, L.Y. Synthesis of decacationic [60] fullerene decaiodides giving photoinduced production of superoxide radicals and effective PDT-mediation on antimicrobial photoinactivation EUR. J. Med. Chem. 2013, 63, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Vani, J.R.; Mohammadi, M.T.; Foroshani, M.S.; Jafari, M. Polyhydroxylated fullerene nanoparticles attenuate brain infarction and oxidative stress in rat model of ischemic stroke. EXCLI J. 2016, 15, 378–390. [Google Scholar] [CrossRef]
- Castro, E.; Garcia, A.H.; Zavala, G.; Echegoyen, L. Fullerenes in biology and medicine. J. Mater. Chem. B 2017, 5, 6523–6535. [Google Scholar] [CrossRef] [PubMed]
- Panova, G.G.; Kanash, E.V.; Semenov, K.N.; Charykov, N.A.; Khomyakov, Y.V.; Anokona, L.M.; Artem’eva, A.M.; Kornyukhin, D.L.; Vertebnyi, V.E.; Sinyavina, N.G.; et al. Fullerene derivatives influence production process, growth and resistance to oxidative stress in barley and wheat plants. Agric. Biol. 2018, 53, 38–49. [Google Scholar] [CrossRef]
- Yin, J.-J.; Lao, F.; Fu, P.P.; Wamer, W.G.; Zhao, Y.; Wang, P.C.; Qiu, Y.; Sun, B.; Xing, G.; Dong, J.; et al. The scavenging of reactive oxygen species and potential for cell protection by functionalized fullerene materials. Biomaterials 2009, 30, 611–621. [Google Scholar] [CrossRef]
- Mironov, K.S.; Sinetova, M.A.; Shumskaya, M.; Los, D.A. Universal molecular triggers of stress responses in cyanobacterium Synechocystis. Life 2019, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Beuerle, F.; Lebovitz, R.; Hirsch, A. Antioxidant Properties of Water-Soluble Fullerene Derivatives. In Medicinal Chemistry and Pharmacological Potential of Fullerenes and Carbon Nanotubes; Cataldo, F., Da Ros, T., Eds.; Springer Science & Business Media: Dordrecht, The Netherlands, 2008; Volume 1, pp. 51–78. [Google Scholar] [CrossRef]
- Chi, Y.; Bhonsle, J.B.; Canteenwala, T.; Huang, J.-P.; Shiea, J.; Chen, B.-J.; Chiang, L.Y. Novel watersoluble hexakis(4-sulfobutylfullerenes as potent free radical scavengers. Chem. Lett. 1998, 27, 465–466. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.; Lu, Z.; Gao, X. Syntheses, structures and antioxidant activities of fullerenols: Knowledge learned at the atomistic level. J. Clust. Sci. 2015, 26, 375–388. [Google Scholar] [CrossRef]
- Mirkov, S.M.; Djordjevic, A.N.; Andric, N.L.; Andric, S.A.; Kostic, S.A.; Bogdanovic, G.M.; Vojinovic-Miloradov, M.B.; Kvacevic, R.A. Nitric oxide-scavenging activity of polyhydroxylated fullerenol, C60(OH)24. Nitric. Oxide. 2004, 11, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Bensasson, R.V.; Brettreich, M.; Frederiksen, J.; Göttinger, H.; Hirsch, A.; Land, E.J.; Leach, S.; McGarvey, D.J.; Schönberger, H. Reactions of e−aq, CO2•−, HO•, O2•− and O2(1Δg) with a dendro [60]fullerene and C60[C(COOH)2]n (n = 2–6). Free Radic. Biol. Med. 2000, 29, 26–33. [Google Scholar] [CrossRef]
- Andreev, I.M.; Romanova, V.S.; Petrukhina, A.O.; Andreev, S.M. Amino-acid derivatives of fullerene C60 behave as lipophilic ions penetrating through biomembranes. Physic. Solid State 2002, 44, 683–685. [Google Scholar] [CrossRef]
- Schreiner, K.M.; Filley, T.R.; Blanchette, R.A.; Bowen, B.B.; Bolskar, R.D.; Hockaday, W.C.; Masiello, C.A.; Raebigeret, J.W. White-Rot Basidiomycete-Mediated decomposition of C60 Fullerol. Environ. Sci. Technol. 2009, 43, 3162–3168. [Google Scholar] [CrossRef] [PubMed]
- Andrievsky, G.V.; Kosevich, M.V.; Vovk, M.; Shelkovsky, V.S.; Vashchenko, L.A. On the production of an aqueous colloidal solution of fullerenes. J. Chem. Soc. Chem. Commun. 1995, 1281–1282. [Google Scholar] [CrossRef]
- Romanova, V.S.; Tsyryapkin, V.A.; Lyakhovetsky, Y.I.; Parnes, Z.N.; Vol’pin, M.E. Addition of amino acids and dipeptides to fullerene C60 giving rise to monoadducts. Russ. Chem. Bull. 1994, 43, 1090–1091. [Google Scholar] [CrossRef]
- Piotrovsky, L.B.; Eropkin, M.Y.; Eropkina, E.M.; Dumpis, M.A.; Kiselev, O.I. Mechanisms of biological action of fullerenes–dependence on aggregate state. Psychopharmacol. Biol. Narcology 2007, 7, 1548–1554. [Google Scholar]
- Heredia, D.A.; Durantini, A.M.; Durantini, J.E.; Durantini, E.N. Fullerene C60 derivatives as antimicrobial photodynamic agents. J. Photochem. Photobiol. C Photochem. Rev. 2022, 51, 100471. [Google Scholar] [CrossRef]
- Dudka, I.A.; Vasser, S.P.; Ellanskaya, I.A.; Koval, E.E.; Gorbik, L.T.; Bilay, V.I. Methods of Experimental Mycology: Handbook; Bilay, V.I., Ed.; Naukova Dumka: Kiev, Ukraine, 1982. (In Russian) [Google Scholar]
- Bondet, W.; Brand-Williams, W.; Berset, C. Kinetics and Mechanisms of Antioxidant Activity using the DPPH• Free Radical Method. Lebensm. Wiss U. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Volkov, V.A.; Sazhina, N.N.; Pakhomov, P.M.; Misin, V.M. Content and activity of low-molecular antioxidants in food and medicinal plants. Russ. J. Phys. Chem. B 2010, 4, 676–679. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified Thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Volkov, V.A.; Voronkov, M.V.; Misin, V.M.; Yamskova, O.V.; Romanova, V.S.; Kurilov, D.V.; Gagarina, I.N.; Pavlovskaya, N.E.; Gorkova, I.V.; Lushnikov, A.V. New plant growth stimulants based on water-soluble nanoparticles of N-substituted monoamino-acid derivatives of fullerene C60 and the study of their mechanisms of action. Biophysics 2020, 65, 635–641. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Puttaraju, N.G.; Venkateshaiah, S.U.; Dharmesh, S.M.; Urs, S.M.N.; Somasundaram, R. Antioxidant Activity of Indigenous Edible Mushrooms. J. Agric. Food Chem. 2006, 54, 9764–9772. [Google Scholar] [CrossRef]
- Shafiq, F.; Iqbal, M.; Ali, M.; Ashraf, M.A. Seed pre-treatment with polyhydroxy fullerene nanoparticles confer salt tolerance in wheat through up-regulation of H2O2 neutralizing enzymes and phosphorus uptake. J. Soil Sci. Plant Nutr. 2019, 19, 734–742. [Google Scholar] [CrossRef]
- Ferreira, I.C.F.R.; Barros, L.; Abreu, R. Antioxidants in wild mushrooms. Curr. Med. Chem. 2009, 16, 1543–1560. [Google Scholar] [CrossRef]
- Krsmanović, N.; Rašeta, M.; Mišković, J.; Bekvalac, K.; Bogavac, M.; Karaman, M.; Isikhuemhen, O.S. Effects of UV stress in promoting antioxidant activities in fungal species Trametes versicolor (L.) Lloyd and Flammulina velutipes (Curtis) Singer. Antioxidants 2023, 12, 302. [Google Scholar] [CrossRef]
- Hall, C. Sources of natural antioxidants: Oilseeds, nuts, cereals, legumes, animal products and microbial sources. In Antioxidants in Food: Practical Applications; Pokorn, J., Yanishlieva, N., Gordon, M., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2001; pp. 159–209. [Google Scholar]
- Kim, M.-Y.; Seguin, P.; Ahn, J.-K.; Kim, J.-J.; Chun, S.-C.; Kim, E.-H.; Seo, S.-H.; Kang, E.-Y.; Kim, S.-L.; Park, Y.-J.; et al. Phenolic compound concentration and antioxidant activities of edible and medicinal mushrooms from Korea. J. Agric. Food Chem. 2008, 56, 7265–7270. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Tankhaeva, L.M.; Agafonova, S.V. Antioxidant components of Laetiporus sulphureus (Bull.: Fr.) Murr. Fruit bodies. Appl. Biochem. Microbiol. 2011, 47, 419–425. [Google Scholar] [CrossRef]
- Rašeta, M.; Karaman, M.; Jakšić, M.; Šibul, F.; Kebert, M.; Novaković, A.; Popovićet, M. Mineral composition, antioxidant and cytotoxic biopotentials of wild-growing Ganoderma species (Serbia): G. lucidum (Curtis) P. Karst vs. G. applanatum (Pers.) Pat. Int. J. Food Sci. Technol. 2016, 51, 2583–2590. [Google Scholar] [CrossRef]
- Volkov, V.A.; Voronkov, M.V.; Sazhina, N.N.; Kurilov, D.V.; Vokhmyanina, D.V.; Yamskova, O.V.; Martirosyan, Y.T.; Atroshenko, D.L.; Martirosyan, L.Y.; Romanova, V.S. Mechanism of the antioxidant activity and structure–activity relationship of N-monosubstituted amino acid derivatives of fullerene C60. Kinet. Catal. 2021, 62, 395–403. [Google Scholar] [CrossRef]
- Krustic, P.J.; Wasserman, E.; Keizer, P.N.; Morton, J.R.; Preston, K.F. Radical reaction of C60. Science 1991, 254, 1183. [Google Scholar] [CrossRef]
- Foley, S.; Growley, C.; Smaihi, M.; Bonfils, C.; Erlanger, B.F.; Seta, P.; Larroque, C. Cellular localisation of a water-soluble fullerene derivative. Biochem. Biophys. Res. Commun. 2002, 294, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Kotelnikova, R.A.; Kotelnikov, A.I.; Bogdanov, G.N.; Romanova, V.S.; Kuleshova, E.F.; Parnes, Z.N.; Volpin, M.E. Membranotropic properties of the water-soluble amino acid and peptide derivatives of fullerene C60. FEBS Lett. 1996, 389, 111–114. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Name of the Compound | Hydrodynamic Radius r, nm |
|---|---|
| C60 | 55.0 ± 22.7 |
| C60-L-Ala-OK | 4.5 ± 0.8 |
| C60-D-Ala-OK | 2.9 ± 1 |
| C60-D-Val-OK | 15.1 ± 2.7 |
| C60-D-ε-ACA-OK | 6.8 ± 3.5 |
| Sample | Biomass, mg (Control) | Biomass, mg (Experiment) | Biomass, % to Control | CAO, mM/g (Control) | CAO, mM/g (Experiment) | CAO, % to Control | TBARS, microM/g (Control) | TBARS, microM/g (Experiment) | TBARS, % to Control |
|---|---|---|---|---|---|---|---|---|---|
| F. vel. with C60 | 108.2 ± 6.2 | 129.5 ± 4.7 | 119.6 ± 3.6 | 2.0 ± 0.1 | 2.0 ± 0.1 | 97.5 ± 5.6 | 8.5 ± 0.5 | 6.3 ± 0.4 | 74.7 ± 4.8 |
| F. vel. with H-C60-L-Ala-OK | 102.5 ± 1.9 | 116.1 ± 6.5 | 113.2 ± 5.6 | 2.0 ± 0.1 | 2.1 ± 0.1 | 105.0 ± 6.0 | 9.4 ± 0.6 | 9.0 ± 0.6 | 96.0 ± 6.1 |
| F. vel. with H-C60-D-Ala-OK | 125.6 ± 3.6 | 130.9 ± 3.7 | 104.2 ± 2.8 | 1.9 ± 0.1 | 1.6 ± 0.1 | 81.6 ± 4.7 | 6.4 ± 0.4 | 5.9 ± 0.4 | 91.2 ± 5.8 |
| F. vel. with H-C60-D-Val-OK | 102.5 ± 1.9 | 112.1 ± 1.7 | 109.4 ± 1.6 | 2.0 ± 0.1 | 1.9 ± 0.1 | 95.0 ± 6.0 | 9.4 ± 0.6 | 9.3 ± 0.6 | 98.4 ± 6.3 |
| G. app. with H-C60-D-Ala-OK | 38.0 ± 1.8 | 45.0 ± 2.0 | 118.4 ± 4.5 | 5.6 ± 0.3 | 5.7 ± 0.3 | 101.8 ± 5.8 | 15.9 ± 1.0 | 21.5 ± 1.2 | 134.7 ± 7.8 |
| L. sulph. with C60 | 20.1 ± 3.3 | 43.7 ± 3.3 | 217.4 ± 7.6 | 3.4 ± 0.2 | 2.9 ± 0.2 | 85.3 ± 4.9 | 69.3 ± 4.4 | 41.3 ± 2.6 | 59.6 ± 3.8 |
| L. sulph. with H-C60-L-Ala-OK | 18.2 ± 3.8 | 35.8 ± 4.4 | 196.7 ± 12.3 | 5.6 ± 0.3 | 6.6 ± 0.4 | 117.0 ± 6.7 | 57.6 ± 3.7 | 32.0 ± 2.1 | 55.6 ± 3.6 |
| L. sulph. with H-C60-D-Ala-OK | 18.2 ± 3.8 | 43.7 ± 5.5 | 240.1 ± 12.6 | 5.6 ± 0.3 | 6.2 ± 0.4 | 110.7 ± 6.3 | 57.6 ± 3.7 | 29.9 ± 1.9 | 51.9 ± 3.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voronkov, M.; Tsivileva, O.; Volkov, V.; Romanova, V.; Misin, V. Novel Stimulants of Medicinal Basidiomycetes Growth Based on Nanoparticles of N-monosubstituted Amino Acid Derivatives of Fullerene C60. Processes 2023, 11, 1695. https://doi.org/10.3390/pr11061695
Voronkov M, Tsivileva O, Volkov V, Romanova V, Misin V. Novel Stimulants of Medicinal Basidiomycetes Growth Based on Nanoparticles of N-monosubstituted Amino Acid Derivatives of Fullerene C60. Processes. 2023; 11(6):1695. https://doi.org/10.3390/pr11061695
Chicago/Turabian StyleVoronkov, Mikhail, Olga Tsivileva, Vladimir Volkov, Valentina Romanova, and Vyacheslav Misin. 2023. "Novel Stimulants of Medicinal Basidiomycetes Growth Based on Nanoparticles of N-monosubstituted Amino Acid Derivatives of Fullerene C60" Processes 11, no. 6: 1695. https://doi.org/10.3390/pr11061695