
Structure-Dependent Eco-Toxicity of Vegetable Tannin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Determination of Tannin Content
2.3. Acute Toxicity Experiments
2.4. Field Emission Scanning Electron Microscope (FE-SEM)
2.5. Molecular Simulation
2.5.1. Molecular Docking
2.5.2. Molecular Dynamics Simulation
3. Result and Discussion
3.1. Component Analysis
3.2. Toxicity of Vegetable Tannin and the Precursor Chemicals
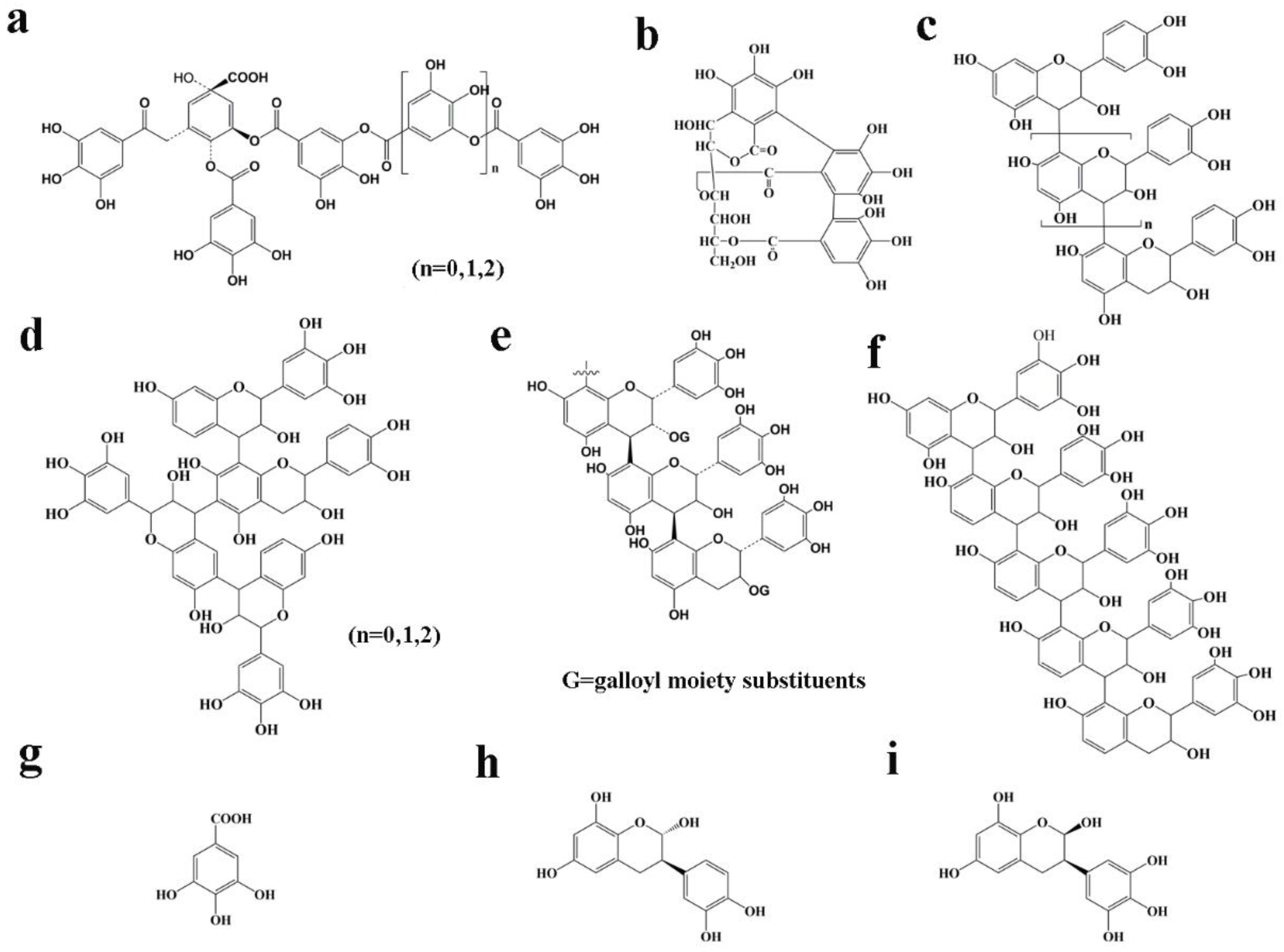
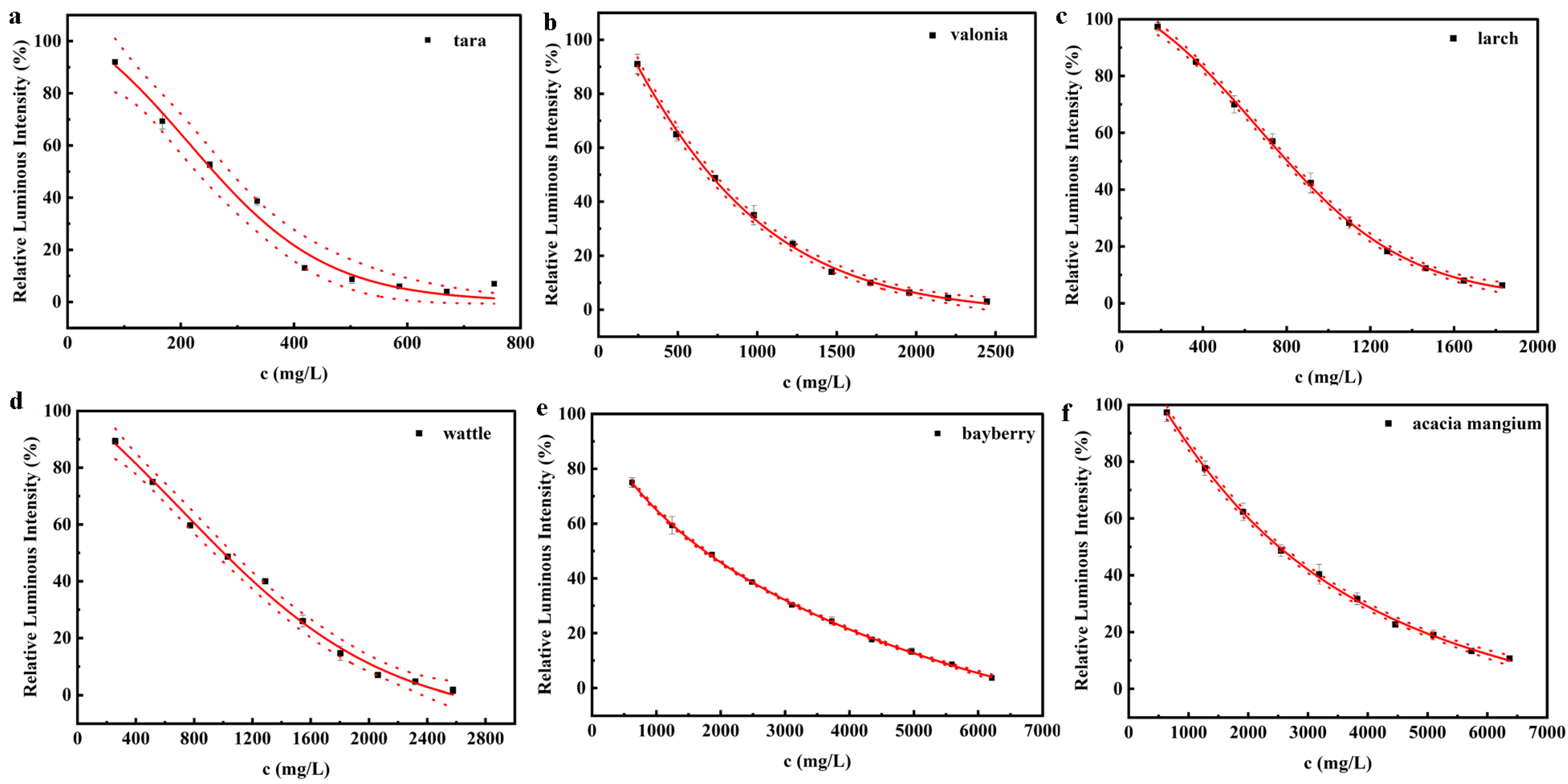
3.2.1. Toxicity of Hydrolyzed Tannin
3.2.2. Toxicity of Condensed Tannin
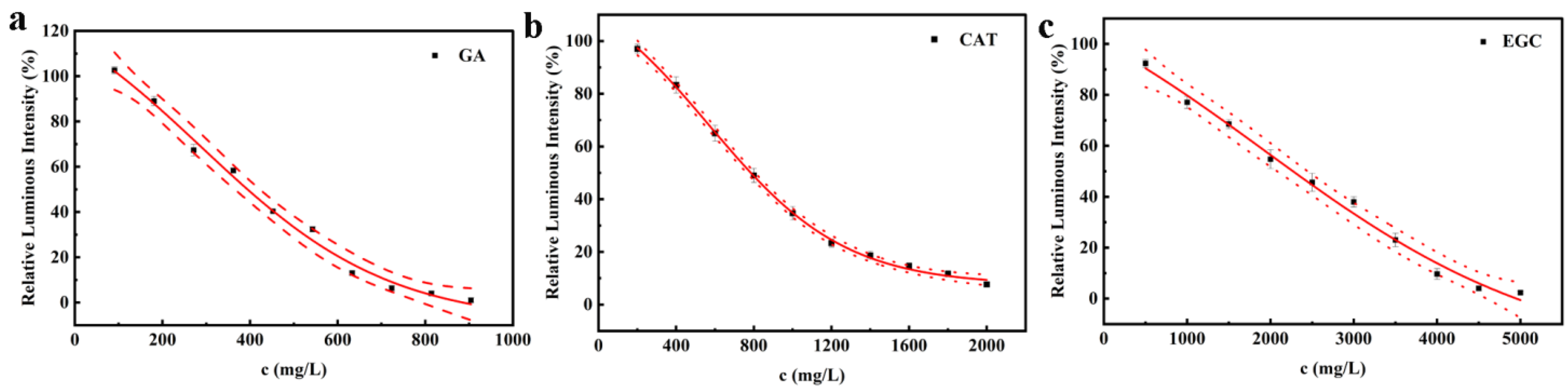
3.2.3. Toxicity of Model Compounds
3.3. Morphologies of P. phosphoreum
3.4. Toxicity Mechanism
3.4.1. Molecular Docking
3.4.2. Molecular Dynamics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cassano, A.; Adzet, J.; Molinari, R.; Buonomenna, M.G.; Roig, J.; Drioli, E. Membrane treatment by nanofiltration of exhausted vegetable tannin liquors from the leather industry. Water Res. 2003, 37, 2426–2434. [Google Scholar] [CrossRef]
- Aguilar, C.N.; Rodriguez, R.; Gutierrez-Sanchez, G.; Augur, C.; Favela-Torres, E.; Prado-Barragan, L.A.; Ramirez-Coronel, A.; Contreras-Esquivel, J.C. Microbial tannases: Advances and perspectives. Appl. Microbiol. Biotechnol. 2007, 76, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Romero-Dondiz, E.M.; Almazán, J.E.; Rajal, V.B.; Castro-Vidaurre, E.F. Removal of vegetable tannins to recover water in the leather industry by ultrafiltration polymeric membranes. Chem. Eng. Res. Des. 2015, 93, 727–735. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Peter Constabel, C. Tannins in plant-herbivore interactions. Phytochemistry 2011, 72, 1551–1565. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Puupponen-Pimia, R.; Dauer, A.; Aura, A.M.; Saura-Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Res. 2009, 53 (Suppl. S2), S310–S329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanbabaee, K.; Ree, V. Tannins: Classification and definition. Nat. Prod. Rep. 2001, 18, 641–649. [Google Scholar] [CrossRef] [PubMed]
- China, C.R.; Nyandoro, S.S.; Munissi, J.J.E.; Maguta, M.M.; Meyer, M.; Schroepfer, M. Tanning capacity of tessmannia burttii extracts: The potential eco-friendly tanning agents for the leather industry. J. Leather Sci. Eng. 2021, 3, 149–157. [Google Scholar] [CrossRef]
- Scholz, W.; Lucas, M. Techno-economic evaluation of membrane filtration for the recovery and re-use of tanning chemicals. Water Res. 2003, 37, 1859–1867. [Google Scholar] [CrossRef]
- Falcao, L.; Araujo, M.E.M. Vegetable tannins used in the manufacture of historic leathers. Molecules 2018, 23, 1081. [Google Scholar] [CrossRef] [Green Version]
- Kanth, S.V.; Venba, R.; Madhan, B.; Chandrababu, N.K.; Sadulla, S. Cleaner tanning practices for tannery pollution abatement: Role of enzymes in eco-friendly vegetable tanning. J. Clean. Prod. 2009, 17, 507–515. [Google Scholar] [CrossRef]
- Ibrahim, A.; Yaser, A.Z.; Lamaming, J. Synthesising tannin-based coagulants for water and wastewater application: A review. J. Environ. Chem. Eng. 2021, 9, 105007. [Google Scholar] [CrossRef]
- Balakrishnan, A.; Kanchinadham, S.; Kalyanaraman, C. Respirometric assessment of vegetable tanning process wastewater generated from tanneries. Eng. Rep. 2020, 2, e12229. [Google Scholar] [CrossRef]
- Jochimsen, J.C.; Jekel, M.R. Partial oxidation effects during the combined oxidative and biological treatment of separated streams of tannery wastewater. Water Sci. Technol. 1997, 35, 337–345. [Google Scholar] [CrossRef]
- Barbero-Lopez, A.; Akkanen, J.; Lappalainen, R.; Peraniemi, S.; Haapala, A. Bio-based wood preservatives: Their efficiency, leaching and ecotoxicity compared to a commercial wood preservative. Sci. Total Environ. 2021, 753, 142013. [Google Scholar] [CrossRef] [PubMed]
- Libralato, G.; Avezzu, F.; Volpi Ghirardini, A. Lignin and tannin toxicity to Phaeodactylum tricornutum (Bohlin). J. Hazard. Mater. 2011, 194, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Germirli Babuna, F.; Yilmaz, Z.; Okay, O.S.; Arslan Alaton, I.; Iskender, G. Ozonation of synthetic versus natural textile tannins: Recalcitrance and toxicity towards Phaeodactylum tricornutum. Water Sci. Technol. 2007, 55, 45–52. [Google Scholar] [CrossRef]
- De Nicola, E.; Gallo, M.; Iaccarino, M.; Meric, S.; Oral, R.; Russo, T.; Sorrentino, T.; Tunay, O.; Vuttariello, E.; Warnau, M.; et al. Hormetic versus toxic effects of vegetable tannin in a multitest study. Arch. Environ. Contam. Toxicol. 2004, 46, 336–344. [Google Scholar] [CrossRef]
- De Nicola, E.; Meric, S.; Gallo, M.; Iaccarino, M.; Della Rocca, C.; Lofrano, G.; Russo, T.; Pagano, G. Vegetable and synthetic tannins induce hormesis/toxicity in sea urchin early development and in algal growth. Environ. Pollut. 2007, 146, 46–54. [Google Scholar] [CrossRef]
- Mariscal, A.; Peinado, M.T.; Carnero-Varo, M.; Fernandez-Crehuet, J. Influence of organic solvents on the sensitivity of a bioluminescence toxicity test with Vibrio harveyi. Chemosphere 2003, 50, 349–354. [Google Scholar] [CrossRef]
- Parvez, S.; Venkataraman, C.; Mukherji, S. A review on advantages of implementing luminescence inhibition test (Vibrio fischeri) for acute toxicity prediction of chemicals. Environ. Int. 2006, 32, 265–268. [Google Scholar] [CrossRef]
- Mohseni, M.; Abbaszadeh, J.; Maghool, S.S.; Chaichi, M.J. Heavy metals detection using biosensor cells of a novel marine luminescent bacterium Vibrio sp. mm1 isolated from the caspian sea. Ecotoxicol. Environ. Saf. 2018, 148, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.J.; Long, W.J.; Xu, T.; Peng, L.Q.; Zhang, W.H. Organic ligands unexpectedly increase the toxicity of chromium(III) for luminescent bacteria. Environ. Chem. Lett. 2019, 17, 1849–1855. [Google Scholar] [CrossRef]
- Li, J.; Yuan, T.; Ma, Y.; Shen, Z.; Tian, Y.; Gao, L.; Dong, X. Acute toxicity assessment of indoor dust extracts by luminescent bacteria assays with photobacterium phosphoreum T3. Environ. Res. 2020, 198, 110447. [Google Scholar] [CrossRef]
- Liu, L.; Xiao, X.; Li, K.; Li, X.; Yu, K.; Liao, X.; Shi, B. Prevention of bacterial colonization based on self-assembled metal-Phenolic nanocoating from rare-earth ions and catechin. ACS Appl. Mater. Inter. 2020, 12, 22237–22245. [Google Scholar] [CrossRef]
- Ishibashi, H.; Uchida, M.; Hirano, M.; Hayashi, T.; Yamamoto, R.; Kubota, A.; Ichikawa, N.; Ishibashi, Y.; Tominaga, N.; Arizono, K. In vivo and in silico analyses of estrogenic potential of equine estrogens in medaka (Oryzias latipes). Sci. Total Environ. 2021, 767, 144379. [Google Scholar] [CrossRef] [PubMed]
- Sigurnjak Bures, M.; Ukic, S.; Cvetnic, M.; Prevaric, V.; Markic, M.; Rogosic, M.; Kusic, H.; Bolanca, T. Toxicity of binary mixtures of pesticides and pharmaceuticals toward Vibrio fischeri: Assessment by quantitative structure-activity relationships. Environ. Pollut. 2021, 275, 115885. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Tang, J.; Chen, J.; Zhang, Q. Quantitative structure-toxicity relationship analysis of combined toxic effects of lignocellulose-derived inhibitors on bioethanol production. Bioresour. Technol. 2019, 289, 121724. [Google Scholar] [CrossRef]
- Han, W.M.; Hou, M.C.; He, F.M.; Zhang, W.H.; Shi, B. Ecotoxicity and interacting mechanism of anionic surfactant sodium dodecyl sulfate (SDS) and its mixtures with nonionic surfactant fatty alcohol-polyoxyethlene ether (AEO). Aqua. Toxicol. 2020, 222, 9. [Google Scholar] [CrossRef]
- Shen, L.; Johnson, T.L.; Clugston, S.; Huang, H.; Butenhof, K.J.; Stanton, R.V. Molecular dynamics simulation and binding energy calculation for estimation of oligonucleotide duplex thermostability in rna-based therapeutics. J. Chem. Inf. Model. 2011, 51, 1957–1965. [Google Scholar] [CrossRef]
- Wang, L.; Ma, M.; Yu, Z.; Du, S.K. Preparation and identification of antioxidant peptides from cottonseed proteins. Food Chem. 2021, 352, 129399. [Google Scholar] [CrossRef]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved side-chain torsion potentials for the Amber ff99SB protein force field. Proteins 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, C.; Kasavajhala, K.; Belfon, K.A.A.; Raguette, L.; Huang, H.; Migues, A.N.; Bickel, J.; Wang, Y.Z.; Pincay, J.; Wu, Q.; et al. ff19SB: Amino-acid specific protein backbone parameters trained against quantum mechanics energy surfaces in solution. J. Chem. Theory Comput. 2020, 16, 528–552. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Tang, Q.; He, J.; Du, S.; Xu, S.; Wang, C.; Xu, Y.; Lin, F. VFFDT: A new software for preparing amber force field parameters for metal-containing molecular systems. J. Chem. Inf. Model. 2016, 56, 811–818. [Google Scholar] [CrossRef]
- Hofer, F.; Kraml, J.; Kahler, U.; Kamenik, A.S.; Liedl, K.R. Catalytic Site pKa Values of aspartic, cysteine, and serine proteases: Constant pH MD Simulations. J. Chem. Inf. Model 2020, 60, 3030–3042. [Google Scholar] [CrossRef]
- Figueira, T.R.E.S.; Okura, V.; Rodrigues da Silva, F.; Jose da Silva, M.; Kudrna, D.; Ammiraju, J.S.S.; Talag, J.; Wing, R.; Arruda, P. A bac library of the sp80-3280 sugarcane variety (Saccharum sp.) and its inferred microsynteny with the sorghum genome. BMC Res. Notes 2012, 5, 185. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.W.S.D.; Vranken, W.F.V. ACPYPE-Antechamber python parser interface. BMC Res. Notes 2012, 5, 367. [Google Scholar] [CrossRef] [Green Version]
- Boyd, N.J.; Wilson, M.R. Optimization of the GAFF force field to describe liquid crystal molecules: The path to a dramatic improvement in transition temperature predictions. Phys. Chem. Chem. Phys. 2015, 17, 24851–24865. [Google Scholar] [CrossRef] [Green Version]
- Enoch, S.J.; Cronin, M.T.; Schultz, T.W.; Madden, J.C. An evaluation of global QSAR models for the prediction of the toxicity of phenols to tetrahymena pyriformis. Chemosphere 2008, 71, 1225–1232. [Google Scholar] [CrossRef]
- Verhaar, H.; Leeuwen, C.; Hermens, J. Classifying environmental pollutants. Chemosphere 1992, 25, 471–491. [Google Scholar] [CrossRef]
- Wang, X.H.; Fan, L.Y.; Wang, S.; Wang, Y.; Yan, L.C.; Zheng, S.S.; Martyniuk, C.J.; Zhao, Y.H. Relationship between acute and chronic toxicity for prevalent organic pollutants in Vibrio fischeri based upon chemical mode of action. J. Hazard. Mater. 2017, 338, 458–465. [Google Scholar] [CrossRef]
- Uribe, S.T.; López-Giraldo, L.J.; Decker, E.A. Relationship between the physiochemical properties of cocoa procyanidins and their ability to inhibit lipid oxidation in liposomes. J. Agric. Food Chem. 2018, 66, 4490–4502. [Google Scholar] [CrossRef] [PubMed]
- Casoni, D.; Sarbu, C. Modeling of food preservatives chromatographic lipophilicity applying genetic algorithm and multiple linear regression. Rev. Roum. Chim. 2011, 56, 381–389. [Google Scholar] [CrossRef]
- Bacilieri, M.; Paoletta, S.; Basili, S.; Fanton, M.; Moro, S. A novel generalized 3d-qsar model of camptothecin analogs. Qsar Comb. Sci. 2011, 30, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Tinikul, R.; Chunthaboon, P.; Phonbuppha, J.; Paladkong, T. Bacterial luciferase: Molecular mechanisms and applications. Enzymes 2020, 47, 427–455. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.Y.; Huang, Y.; Huang, T.; Zhao, K.; Zhang, Y.N.; Li, C.; Zhao, Y.H. Photolysis and photo-induced toxicity of pyraclostrobin to Vibrio fischeri: Pathway and toxic mechanism. Aqua. Toxicol. 2020, 220, 105417. [Google Scholar] [CrossRef] [PubMed]
- Bayles, A.V.; Fisher, J.M.; Valentine, C.S.; Nowbahar, A.; Helgeson, M.E.; Squires, T.M. Hydrogen bonding strength determines water diffusivity in polymer ionogels. J. Phys. Chem. B 2021, 125, 5408–5419. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Tara Tannin | Valonia Tannin | Larch Tannin | Wattle Tannin | Bayberry Tannin | Acacia Mangium Tannin |
|---|---|---|---|---|---|---|
| Moisture/% | 11.0 | 14.1 | 10.6 | 11.5 | 12.7 | 13.1 |
| Tannin/% | 83.7 | 62.6 | 61.0 | 64.4 | 62.1 | 63.7 |
| Non-tannin/% | 0.5 | 18.9 | 25.6 | 22.0 | 20.0 | 19.9 |
| Insoluble matter/% | 5.0 | 4.5 | 2.8 | 2.1 | 5.2 | 3.3 |
| Compound | CAS | Abb. | MW | Log Kow | Log 1/EC50 (Pred) | Log 1/EC50 (Exp) |
|---|---|---|---|---|---|---|
| Gallic acid | 149-91-7 | GA | 170.12 | 0.70 | 1.45 | 2.63 |
| Catechin | 7295-85-4 | CAT | 290.27 | 0.37 | 1.18 | 2.57 |
| Epigallocatechin | 970-74-1 | EGC | 306.27 | 1.12 | 1.88 | 2.13 |
| Compound | Site | CDOCKER Energy (kcal/mol) | H-Bonds | Pi-Bonds |
|---|---|---|---|---|
| GA | 2 | −39.35 | THR80 (LUX A) PHE117 (LUX A) HIS82 (LUX B) ARG85 (LUX B) ARG119 (LUX B) | ALA81 (LUX A) |
| CAT | 2 | −36.63 | PRO79 (LUX A) ARG85 (LUX A) THR119 (LUX A) LYS274 (LUX A) HIS81 (LUX B) | ALA81 (LUX A) ARG85 (LUX B) |
| EGC | 1 | −35.88 | TYR110 (LUX A) ALA174 (LUX A) GLU175 (LUX A) ILE191 (LUX A) SER227 (LUX A) TYR254 (LUX A) | LEU109 (LUX A) VAL173 (LUX A) LEU192 (LUX A) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, F.; Peng, L.; Long, W.; Jiang, X.; Zhang, W. Structure-Dependent Eco-Toxicity of Vegetable Tannin. Processes 2022, 10, 816. https://doi.org/10.3390/pr10050816
He F, Peng L, Long W, Jiang X, Zhang W. Structure-Dependent Eco-Toxicity of Vegetable Tannin. Processes. 2022; 10(5):816. https://doi.org/10.3390/pr10050816
Chicago/Turabian StyleHe, Faming, Liangqiong Peng, Wenjun Long, Xiaofeng Jiang, and Wenhua Zhang. 2022. "Structure-Dependent Eco-Toxicity of Vegetable Tannin" Processes 10, no. 5: 816. https://doi.org/10.3390/pr10050816