
(S)-5-Methylmellein Isolated from an Endogenous Lichen Fungus Rosellinia corticium as a Potent Inhibitor of Human Monoamine Oxidase A

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining and Preparation of ELF Extracts to Assess MAO-A Inhibitory Activity
2.2. Enzyme Activity Assays
2.3. Culture and Extraction of ELF68
2.4. Isolation and Identification of Compounds
2.5. Kinetic Study of the Compounds
2.6. Analysis of Inhibitor Reversibility
2.7. Molecular Docking Simulation of the Compound with hMAO-A and hMAO-B
2.8. Analysis of Pharmacokinetic and Physicochemical Parameters In Silico
3. Results
3.1. Inhibitory Activity of ELF Extracts against hMAO-A
3.2. Compound Isolation of ELF68 Using Prep TLC
3.3. Identification of ELF68
3.4. Inhibitory Activities of 5MM against MAOs, ChEs, and BACE1
3.5. Kinetic Studies of 5MM
3.6. Analysis of Reversibility for hMAO-A by 5MM
3.7. Molecular Docking Simulation
3.8. In Silico Pharmacokinetics of 5MM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramsay, R.R. Monoamine Oxidases: The Biochemistry of the Proteins as Targets in Medicinal Chemistry and Drug Discovery. Curr. Top. Med. Chem. 2012, 12, 2189–2209. [Google Scholar] [CrossRef] [PubMed]
- Youdim, M.B.H.; Edmondson, D.; Tipton, K.F. The Therapeutic Potential of Monoamine Oxidase Inhibitors. Nat. Rev. Neurosci. 2006, 7, 295–309. [Google Scholar] [CrossRef]
- Joao Matos, M.; Vina, D.; Vazquez-Rodriguez, S.; Uriarte, E.; Santana, L. Focusing on New Monoamine Oxidase Inhibitors: Differently Substituted Coumarins As An Interesting Scaffold. Curr. Top. Med. Chem. 2012, 12, 2210–2239. [Google Scholar] [CrossRef]
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase Inhibitors: Pharmacology and Toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [Green Version]
- Bierer, L.M.; Haroutunian, V.; Gabriel, S.; Knott, P.J.; Carlin, L.S.; Purohit, D.P.; Perl, D.P.; Schmeidler, J.; Kanof, P.; Davis, K.L. Neurochemical Correlates of Dementia Severity in Alzheimer’s Disease: Relative Importance of the Cholinergic Deficits. J. Neurochem. 1995, 64, 749–760. [Google Scholar] [CrossRef]
- Kumar, A.; Pintus, F.; Di Petrillo, A.; Medda, R.; Caria, P.; Matos, M.J.; Viña, D.; Pieroni, E.; Delogu, F.; Era, B.; et al. Novel 2-Pheynlbenzofuran Derivatives as Selective Butyrylcholinesterase Inhibitors for Alzheimer’s Disease. Sci. Rep. 2018, 8, 4424. [Google Scholar] [CrossRef] [Green Version]
- Sicinska, P.; Bukowska, B.; Pajak, A.; Koceva-Chyla, A.; Pietras, T.; Nizinkowski, P.; Gorski, P.; Koter-Michalak, M. Decreased Activity of Butyrylcholinesterase in Blood Plasma of Patients with Chronic Obstructive Pulmonary Disease. Arch. Med. Sci. 2017, 3, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, P.; Gilbert, P. Psychotherapy and Counselling for Depression; SAGE: Los Angeles, CA, USA, 2007; ISBN 978-1-84920-349-4. [Google Scholar]
- De Zwart, P.L.; Jeronimus, B.F.; de Jonge, P. Empirical Evidence for Definitions of Episode, Remission, Recovery, Relapse and Recurrence in Depression: A Systematic Review. Epidemiol. Psychiatr. Sci. 2019, 28, 544–562. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.; Blier, P. Monoamine Neurocircuitry in Depression and Strategies for New Treatments. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef] [PubMed]
- López-Muñoz, F.; Alamo, C. Monoaminergic Neurotransmission: The History of the Discovery of Antidepressants from 1950s until Today. Curr. Pharm. Des. 2009, 15, 1563–1586. [Google Scholar] [CrossRef] [Green Version]
- Suerinck, A.; Suerinck, E. Depressive states in a sanatorium milieu and monoamine oxidase inhibitors. (Therapeutic results by the combination of iproclozide and chlordiazepoxide). Apropos of 146 cases. J. Med. Lyon 1966, 47, 573–586. [Google Scholar] [PubMed]
- Fagervall, I.; Ross, S.B. Inhibition of Monoamine Oxidase in Monoaminergic Neurones in the Rat Brain by Irreversible Inhibitors. Biochem. Pharmacol. 1986, 35, 1381–1387. [Google Scholar] [CrossRef]
- Burns, A.; Iliffe, S. Alzheimer’s Disease. BMJ 2009, 338, b158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.M.; Holtzman, D.M. Alzheimer Disease: An Update on Pathobiology and Treatment Strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.-M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The Cholinergic System in the Pathophysiology and Treatment of Alzheimer’s Disease. Brain J. Neurol. 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Paroni, G.; Bisceglia, P.; Seripa, D. Understanding the Amyloid Hypothesis in Alzheimer’s Disease. J. Alzheimers Dis. 2019, 68, 493–510. [Google Scholar] [CrossRef]
- Riederer, P.; Danielczyk, W.; Grünblatt, E. Monoamine Oxidase-B Inhibition in Alzheimer’s Disease. Neurotoxicology 2004, 25, 271–277. [Google Scholar] [CrossRef]
- Zheng, Y.; Xiao, C.-J.; Guo, K.; Wang, Y.; Liu, Y.; Luo, S.-H.; Li, X.-N.; Li, S.-H. Lobarioid A, Unusual Antibacterial Depsidone Possessing an Eight-Membered Diether Ring from the Edible Lichen Lobaria sp. Tetrahedron Lett. 2018, 59, 743–746. [Google Scholar] [CrossRef]
- Dobson, F.S. Lichens: An Illustrated Guide to the British and Irish Species, Sixth Revised and Enlarged Edition; The Richmond Publishing Co., Ltd.: Slough, UK, 2011; ISBN 978-0-85546-315-1. [Google Scholar]
- Dean, J. Wild Color; Watson-Guptill Publications: New York, NY, USA, 1999; ISBN 978-0-8230-5727-6. [Google Scholar]
- Solárová, Z.; Liskova, A.; Samec, M.; Kubatka, P.; Büsselberg, D.; Solár, P. Anticancer Potential of Lichens’ Secondary Metabolites. Biomolecules 2020, 10, 87. [Google Scholar] [CrossRef] [Green Version]
- Samanthi, K.; Wickramaarachchi, S.; Wijeratne, E.; Paranagama, P. Two New Antioxidant Active Polyketides from Penicillium Citrinum, an Endolichenic Fungus Isolated from Parmotreama Species in Sri Lanka. J. Natl. Sci. Found. Sri Lanka 2015, 43, 119. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Kim, Y.; Gang, H.-S.; Han, J.; Ha, H.-H.; Kim, H. Antimicrobial Activity of Divaricatic Acid Isolated from the Lichen Evernia Mesomorpha against Methicillin-Resistant Staphylococcus Aureus. Molecules 2018, 23, 3068. [Google Scholar] [CrossRef] [Green Version]
- Jeong, G.-S.; Kang, M.-G.; Han, S.-A.; Noh, J.-I.; Park, J.-E.; Nam, S.-J.; Park, D.; Yee, S.-T.; Kim, H. Selective Inhibition of Human Monoamine Oxidase B by 5-Hydroxy-2-Methyl-Chroman-4-One Isolated from an Endogenous Lichen Fungus Daldinia Fissa. J. Fungi 2021, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Xiong, J.; Guan, H.-D.; Wang, C.-H.; Lei, X.; Hu, J.-F. Discovery, Synthesis, Biological Evaluation and Molecular Docking Study of (R)-5-Methylmellein and Its Analogs as Selective Monoamine Oxidase A Inhibitors. Bioorg. Med. Chem. 2019, 27, 2027–2040. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.S.; Hillman, P.F.; Kang, M.-G.; Hwang, S.; Park, J.-E.; Nam, S.-J.; Park, D.; Kim, H. Potent and Selective Inhibitors of Human Monoamine Oxidase A from an Endogenous Lichen Fungus Diaporthe Mahothocarpus. J. Fungi 2021, 7, 876. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.C.; Choi, B.; Nam, S.-J.; Kim, H. Inhibition of Monoamine Oxidase A and B by Demethoxycurcumin and Bisdemethoxycurcumin. J. Appl. Biol. Chem. 2018, 61, 187–190. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Lee, J.P.; Kang, M.-G.; Lee, J.Y.; Oh, J.M.; Baek, S.C.; Leem, H.H.; Park, D.; Cho, M.-L.; Kim, H. Potent Inhibition of Acetylcholinesterase by Sargachromanol I from Sargassum Siliquastrum and by Selected Natural Compounds. Bioorg. Chem. 2019, 89, 103043. [Google Scholar] [CrossRef]
- Wilson, E.N.; Do Carmo, S.; Iulita, M.F.; Hall, H.; Ducatenzeiler, A.; Marks, A.R.; Allard, S.; Jia, D.T.; Windheim, J.; Cuello, A.C. BACE1 Inhibition by Microdose Lithium Formulation NP03 Rescues Memory Loss and Early Stage Amyloid Neuropathology. Transl. Psychiatry 2017, 7, e1190. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.C.; Lee, H.W.; Ryu, H.W.; Kang, M.-G.; Park, D.; Kim, S.H.; Cho, M.-L.; Oh, S.-R.; Kim, H. Selective Inhibition of Monoamine Oxidase A by Hispidol. Bioorg. Med. Chem. Lett. 2018, 28, 584–588. [Google Scholar] [CrossRef]
- Baek, S.C.; Park, M.H.; Ryu, H.W.; Lee, J.P.; Kang, M.-G.; Park, D.; Park, C.M.; Oh, S.-R.; Kim, H. Rhamnocitrin Isolated from Prunus Padus Var. Seoulensis: A Potent and Selective Reversible Inhibitor of Human Monoamine Oxidase A. Bioorg. Chem. 2019, 83, 317–325. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Son, S.-Y.; Ma, J.; Kondou, Y.; Yoshimura, M.; Yamashita, E.; Tsukihara, T. Structure of Human Monoamine Oxidase A at 2.2-A Resolution: The Control of Opening the Entry for Substrates/Inhibitors. Proc. Natl. Acad. Sci. USA 2008, 105, 5739–5744. [Google Scholar] [CrossRef] [Green Version]
- Binda, C.; Aldeco, M.; Mattevi, A.; Edmondson, D.E. Interactions of Monoamine Oxidases with the Antiepileptic Drug Zonisamide: Specificity of Inhibition and Structure of the Human Monoamine Oxidase B Complex. J. Med. Chem. 2011, 54, 909–912. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.E.J.; Dean, P.M. Three-Dimensional Hydrogen-Bond Geometry and Probability Information from a Crystal Survey. J. Comput. Aided Mol. Des. 1996, 10, 607–622. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera? A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Shigemoto, R.; Matsumoto, T.; Masuo, S.; Takaya, N. 5-Methylmellein Is a Novel Inhibitor of Fungal Sirtuin and Modulates Fungal Secondary Metabolite Production. J. Gen. Appl. Microbiol. 2018, 64, 240–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuno, T.; Oikawa, S.; Goto, T.; Sawai, K.; Shirahama, H.; Matsumoto, T. Structures and Phytotoxicity of Metabolites from Valsa Ceratosperma. Agric. Biol. Chem. 1986, 50, 997–1001. [Google Scholar] [CrossRef]
- Türk, H.; Yılmaz, M.; Tay, T.; Türk, A.Ö.; Kıvanç, M. Antimicrobial Activity of Extracts of Chemical Races of the Lichen Pseudevernia Furfuracea and Their Physodic Acid, Chloroatranorin, Atranorin, and Olivetoric Acid Constituents. Z. Für Nat. C 2006, 61, 499–507. [Google Scholar] [CrossRef]
- González-Tejero, M.R.; Martínez-Lirola, M.J.; Casares-Porcel, M.; Molero-Mesa, J. Three Lichens Used in Popular Medicine in Eastern Andalucia (Spain). Econ. Bot. 1995, 49, 96–98. [Google Scholar] [CrossRef]
- Geyikoglu, F.; Turkez, H.; Aslan, A. The Protective Roles of Some Lichen Species on Colloidal Bismuth Subcitrate Genotoxicity. Toxicol. Ind. Health 2007, 23, 487–492. [Google Scholar] [CrossRef]
- Ates, A.; Yildiz, A.; Yildiz, N.; Calimli, A. Heavy Metal Removal from Aqueous Solution by Pseudevernia Furfuracea (L.) Zopf. Ann. Chim. 2007, 97, 385–393. [Google Scholar] [CrossRef]
- Bari, A.; Rosso, A.; Minciardi, M.R.; Troiani, F.; Piervittori, R. Analysis of Heavy Metals in Atmospheric Particulates in Relation to Their Bioaccumulation in Explanted Pseudevernia Furfuracea Thalli. Environ. Monit. Assess. 2001, 69, 205–220. [Google Scholar] [CrossRef]
- Petrini, L.E.; Petrini, O. Morphological Studies in Rosellinia (Xylariaceae): The First Step towards a Polyphasic Taxonomy. Mycol. Res. 2005, 109, 569–580. [Google Scholar] [CrossRef]
- Pliego, C.; López-Herrera, C.; Ramos, C.; Cazorla, F.M. Developing Tools to Unravel the Biological Secrets of Rosellinia Necatrix, an Emergent Threat to Woody Crops: Tools to Study Rosellinia Necatrix. Mol. Plant Pathol. 2012, 13, 226–239. [Google Scholar] [CrossRef]
- Sharma, N.; Kushwaha, M.; Arora, D.; Jain, S.; Singamaneni, V.; Sharma, S.; Shankar, R.; Bhushan, S.; Gupta, P.; Jaglan, S. New Cytochalasin from Rosellinia sanctae-cruciana, an Endophytic Fungus of Albizia lebbeck. J. Appl. Microbiol. 2018, 125, 111–120. [Google Scholar] [CrossRef]
- Wittstein, K.; Cordsmeier, A.; Lambert, C.; Wendt, L.; Sir, E.B.; Weber, J.; Wurzler, N.; Petrini, L.E.; Stadler, M. Identification of Rosellinia Species as Producers of Cyclodepsipeptide PF1022 A and Resurrection of the Genus Dematophora as Inferred from Polythetic Taxonomy. Stud. Mycol. 2020, 96, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Srikitikulchai, P.; Srichomthong, K.; Lumyong, S. Oblongolides from the Endophytic Fungus Phomopsis Sp. BCC 9789. J. Nat. Prod. 2010, 73, 55–59. [Google Scholar] [CrossRef]
- Zheng, N.; Yao, F.; Liang, X.; Liu, Q.; Xu, W.; Liang, Y.; Liu, X.; Li, J.; Yang, R. A New Phthalide from the Endophytic Fungus Xylaria Sp. GDG-102. Nat. Prod. Res. 2018, 32, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Arora, D.; Sharma, N.; Singamaneni, V.; Sharma, V.; Kushwaha, M.; Abrol, V.; Guru, S.; Sharma, S.; Gupta, A.P.; Bhushan, S.; et al. Isolation and Characterization of Bioactive Metabolites from Xylaria Psidii, an Endophytic Fungus of the Medicinal Plant Aegle Marmelos and Their Role in Mitochondrial Dependent Apoptosis against Pancreatic Cancer Cells. Phytomed. Int. J. Phytother. Phytopharm. 2016, 23, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Yang, Z.-D.; Chen, X.; Zhou, S.-Y.; Yu, H.-T.; Sun, J.-Y.; Yao, X.-J.; Wang, Y.; Xue, H.-Y. Colletotrilactam A–D, Novel Lactams from Colletotrichum Gloeosporioides GT-7, a Fungal Endophyte of Uncaria Rhynchophylla. Fitoterapia 2016, 113, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, E.; Hossain, C.F.; Yamazaki, M. Monoamine Oxidase Inhibitors from a Lichen, Solorina crocea (L.) ACH. Jpn. J. Pharmacogn. 1991, 45, 159–162. [Google Scholar]
- Kinoshita, K.; Saito, D.; Koyama, K.; Takahashi, K.; Sato, Y.; Okuyama, E.; Fujimoto, H.; Yamazaki, M. Monoamine Oxidase Inhibitory Effects of Some Lichen Compounds and Their Synthetic Analogues. J.-Hattori Bot. Lab. 2002, 92, 277–284. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
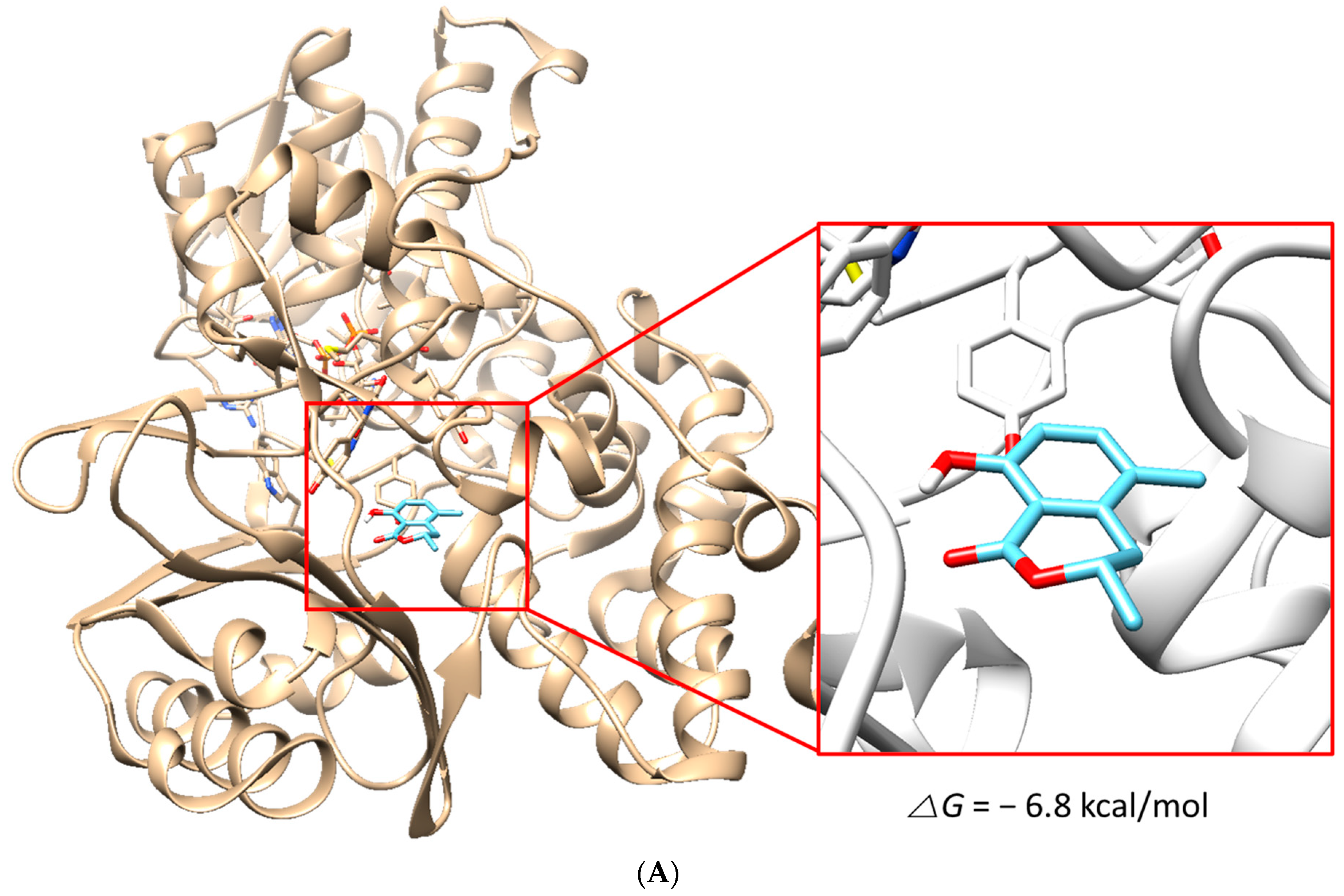{kind=link}
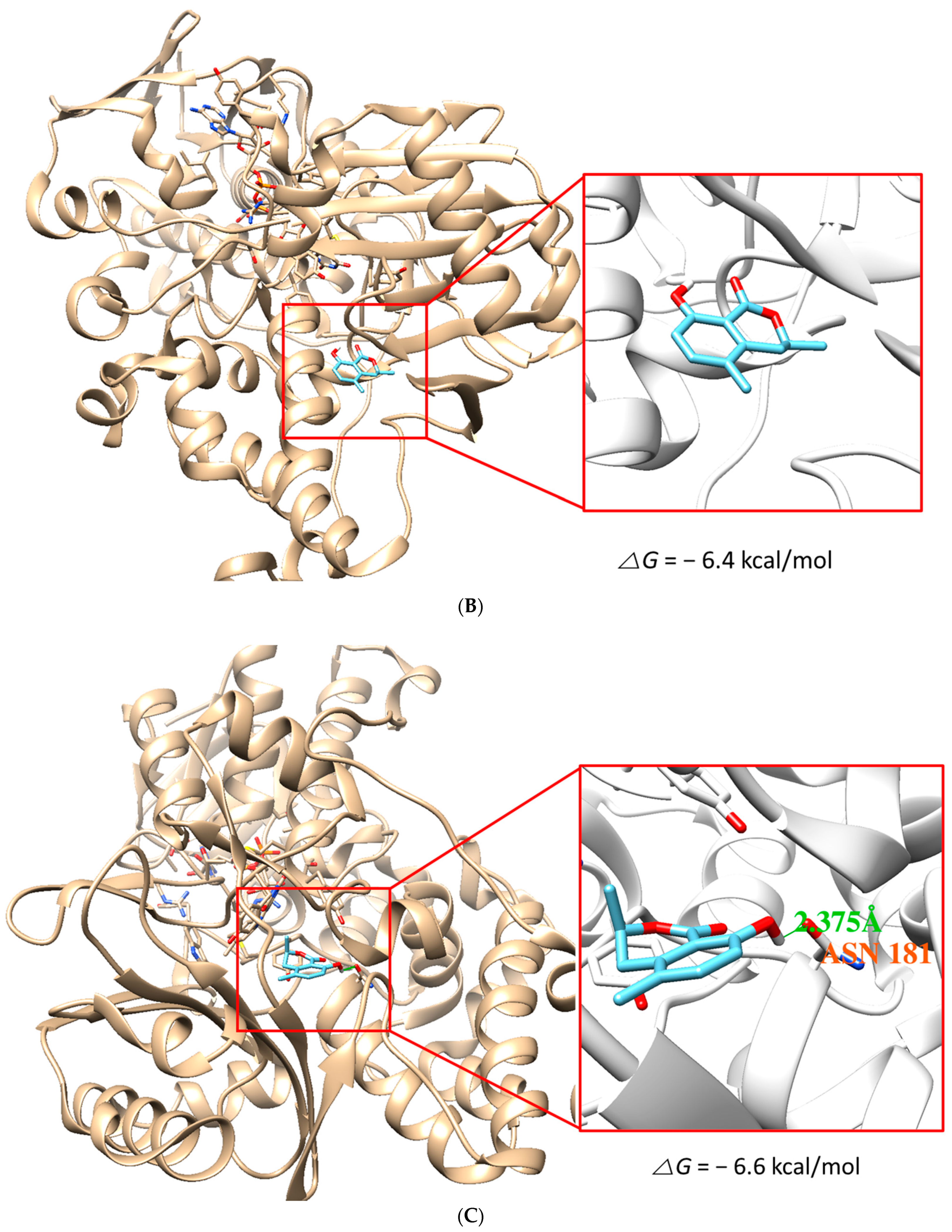{kind=link}
{kind=link}
| ELF No. | Residual Activity at 20 µg/mL (%) | |
|---|---|---|
| hMAO-A | hMAO-B | |
| 56 | 51.1 ± 2.2 | 34.7 ± 1.7 |
| 62 | 50.4 ± 1.1 | 43.7 ± 2.0 |
| 68 | 26.2 ± 5.8 | 61.8 ± 4.3 |
| 71 | 43.6 ± 2.5 | 72.7 ± 2.5 |
| 82 | 58.3 ± 3.2 | 52.2 ± 1.2 |
| 100 | 42.3 ± 1.0 | 43.9 ± 5.7 |
| 116 | 50.0 ± 4.6 | 56.4 ± 2.3 |
| Residual Activity at 10 µM (%) | ||||||
|---|---|---|---|---|---|---|
| hMAO-A | hMAO-B | AChE | BChE | BACE1 | ||
| 5MM | 35.2 ± 0.9 | 44.5 ± 2.6 | 71.4 ± 2.1 | 92.5 ± 2.5 | 93.3 ± 2.7 | |
| IC50 (µM) | SI | |||||
| hMAO-A | hMAO-B | AChE | BChE | BACE1 | ||
| 5MM | 5.31 ± 0.24 | 9.15 ± 0.92 | 27.07 ± 0.04 | >40 | >40 | 1.72 |
| Toloxatone | 1.08 ± 0.03 | |||||
| Clorgyline | 0.007 ± 0.001 | |||||
| Lazabemide | 0.063 ± 0.015 | |||||
| Pargyline | 0.028 ± 0.004 | |||||
| Donepezil | 0.009 ± 0.002 | 0.180 ± 0.004 | ||||
| Quercetin | 20.46 ± 0.60 | |||||
| Compound | GI Absorption | BBB Permeant | P-gp Substrate | CYP1A2 Inhibitor | CYP2C19 Inhibitor | CYP2C9 Inhibitor | CYP2D6 Inhibitor | CYP3A4 Inhibitor | Log Kp (Skin Permeation) |
|---|---|---|---|---|---|---|---|---|---|
| 5MM | High | Yes | No | Yes | No | No | No | No | −5.48 cm/s |
| Compound | Mw(g/mol) | cLog P | HBD | HBA | TPSA (Å2) | RB | Lipinski Violations |
|---|---|---|---|---|---|---|---|
| 5MM | 192.21 | 2.19 | 1 | 3 | 46.53 | No | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, G.-S.; Lee, E.-Y.; Kang, M.-G.; Nam, S.-J.; Park, D.; Kim, H. (S)-5-Methylmellein Isolated from an Endogenous Lichen Fungus Rosellinia corticium as a Potent Inhibitor of Human Monoamine Oxidase A. Processes 2022, 10, 166. https://doi.org/10.3390/pr10010166
Jeong G-S, Lee E-Y, Kang M-G, Nam S-J, Park D, Kim H. (S)-5-Methylmellein Isolated from an Endogenous Lichen Fungus Rosellinia corticium as a Potent Inhibitor of Human Monoamine Oxidase A. Processes. 2022; 10(1):166. https://doi.org/10.3390/pr10010166
Chicago/Turabian StyleJeong, Geum-Seok, Eun-Young Lee, Myung-Gyun Kang, Sang-Jip Nam, Daeui Park, and Hoon Kim. 2022. "(S)-5-Methylmellein Isolated from an Endogenous Lichen Fungus Rosellinia corticium as a Potent Inhibitor of Human Monoamine Oxidase A" Processes 10, no. 1: 166. https://doi.org/10.3390/pr10010166