Regulatory Role of Orexin in the Antistress Effect of “Press Tack Needle” Acupuncture Treatment

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs
2.3. PTN
2.4. Influence of the PTN Treatment on the Plasma Orexin A Level
2.5. Antistress Effect of the PTN Treatment
2.5.1. Groups and Stress Procedure
2.5.2. Aggressive Behavior Test
2.5.3. Plasma Orexin A and Corticosterone Levels
2.5.4. Immunofluorescence of Orexin A
2.6. Involvement of the Orexin Receptor in Aggressive Behavior
2.7. Statistical Analysis
3. Results
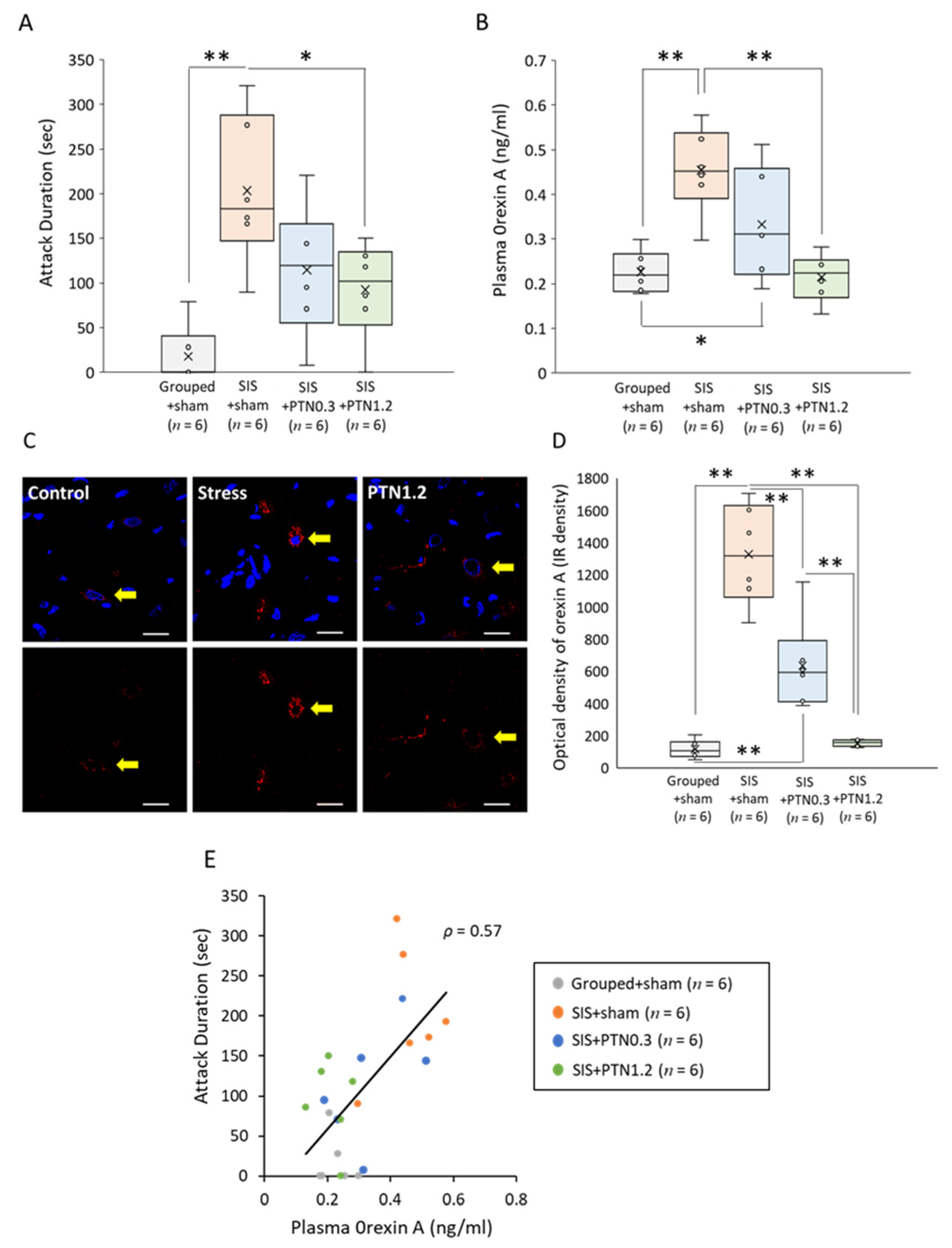
3.1. Influence of PTN on the Plasma Orexin A Level
3.2. A Level Aggressive Behavior Test
3.3. Plasma Level and Immunofluorescent Staining of Orexin A
3.4. Consideration of the Correlation between Attack Duration and Orexin A Level
3.5. Plasma Corticosterone Level
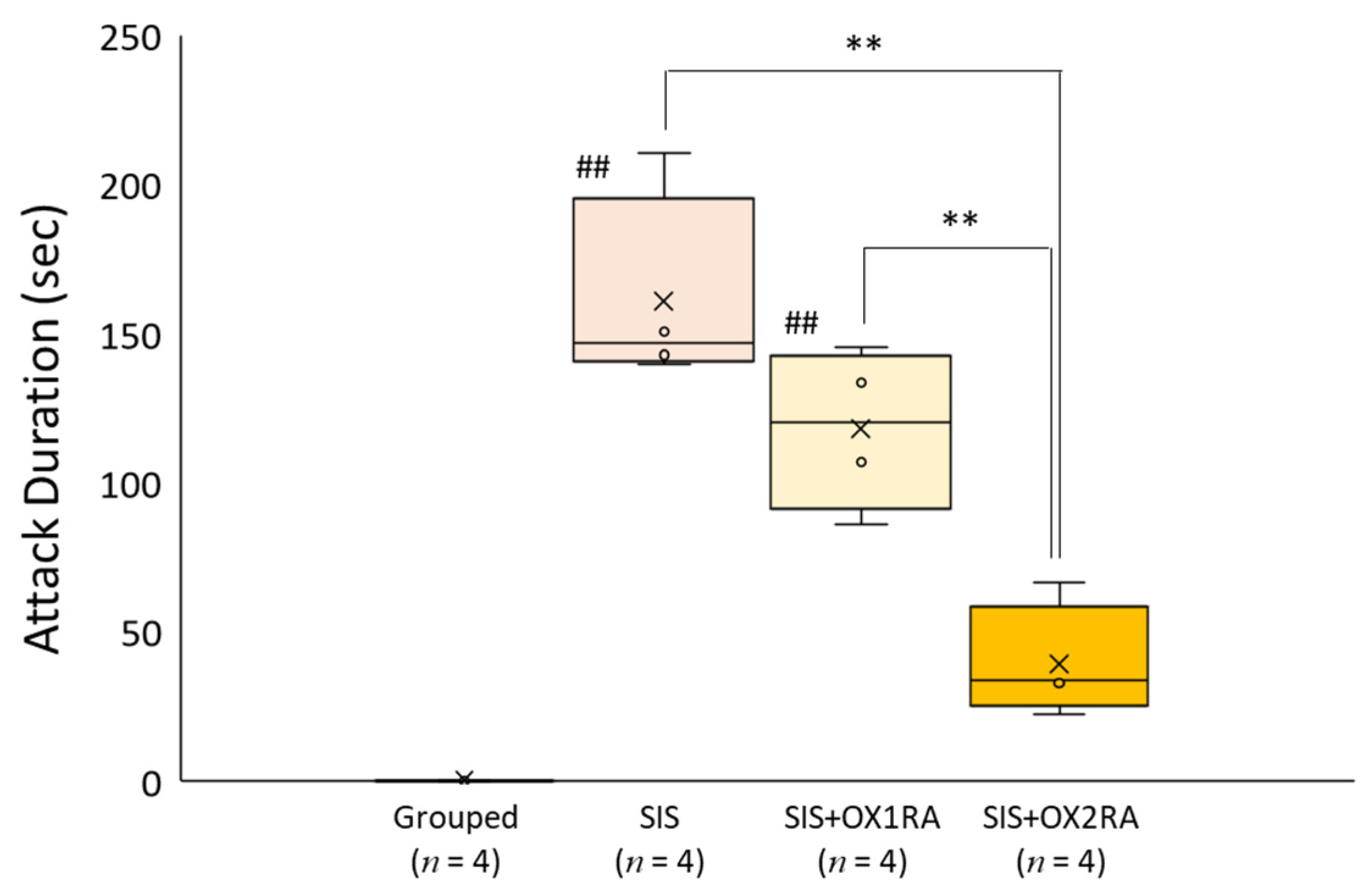
3.6. Involvement of Orexin Receptor in Aggressive Behavior
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [CrossRef]
- Gerra, G.; Zaimovic, A.; Mascetti, G.G.; Gardini, S.; Zambelli, U.; Timpano, M.; Raggi, M.A.; Brambilla, F. Neuroendocrine responses to experimentally-induced psychological stress in healthy humans. Psychoneuroendocrinology 2001, 26, 91–107. [Google Scholar] [CrossRef]
- Peyron, C.; Tighe, D.K.; van den Pol, A.N.; de Lecea, L.; Heller, H.C.; Sutcliffe, J.G.; Kilduff, T.S. Neurons containing hypocretin (orexin) project to multiple neuronal systems. J. Neurosci. 1998, 18, 9996–10015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef] [Green Version]
- Chemelli, R.M.; Willie, J.T.; Sinton, C.M.; Elmquist, J.K.; Scammell, T.; Lee, C.; Richardson, J.A.; Williams, S.C.; Xiong, Y.; Kisanuki, Y.; et al. Narcolepsy in orexin knockout mice: Molecular genetics of sleep regulation. Cell 1999, 98, 437–451. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Faraco, J.; Li, R.; Kadotani, H.; Rogers, W.; Lin, X.; Qiu, X.; de Jong, P.J.; Nishino, S.; Mignot, E. The Sleep Disorder Canine Narcolepsy Is Caused by a Mutation in the Hypocretin (Orexin) Receptor 2 Gene. Cell 1999, 98, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Yokobori, E.; Kojima, K.; Azuma, M.; Kang, K.S.; Maejima, S.; Uchiyama, M.; Matsuda, K. Stimulatory effect of intracerebroventricular administration of orexin A on food intake in the zebrafish, Danio rerio. Peptides 2011, 32, 1357–1362. [Google Scholar] [CrossRef]
- Perez-Leighton, C.E.; Boland, K.; Teske, J.A.; Billington, C.; Kotz, C.M. Behavioral responses to orexin, orexin receptor gene expression, and spontaneous physical activity contribute to individual sensitivity to obesity. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E865–E874. [Google Scholar] [CrossRef] [Green Version]
- Bingham, S.; Davey, P.T.; Babbs, A.J.; Irving, E.A.; Sammons, M.J.; Wyles, M.; Jeffrey, P.; Cutler, L.; Riba, I.; Johns, A.; et al. Orexin-A, an hypothalamic peptide with analgesic properties. Pain 2001, 92, 81–90. [Google Scholar] [CrossRef]
- Yan, J.A.; Ge, L.; Huang, W.; Song, B.; Chen, X.W.; Yu, Z.P. Orexin affects dorsal root ganglion neurons: A mechanism for regulating the spinal nociceptive processing. Physiol. Res. 2008, 57, 797–800. [Google Scholar] [PubMed]
- Samson, W.K.; Gosnell, B.; Chang, J.K.; Resch, Z.T.; Murphy, T.C. Cardiovascular regulatory actions of the hypocretins in brain. Brain Res. 1999, 831, 248–253. [Google Scholar] [CrossRef]
- Shirasaka, T.; Nakazato, M.; Matsukura, S.; Takasaki, M.; Kannan, H. Sympathetic and cardiovascular actions of orexins in conscious rats. Am. J. Physiol. 1999, 277, R1780–R1785. [Google Scholar] [CrossRef]
- Williams, R.H.; Jensen, L.T.; Verkhratsky, A.; Fugger, L.; Burdakov, D. Control of hypothalamic orexin neurons by acid and CO2. Proc. Natl. Acad. Sci. USA 2007, 104, 10685–10690. [Google Scholar] [CrossRef] [Green Version]
- Wayner, M.J.; Armstrong, D.L.; Phelix, C.F.; Oomura, Y. Orexin-A (Hypocretin-1) and leptin enhance LTP in the dentate gyrus of rats in vivo. Peptides 2004, 25, 991–996. [Google Scholar] [CrossRef]
- Yang, L.; Zou, B.; Xiong, X.; Pascual, C.; Xie, J.; Malik, A.; Xie, J.; Sakurai, T.; Xie, X.S. Hypocretin/orexin neurons contribute to hippocampus-dependent social memory and synaptic plasticity in mice. J. Neurosci. 2013, 33, 5275–5284. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, A.; Muraki, Y.; Tsujino, N.; Goto, K.; Sakurai, T. Regulation of orexin neurons by the monoaminergic and cholinergic systems. Biochem. Biophys. Res. Commun. 2003, 303, 120–129. [Google Scholar] [CrossRef]
- Kok, S.W.; Roelfsema, F.; Overeem, S.; Lammers, G.J.; Strijers, R.L.; Frolich, M.; Meinders, A.E.; Pijl, H. Dynamics of the pituitary-adrenal ensemble in hypocretin-deficient narcoleptic humans: Blunted basal adrenocorticotropin release and evidence for normal time-keeping by the master pacemaker. J. Clin. Endocrinol. Metab. 2002, 87, 5085–5091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ida, T.; Nakahara, K.; Murakami, T.; Hanada, R.; Nakazato, M.; Murakami, N. Possible involvement of orexin in the stress reaction in rats. Biochem. Biophys. Res. Commun. 2000, 270, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Saito, T.; Ohiwa, N.; Tateoka, M.; Deocaris, C.C.; Fujikawa, T.; Soya, H. Inhibitory effects of an orexin-2 receptor antagonist on orexin A- and stress-induced ACTH responses in conscious rats. Neurosci. Res. 2007, 57, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Katahira, H.; Sunagawa, M.; Watanabe, D.; Kanada, Y.; Katayama, A.; Yamauchi, R.; Takashima, M.; Ishikawa, S.; Hisamitsu, T. Antistress effects of Kampo medicine “Yokukansan” via regulation of orexin secretion. Neuropsychiatr. Dis. Treat. 2017, 13, 863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuru, M.; Ueta, Y.; Serino, R.; Nakazato, M.; Yamamoto, Y.; Shibuya, I.; Yamashita, H. Centrally administered orexin/hypocretin activates HPA axis in rats. Neuroreport 2000, 11, 1977–1980. [Google Scholar] [CrossRef]
- Malendowicz, L.K.; Hochol, A.; Ziolkowska, A.; Nowak, M.; Gottardo, L.; Nussdorfer, G.G. Prolonged orexin administration stimulates steroid-hormone secretion, acting directly on the rat adrenal gland. Int. J. Mol. Med. 2001, 7, 401–404. [Google Scholar] [CrossRef]
- Nemoto, T.; Toyoshima-Aoyama, F.; Ueda, Y.; Ohba, T.; Yanagita, T.; Watanabe, H.; Shirasaka, T.; Tsuneyoshi, I.; Ishida, Y.; Hirota, K.; et al. Involvement of the orexin system in adrenal sympathetic regulation. Pharmacology 2013, 91, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Simon, G.E.; Cherkin, D.C.; Sherman, K.J.; Eisenberg, D.M.; Deyo, R.A.; Davis, R.B. Mental health visits to complementary and alternative medicine providers. Gen. Hosp. Psychiatry 2004, 26, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, K. Acupuncture therapy for psychiatric illness. Int. Rev. Neurobiol. 2013, 111, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Eshkevari, L.; Permaul, E.; Mulroney, S.E. Acupuncture blocks cold stress-induced increases in the hypothalamus-pituitary-adrenal axis in the rat. J. Endocrinol. 2013, 217, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Park, H.J.; Chae, Y.; Kim, J.W.; Lee, H.; Chung, J.H. Effect of acupuncture on hypothalamic-pituitary-adrenal system in maternal separation rats. Cell. Mol. Neurobiol. 2011, 31, 1123–1127. [Google Scholar] [CrossRef]
- Lee, B.; Shim, I.; Lee, H.J.; Yang, Y.; Hahm, D.H. Effects of acupuncture on chronic corticosterone-induced depression-like behavior and expression of neuropeptide Y in the rats. Neurosci. Lett. 2009, 453, 151–156. [Google Scholar] [CrossRef]
- Yang, C.H.; Lee, B.B.; Jung, H.S.; Shim, I.; Roh, P.U.; Golden, G.T. Effect of electroacupuncture on response to immobilization stress. Pharmacol. Biochem. Behav. 2002, 72, 847–855. [Google Scholar] [CrossRef]
- Han, S.H.; Yoon, S.H.; Cho, Y.W.; Kim, C.J.; Min, B.I. Inhibitory effects of electroacupuncture on stress responses evoked by tooth-pulp stimulation in rats. Physiol. Behav. 1999, 66, 217–222. [Google Scholar] [CrossRef]
- Tanahashi, N.; Takagi, K.; Amagasu, N.; Wang, G.; Mizuno, K.; Kawanoguchi, J.; Yi, G.; Ishida, T. Effect of acupuncture stimulation on rats with depression induced by water-immersion stress. Neurosci. Lett. 2016, 618, 99–103. [Google Scholar] [CrossRef]
- Kaneko, Y.; Furuya, E.; Sakamoto, A. The effects of press tack needle treatment on muscle soreness after triathlon race—Sham controlled study. JAM 2009, 1, 22–30. [Google Scholar]
- Furuya, E.; Nayuki, T.; Yakame, M.; Furuumi, H.; Shinohara, R.; Nimura, R.; Sakamoto, A. The effects of press tack needle treatment on upper back muscle stiffness—Comparative study using sham needle. JAM 2009, 1, 12–21. [Google Scholar]
- Schröder, S.; Meyer-Hamme, G.; Friedemann, T.; Kirch, S.; Hauck, M.; Plaetke, R.; Friedrichs, S.; Gulati, A.; Briem, D. Immediate pain relief in adhesive capsulitis by acupuncture—A randomized controlled double-blinded study. Pain Med. 2017, 18, 2235–2247. [Google Scholar] [CrossRef]
- Ruengwongroj, P.; Muengtaweepongsa, S.; Patumanond, J.; Phinyo, P. Effectiveness of press needle treatment and electroacupuncture in patients with postherpetic neuralgia: A matched propensity score analysis. Complement. Ther. Clin. Pract. 2020, 40, 101202. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, S.; Hagihara, A.; Kanda, R.; Mukaino, Y.; Nobutomo, K. Applicability of press needles to a double-blind trial: A randomized, double-blind, placebo-controlled trial. Clin. J. Pain 2009, 25, 438–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, S.; Kamiya, T.; Watanabe, M.; Hirano, A.; Matsuda, A.; Monma, Y.; Numata, T.; Kusuyama, H.; Yaegashi, N. Report on disaster medical operations with acupuncture/massage therapy after the great East Japan earthquake. Integr. Med. Insights 2012, 7, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Staples, L.G.; Cornish, J.L. The orexin-1 receptor antagonist SB-334867 attenuates anxiety in rats exposed to cat odor but not the elevated plus maze: An investigation of Trial 1 and Trial 2 effects. Horm. Behav. 2014, 65, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Mould, R.; Brown, J.; Marshall, F.; Langmead, C. Binding kinetics differentiates functional antagonism of orexin-2 receptor ligands. Br. J. Pharmacol. 2014, 171, 351–363. [Google Scholar] [CrossRef]
- Duncan, M.J.; Franklin, K.M.; Peng, X.; Yun, C.; Legan, S.J. Circadian rhythm disruption by a novel running wheel: Roles of exercise and arousal in blockade of the luteinizing hormone surge. Physiol. Behav. 2014, 131, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Thoa, N.B.; Tizabi, Y.; Jacobowitz, D.M. The effect of isolation on catecholamine concentration and turnover in discrete areas of the rat brain. Brain Res. 1977, 131, 259–269. [Google Scholar] [CrossRef]
- Niesink, R.J.; Van Ree, J.M. Short-term isolation increases social interactions of male rats: A parametric analysis. Physiol. Behav. 1982, 29, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Varlinskaya, E.I.; Spear, L.P.; Spear, N.E. Social behavior and social motivation in adolescent rats: Role of housing conditions and partner’s activity. Physiol. Behav. 1999, 67, 475–482. [Google Scholar] [CrossRef]
- Gentsch, C.; Lichtsteiner, M.; Frischknecht, H.R.; Feer, H.; Siegfried, B. Isolation-induced locomotor hyperactivity and hypoalgesia in rats are prevented by handling and reversed by resocialization. Physiol. Behav. 1988, 43, 13–16. [Google Scholar] [CrossRef]
- Yamada, C.; Saegusa, Y.; Nahata, M.; Sadakane, C.; Hattori, T.; Takeda, H. Influence of Aging and Gender Differences on Feeding Behavior and Ghrelin-Related Factors during Social Isolation in Mice. PLoS ONE 2015, 10, e0140094. [Google Scholar] [CrossRef] [PubMed]
- Huong, N.T.; Murakami, Y.; Tohda, M.; Watanabe, H.; Matsumoto, K. Social isolation stress-induced oxidative damage in mouse brain and its modulation by majonoside-R2, a Vietnamese ginseng saponin. Biol. Pharm. Bull. 2005, 28, 1389–1393. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, S.; Kirouac, G.J. Role of the orexin (hypocretin) system in contextual fear conditioning in rats. Behav. Brain Res. 2017, 316, 47–53. [Google Scholar] [CrossRef]
- Steven, K.J.A.; William, P.C.C. Clinical Introduction to Medical Acupuncture; Thieme: New York, NY, USA, 2006; p. 88. [Google Scholar]
- Wang, G.; Jia, S.; Li, H.; Wang, Z.; Tian, Y.; Zhang, W. Changes of Blood Flux at BL21 and Points along BL Meridian Resulted from Acupuncture or Moxibustion: Case Cross Design Study. Evid. Based Complement. Alternat. Med. 2017, 2017, 8237580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Song, Y.; Bao, T.; Yu, M.; Xu, M.; Guo, Y.; Wang, Y.; Zhang, C.; Zhao, B. Antidepressant-like effects of acupuncture involved the ERK signaling pathway in rats. BMC Complement. Altern. Med. 2016, 16, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, A.; Weiland, C.; Enck, P.; Joos, S.; Streitberger, K.; Maser-Gluth, C.; Zipfel, S.; Bagheri, S.; Herzog, W.; Friederich, H.C. Neuroendocrinological effects of acupuncture treatment in patients with irritable bowel syndrome. Complement. Ther. Med. 2007, 15, 255–263. [Google Scholar] [CrossRef]
- Zhang, X.F.; Zhu, J.; Geng, W.Y.; Zhao, S.J.; Jiang, C.W.; Cai, S.R.; Cheng, M.; Zhou, C.Y.; Liu, Z.B. Electroacupuncture at Feishu (BL13) and Zusanli (ST36) down-regulates the expression of orexins and their receptors in rats with chronic obstructive pulmonary disease. J. Integr. Med. 2014, 12, 417–424. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, B.; Zhang, L.; Liu, S. Electroacupuncture suppresses morphine reward-seeking behavior: Lateral hypothalamic orexin neurons implicated. Neurosci. Lett. 2017, 661, 84–89. [Google Scholar] [CrossRef]
- Feng, X.M.; Mi, W.L.; Xia, F.; Mao-Ying, Q.L.; Jiang, J.W.; Xiao, S.; Wang, Z.F.; Wang, Y.Q.; Wu, G.C. Involvement of spinal orexin A in the electroacupuncture analgesia in a rat model of post-laparotomy pain. BMC Complement. Altern. Med. 2012, 12, 225. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Lee, H.J.; Lee, M.T.; Wu, Y.T.; Lee, Y.H.; Hwang, L.L.; Hung, M.S.; Zimmer, A.; Mackie, K.; Chiou, L.C. Median nerve stimulation induces analgesia via orexin-initiated endocannabinoid disinhibition in the periaqueductal gray. Proc. Natl. Acad. Sci. USA 2018, 115, E10720–E10729. [Google Scholar] [CrossRef] [Green Version]
- Flanigan, M.E.; Aleyasin, H.; Li, L.; Burnett, C.J.; Chan, K.L.; LeClair, K.B.; Lucas, E.K.; Matikainen-Ankney, B.; Durand-de Cuttoli, R.; Takahashi, A.; et al. Orexin signaling in GABAergic lateral habenula neurons modulates aggressive behavior in male mice. Nat. Neurosci. 2020, 23, 638–650. [Google Scholar] [CrossRef]
- Murgatroyd, C.A.; Taliefar, M.; Bradburn, S.; Carini, L.M.; Babb, J.A.; Nephew, B.C. Social stress during lactation, depressed maternal care, and neuropeptidergic gene expression. Behav. Pharmacol. 2015, 26, 642–653. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wang, X.Y.; Shi, H.; Bai, W.Z.; Cheng, B.; Su, Y.S.; Yu, X.C.; Jing, X.H.; Zhu, B. Cutaneous neurogenic inflammation in the sensitized acupoints induced by gastric mucosal injury in rats. BMC Complement. Altern. Med. 2017, 17, 141. [Google Scholar] [CrossRef] [Green Version]
- Ding, N.; Jiang, J.; Qin, P.; Wang, Q.; Hu, J.; Li, Z. Mast cells are important regulator of acupoint sensitization via the secretion of tryptase, 5-hydroxytryptamine, and histamine. PLoS ONE 2018, 13, e0194022. [Google Scholar] [CrossRef]
- Yao, W.; Yang, H.; Yin, N.; Ding, G. Mast cell-nerve cell interaction at acupoint: Modeling mechanotransduction pathway induced by acupuncture. Int. J. Biol. Sci. 2014, 10, 511. [Google Scholar] [CrossRef] [PubMed]
- Goldman, N.; Chen, M.; Fujita, T.; Xu, Q.; Peng, W.; Liu, W.; Jensen, T.K.; Pei, Y.; Wang, F.; Han, X.; et al. Adenosine A1 receptors mediate local anti-nociceptive effects of acupuncture. Nat. Neurosci. 2010, 13, 883–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Wang, X.; Xing, B.; Yang, H.; Sa, Z.; Zhang, D.; Yao, W.; Yin, N.; Xia, Y.; Ding, G. Critical roles of TRPV2 channels, histamine H1 and adenosine A1 receptors in the initiation of acupoint signals for acupuncture analgesia. Sci. Rep. 2018, 8, 6523. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | Types of Stressor | Ways of Stimulation | Acupuncture Points | Results Effects on the HPA/SAM Axis |
|---|---|---|---|---|
| Eshkevari et al. [26] | cold temperature | electroacupuncture (10 Hz) | ST36 | CRH↓ ACTH↓ CORT↓ NPY↓ |
| Park et al. [27] | maternal separation | twirling | HT7 | ACTH↓ CORT↓ |
| Lee et al. [28] | repeated injection of CORT | needle retention | PC6 | CRH↓ |
| Yang et al. [29] | forced immobilization | electroacupuncture (3 Hz) | HT3 and PC6 | NA↓ A↓ |
| Han et al. [30] | tooth-pulp stimulation | electroacupuncture (3 Hz) | LI4 | ACTH↓ CORT↓ NA↓ DA↓ |
| Tanahashi et al. [31] | water-immersion | needle retention | GV20 and Ex-HN3 | CORT↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujiwara, A.; Tsukada, M.; Ikemoto, H.; Izuno, T.; Hattori, S.; Okumo, T.; Hisamitsu, T.; Sunagawa, M. Regulatory Role of Orexin in the Antistress Effect of “Press Tack Needle” Acupuncture Treatment. Healthcare 2021, 9, 503. https://doi.org/10.3390/healthcare9050503
Fujiwara A, Tsukada M, Ikemoto H, Izuno T, Hattori S, Okumo T, Hisamitsu T, Sunagawa M. Regulatory Role of Orexin in the Antistress Effect of “Press Tack Needle” Acupuncture Treatment. Healthcare. 2021; 9(5):503. https://doi.org/10.3390/healthcare9050503
Chicago/Turabian StyleFujiwara, Aki, Mana Tsukada, Hideshi Ikemoto, Takuji Izuno, Satoshi Hattori, Takayuki Okumo, Tadashi Hisamitsu, and Masataka Sunagawa. 2021. "Regulatory Role of Orexin in the Antistress Effect of “Press Tack Needle” Acupuncture Treatment" Healthcare 9, no. 5: 503. https://doi.org/10.3390/healthcare9050503