
Dynamics in the Reduced Mean-Field Model of Neuron–Glial Interaction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Model
2.1. Astrocytic Dynamics
2.2. Neuron–Glial Interaction
2.3. Reduced Model
3. Methods
3.1. Two-Parameter Maps and the Inheritance Scheme
3.2. Map of Lyapunov Spectrum
3.3. Map of Population Activity
3.4. Detection of Multistability
3.5. Poincare Map and One-Dimensional Analysis
4. Results
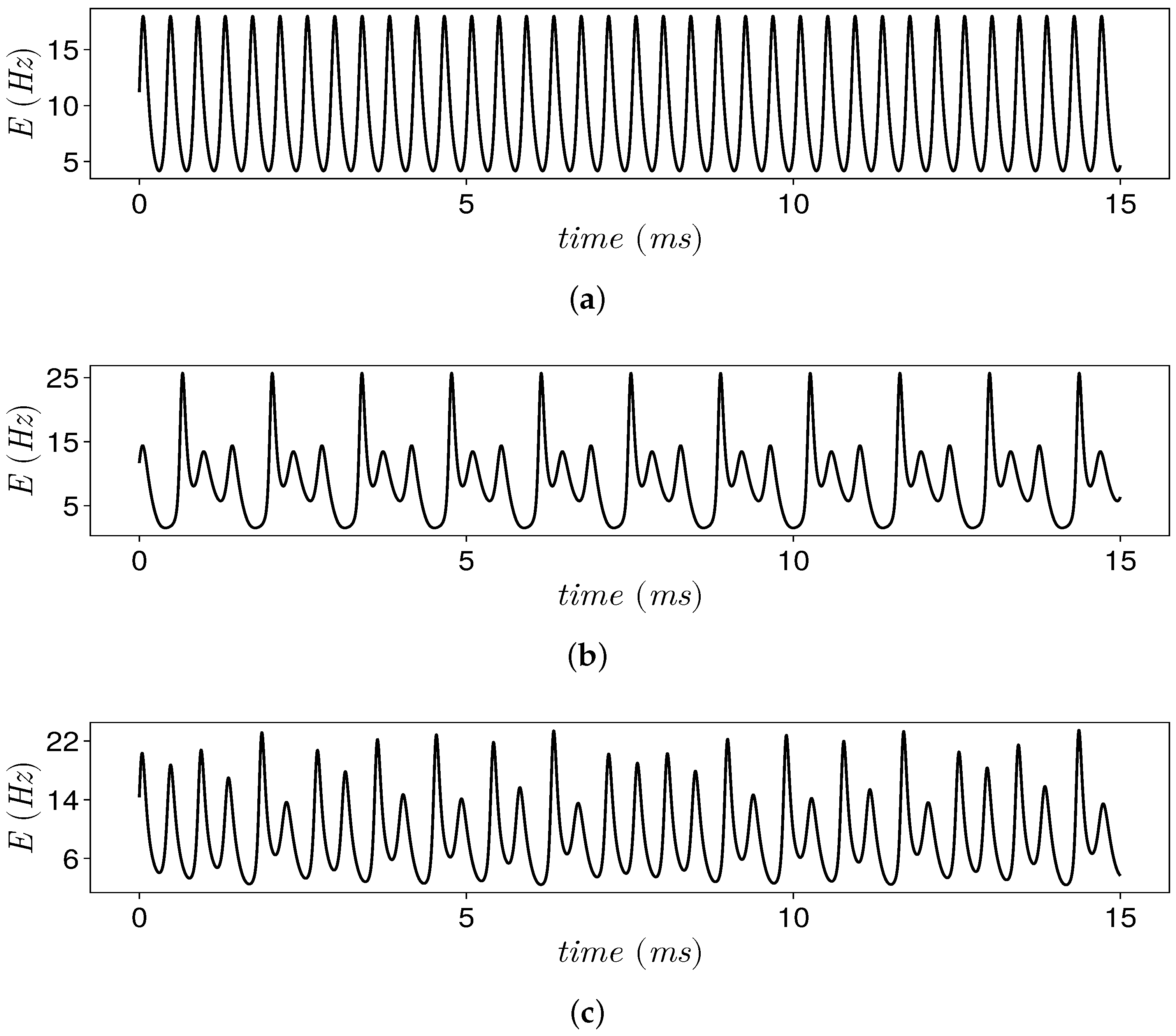
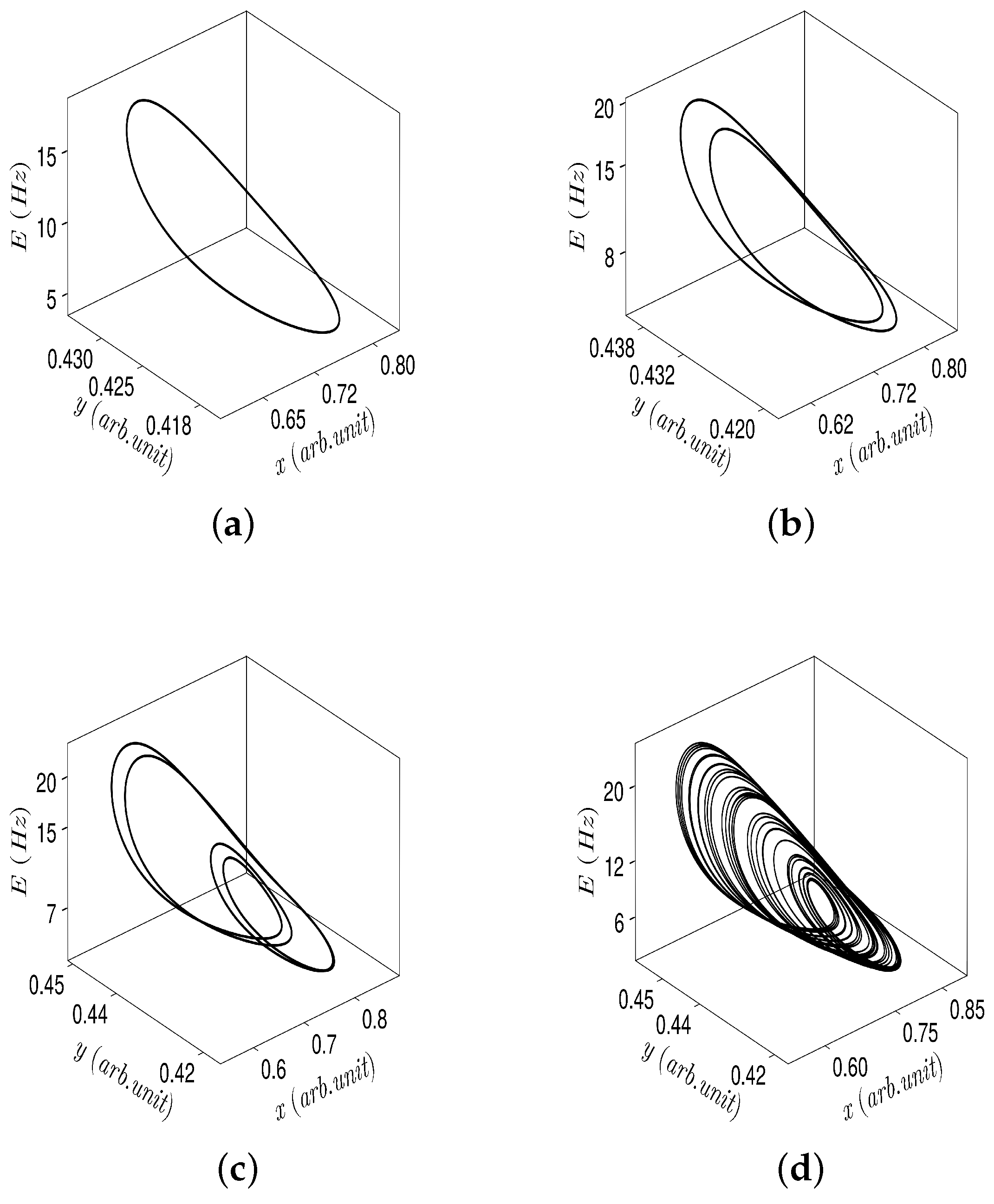
4.1. Patterns of Population Activity
4.2. Bifurcation Analysis
4.3. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varela, F.; Lachaux, J.; Rodriguez, E.; Martinerie, J. The brainweb: Phase synchronization and large-scale integration. Nat. Rev. Neurosci. 2001, 2, 229–239. [Google Scholar] [CrossRef]
- Uhlhaas, P.; Singer, W. Neural synchrony in brain disorders: Relevance for cognitive dysfunctions and pathophysiology. Neuron 2006, 52, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Lisman, J. Bursts as a unit of neural information: Making unreliable synapses reliable. Trends Neurosci. 1997, 20, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Izhikevich, E.; Desai, N.; Walcott, E.; Hoppensteadt, F. Bursts as a unit of neural information: Selective communication via resonance. Trends Neurosci. 2003, 26, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Gabbiani, F.; Metzner, W.; Wessel, R.; Koch, C. From stimulus encoding to feature extraction in weakly electric fish. Nature 1996, 384, 564–567. [Google Scholar] [CrossRef]
- Oswald, A.; Chacron, M.; Doiron, B.; Bastian, J.; Maler, L. Parallel processing of sensory input by bursts and isolated spikes. J. Neurosci. 2004, 24, 4351–4362. [Google Scholar] [CrossRef]
- Lesica, N.; Stanley, G. Encoding of natural scene movies by tonic and burst spikes in the lateral geniculate nucleus. J. Neurosci. 2004, 24, 10731–10740. [Google Scholar] [CrossRef]
- Reinagel, P.; Godwin, D.; Sherman, S.; Koch, C. Encoding of visual information by LGN bursts. J. Neurophysiol. 1999, 81, 2558–2569. [Google Scholar] [CrossRef]
- Segev, R.; Baruchi, I.; Hulata, E.; Ben-Jacob, E. Hidden neuronal correlations in cultured networks. Phys. Rev. Lett. 2004, 92, 118102. [Google Scholar] [CrossRef]
- Hulata, E.; Baruchi, I.; Segev, R.; Shapira, Y.; Ben-Jacob, E. Self-regulated complexity in cultured neuronal networks. Phys. Rev. Lett. 2004, 92, 198105. [Google Scholar] [CrossRef]
- Avoli, M.; Olivier, A. Bursting in human epileptogenic neocortex is depressed by an N-methyl-D-aspartate antagonist. Neurosci. Lett. 1987, 76, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Hofer, K.T.; Kandracs, Á.; Tóth, K.; Hajnal, B.; Bokodi, V.; Toth, E.Z.; Erőss, L.; Entz, L.; Bago, A.G.; Fabo, D.; et al. Bursting of excitatory cells is linked to interictal epileptic discharge generation in humans. Sci. Rep. 2022, 12, 6280. [Google Scholar] [CrossRef]
- Ioannou, P.; Foster, D.L.; Sander, J.W.; Dupont, S.; Gil-Nagel, A.; Dragon O’Flaherty, E.; Alvarez-Baron, E.; Medjedovic, J. The burden of epilepsy and unmet need in people with focal seizures. Brain Behav. 2022, 12, e2589. [Google Scholar] [CrossRef] [PubMed]
- Lehnertz, K. Epilepsy: Extreme Events in the Human Brain. In Extreme Events in Nature and Society; Albeverio, S., Jentsch, V., Kantz, H., Eds.; The Frontiers Collection; Springer: Berlin/Heidelberg, Germany, 2006; pp. 123–143. ISBN 978-3-540-28610-3. [Google Scholar]
- Dingledine, R.; Varvel, N.H.; Dudek, F.E. When and How Do Seizures Kill Neurons, and Is Cell Death Relevant to Epileptogenesis? Adv. Exp. Med. Biol. 2014, 813, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Vezzani, A.; Najjar, S.; De Lanerolle, N.C.; Rogawski, M.A. Glia and epilepsy: Excitability and inflammation. Trends Neurosci. 2013, 36, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Porter, B.E.; Friedman, A.; Kaufer, D. A potential role for glia-derived extracellular matrix remodeling in postinjury epilepsy. J. Neurosci. Res. 2016, 94, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.C.; Tewari, B.P.; Chaunsali, L.; Sontheimer, H. Neuron–glia interactions in the pathophysiology of epilepsy. Nat. Rev. Neurosci. 2019, 20, 282–297. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Pristov, J.B.; Nobili, P.; Nikolić, L. Can glial cells save neurons in epilepsy? Neural Regen. Res. 2022, 18, 1417–1422. [Google Scholar] [CrossRef]
- Tian, G.; Azmi, H.; Takano, T.; Xu, Q.; Peng, W.; Lin, J.; Oberheim, N.; Lou, N.; Wang, X.; Zielke, H. An astrocytic basis of epilepsy. Nat. Med. 2005, 11, 973–981. [Google Scholar] [CrossRef]
- Seifert, G.; Steinhäuser, C. Neuron–astrocyte signaling and epilepsy. Exp. Neurol. 2013, 244, 4–10. [Google Scholar] [CrossRef]
- Henning, L.; Unichenko, P.; Bedner, P.; Steinhauser, C.; Henneberger, C. Overview article astrocytes as initiators of epilepsy. Neurochem. Res. 2023, 48, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Crunfli, F.; Carregari, V.C.; Veras, F.P.; Silva, L.S.; Nogueira, M.H.; Antunes, A.S.L.M.; Vendramini, P.H.; Valença, A.G.F.; Brandão-Teles, C.; da Silva Zuccoli, G.; et al. Morphological, cellular, and molecular basis of brain infection in COVID-19 patients. Proc. Natl. Acad. Sci. USA 2022, 119, e2200960119. [Google Scholar] [CrossRef] [PubMed]
- Hanson, B.A.; Visvabharathy, L.; Ali, S.T.; Kang, A.K.; Patel, T.R.; Clark, J.R.; Lim, P.H.; Orban, Z.S.; Hwang, S.S.; Mattoon, D.; et al. Plasma Biomarkers of Neuropathogenesis in Hospitalized Patients with COVID-19 and Those with Postacute Sequelae of SARS-CoV-2 Infection. Neurol.-Neuroimmunol. Neuroinflamm. 2022, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Stasenko, S.; Hramov, A.; Kazantsev, V. Loss of neuron network coherence induced by virus-infected astrocytes: A model study. Sci. Rep. 2023, 13, 6401. [Google Scholar] [CrossRef] [PubMed]
- Sibille, J.; Zapata, J.; Teillon, J.; Rouach, N. Astroglial calcium signaling displays short-term plasticity and adjusts synaptic efficacy. Front. Cell. Neurosci. 2015, 189. [Google Scholar] [CrossRef]
- Haydon, P.G.; Nedergaard, M. How do astrocytes participate in neural plasticity? Cold Spring Harb. Perspect. Biol. 2015, 7, a020438. [Google Scholar] [CrossRef]
- Halassa, M.M.; Haydon, P.G. Integrated brain circuits: Astrocytic networks modulate neuronal activity and behavior. Annu. Rev. Physiol. 2010, 72, 335–355. [Google Scholar] [CrossRef]
- Jolivet, R.; Coggan, J.S.; Allaman, I.; Magistretti, P.J. Multi-timescale modeling of activity-dependent metabolic coupling in the neuron-glia-vasculature ensemble. PLoS Comput. Biol. 2015, 11, e1004036. [Google Scholar] [CrossRef]
- Linne, M.L.; Aćimović, J.; Saudargiene, A.; Manninen, T. Neuron–Glia Interactions and Brain Circuits. In Computational Modelling of the Brain: Modelling Approaches to Cells, Circuits and Networks; Springer International Publishing: Cham, Switzerland, 2022; pp. 87–103. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Glutamate-dependent astrocyte modulation of synaptic transmission between cultured hippocampal neurons. Eur. J. Neurosci. 1998, 10, 2129–2142. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef]
- Markram, H.; Tsodyks, M. Redistribution of synaptic efficacy between neocortical pyramidal neurons. Nature 1996, 382, 807–810. [Google Scholar] [CrossRef]
- Masquelier, T.; Deco, G. Network bursting dynamics in excitatory cortical neuron cultures results from the combination of different adaptive mechanism. PLoS ONE 2013, 8, e75824. [Google Scholar] [CrossRef]
- Barabash, N.; Levanova, T.; Stasenko, S. STSP model with neuron-glial interaction produced bursting activity. In Proceedings of the 2021 Third International Conference Neurotechnologies and Neurointerfaces (CNN), Kaliningrad, Russia, 13–15 September 2021; pp. 12–15. [Google Scholar] [CrossRef]
- Barabash, N.; Levanova, T.; Stasenko, S. Rhythmogenesis in the mean field model of the neuron–glial network. Eur. Phys. J. Spec. Top. 2023, 1–6. [Google Scholar] [CrossRef]
- Stasenko, S.V.; Lazarevich, I.A.; Kazantsev, V.B. Quasi-synchronous neuronal activity of the network induced by astrocytes. Procedia Comput. Sci. 2020, 169, 704–709. [Google Scholar] [CrossRef]
- Pankratova, E.V.; Kalyakulina, A.I.; Stasenko, S.V.; Gordleeva, S.Y.; Lazarevich, I.A.; Kazantsev, V.B. Neuronal synchronization enhanced by neuron–astrocyte interaction. Nonlinear Dyn. 2019, 97, 647–662. [Google Scholar] [CrossRef]
- Lazarevich, I.A.; Stasenko, S.V.; Kazantsev, V.B. Synaptic multistability and network synchronization induced by the neuron–glial interaction in the brain. JETP Lett. 2017, 105, 210–213. [Google Scholar] [CrossRef]
- Stasenko, S.V.; Kazantsev, V.B. Dynamic Image Representation in a Spiking Neural Network Supplied by Astrocytes. Mathematics 2023, 11, 561. [Google Scholar] [CrossRef]
- Gordleeva, S.Y.; Tsybina, Y.A.; Krivonosov, M.I.; Ivanchenko, M.V.; Zaikin, A.A.; Kazantsev, V.B.; Gorban, A.N. Modeling working memory in a spiking neuron network accompanied by astrocytes. Front. Cell. Neurosci. 2021, 15, 631485. [Google Scholar] [CrossRef]
- Tsybina, Y.; Kastalskiy, I.; Krivonosov, M.; Zaikin, A.; Kazantsev, V.; Gorban, A.N.; Gordleeva, S. Astrocytes mediate analogous memory in a multi-layer neuron–astrocyte network Neural. Comput. Appl. 2022, 34, 9147–9160. [Google Scholar] [CrossRef]
- Volman, V.; Bazhenov, M.; Sejnowski, T.J. Computational models of neuron-astrocyte interaction in epilepsy. Front. Comput. Neurosci. 2012, 6, 58. [Google Scholar] [CrossRef]
- Eid, T. Transforming Glia to Neurons Effectively Treats Temporal Lobe Seizures. Epilepsy Curr. 2022, 22, 130–131. [Google Scholar] [CrossRef]
- Mongillo, G.; Barak, O.; Tsodyks, M. Synaptic theory of working memory. Science 2008, 319, 1543–1546. [Google Scholar] [CrossRef]
- Wilson, H.R.; Cowan, J.D. Excitatory and inhibitory interactions in localized populations of model neurons. Biophys. J. 1972, 12, 1–24. [Google Scholar] [CrossRef]
- Tsodyks, M.; Pawelzik, K.; Markram, H. Neural networks with dynamic synapses. Neural Comput. 1998, 10, 821–835. [Google Scholar] [CrossRef] [PubMed]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef]
- Agulhon, C.; Petravicz, J.; McMullen, A.; Sweger, E.; Minton, S.; Taves, S.; Casper, K.; Fiacco, T.; McCarthy, K. What is the role of astrocyte calcium in neurophysiology? Neuron 2008, 59, 932–946. [Google Scholar] [CrossRef] [PubMed]
- Barres, B. The mystery and magic of glia: A perspective on their roles in health and disease. Neuron 2008, 60, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Benettin, G.; Galgani, L.; Giorgilli, A.; Strelcyn, J.M. Lyapunov characteristic exponents for smooth dynamical systems and for Hamiltonian systems; a method for computing all of them. Part 1: Theory. Meccanica 1980, 15, 9–20. [Google Scholar] [CrossRef]
- Borisov, A.V.; Jalnine, A.Y.; Kuznetsov, S.P.; Sataev, I.R.; Sedova, J.V. Dynamical phenomena occurring due to phase volume compression in nonholonomic model of the rattleback. Regul. Chaotic Dyn. 2012, 17, 512–532. [Google Scholar] [CrossRef]
- Borisov, A.V.; Kazakov, A.O.; Sataev, I.R. The reversal and chaotic attractor in the nonholonomic model of Chaplygin’s top. Regul. Chaotic Dyn. 2014, 19, 718–733. [Google Scholar] [CrossRef]
- Kazakov, A. On bifurcations of Lorenz attractors in the Lyubimov–Zaks model. Chaos Interdiscip. J. Nonlinear Sci. 2021, 31, 093118. [Google Scholar] [CrossRef] [PubMed]
- Karatetskaia, E.; Shykhmamedov, A.; Kazakov, A. Shilnikov attractors in three-dimensional orientation-reversing maps. Chaos Interdiscip. J. Nonlinear Sci. 2021, 31, 011102. [Google Scholar] [CrossRef] [PubMed]
- Levanova, T.A.; Kazakov, A.O.; Osipov, G.V.; Kurths, J. Dynamics of ensemble of inhibitory coupled Rulkov maps. Eur. Phys. J. Spec. Top. 2016, 225, 147–157. [Google Scholar] [CrossRef]
- Datseris, G.; Wagemakers, A. Effortless estimation of basins of attraction. Chaos Interdiscip. J. Nonlinear Sci. 2022, 32, 023104. [Google Scholar] [CrossRef] [PubMed]
- Datseris, G. DynamicalSystems. jl: A Julia software library for chaos and nonlinear dynamics. J. Open Source Softw. 2018, 3, 598. [Google Scholar] [CrossRef]
- Datseris, G.; Parlitz, U. Nonlinear Dynamics: A Concise Introduction Interlaced with Code; Springer Nature: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Pisarchik, A.N.; Hramov, A.E. Multistability in Physical and Living Systems; Springer: Berlin/Heidelberg, Germany, 2022; Volume 2, pp. 1–408. [Google Scholar] [CrossRef]
- Gordleeva, S.Y.; Stasenko, S.V.; Semyanov, A.V.; Dityatev, A.E.; Kazantsev, V.B. Bi-directional astrocytic regulation of neuronal activity within a network. Front. Comput. Neurosci. 2012, 6, 92. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olenin, S.M.; Levanova, T.A.; Stasenko, S.V. Dynamics in the Reduced Mean-Field Model of Neuron–Glial Interaction. Mathematics 2023, 11, 2143. https://doi.org/10.3390/math11092143
Olenin SM, Levanova TA, Stasenko SV. Dynamics in the Reduced Mean-Field Model of Neuron–Glial Interaction. Mathematics. 2023; 11(9):2143. https://doi.org/10.3390/math11092143
Chicago/Turabian StyleOlenin, Sergey M., Tatiana A. Levanova, and Sergey V. Stasenko. 2023. "Dynamics in the Reduced Mean-Field Model of Neuron–Glial Interaction" Mathematics 11, no. 9: 2143. https://doi.org/10.3390/math11092143