1. Introduction
The predator–prey interaction is one of the interactions that occurs in ecological problems. This interaction has been researched for decades by mathematicians who invest their time in creating accurate and useful models. The predator–prey model was first introduced independently by Alfred J. Lotka in 1925 and Vito Volterra in 1926 [
1]. The Lotka–Volterra predator–prey system for two species is modeled as follows:
In the Lotka–Volterra predator–prey model, the variables and represent the prey and predator population densities, respectively. The intrinsic growth rate of the prey is represented by . The predation rate of the predator on the prey is represented by . The growth of the predator is assumed to be influenced only by the predation effect, with a growth rate represented by . Finally, the parameter is the natural mortality rate of the predator.
To incorporate the human effect into a natural growth of a population, researchers then introduced the harvesting effect into the growth model in various ways. As an example, in 1957, Schaefer [
2] introduced a harvesting model in a simple fisheries ecosystem without considering predation. Harvesting is expressed as a function
, where
and
represent catchability coefficient and harvesting effort, respectively. The Schaefer [
2] model was developed as a management tool for the Eastern Tropical Pacific Tuna Fishery [
3]. The Schaefer [
2] fishery model is presented as follows:
The Schaefer model [
2] consists of only one equation with only one variable. The variable can be represented as either predator or prey. The parameter
represents the tuna population’s carrying capacity. Besides the harvesting function in Schaefer [
2], there are also other harvesting functions, including the constant harvesting function [
4,
5], the rational harvesting function [
6,
7,
8], the periodic harvesting function [
9,
10,
11], and the piecewise harvesting function [
11,
12].
Since real problems are more complex than those modeled by the Lotka–Volterra and Schaefer models, the two models have been widely combined and developed to get realistic modeling or at least one that is close to the real problem. Meng et al. [
13] combined the two models by considering a disease in a species so that the prey species consisted of susceptible and infected preys. Thirthar et al. [
14] studied three species of prey, predator, and superpredator species and added the effect of fear between predators and prey. They showed in their analysis that fear has a noticeable effect on the system’s stability. Suryanto et al. [
15] studied the fractional derivative Caputo model with a ratio-dependent predation function. The fractional model was formed by them because the population growth rate depends on long-term memory. Panigoro et al. [
16] studied the fractional derivative Caputo predator–prey model and harvesting by considering the age structure of predators consisting of juvenile and adult predators. They assumed that only adult predators can prey on preys.
In recent years, the development of research on predator–prey mathematical models and harvesting by mathematicians has extended to marine reserves (can be seen in [
17]). Mapunda et al. [
18] studied the predator–prey system by considering the creation of a marine reserve as a solution to over-exploitation and drought in the system. Based on their results, when the creation of marine reserves is not performed, it will have a negative impact on the predator and prey populations. Abid et al. [
19] analyzed the stability and determined the optimal harvesting of the modified Leslie–Gower predator–prey model by considering the implicit marine reserve. Ibrahim [
20] proposed a predator–prey model that considers the implicit marine reserve and critical biomass level with the harvesting function following the harvesting function in [
2,
21], namely
where
,
, and
are the catchability coefficient, constant harvesting effort, and prey population density. Furthermore, the research model from Ibrahim [
20] is presented as follows:
Based on the model in (2), Ibrahim [
20] assumed that changes in population density increase with prey population growth and decrease with prey harvesting. Prey population growth is described by a logistic function,
, with
and
being the prey’s carrying capacity and intrinsic growth rate, respectively. Then, prey harvesting is described by the function in (1), with
being the implicit marine reserve fraction, i.e., the fraction of prey stock allowed to be harvested. Therefore,
is the fraction of prey stock that is not allowed to be harvested. Furthermore, the change in fishing effort can increase or decrease depending on the value of
, where
is the economically critical stock size level. The term
has the interpretation that when the prey population density
is greater than the threshold
(
), the effort (
) in harvesting the prey population increases. When the prey population density is smaller than threshold
, harvesting effort (
) decreases. In this condition, if
is still increased, the fishermen will not get profit (loss). Finally, when
, the effort in harvesting does not change (constant).
In this study, we modify the model in [
20] by replacing the term
with
. The modified model is presented as follows:
Based on the economic system prevailing in most regions, fishermen will flock there when the fishery shows success. As a result, the fishing rate increases and the fish population catches up [
21]. Therefore, the amount of fishing effort made by fishermen also greatly affects the rise and fall of the fishing rate. Based on the term
, the harvesting effort on prey increases when
, which is the ratio of prey population density to fishing effort greater than
. Then, when
, the harvesting effort on the prey decreases. This means that some fishermen choose to move to other waters because fishermen can experience losses if they continue harvesting in these waters. Finally, when
, the harvesting effort does not change.
This study aims to explore the solution properties of the model developed in (3). The exploration of the solution properties of the model in (3) is presented in
Section 2. Then, this study determines the equilibrium points of the model in (3) and analyzes their local stability, which is presented in
Section 3 and
Section 4, respectively. In
Section 5 and
Section 6, the asymptotic global stability and existence of limit cycles are analyzed. The bionomic equilibrium point is determined and discussed in
Section 7. In
Section 8, numerical simulations are performed to demonstrate the results obtained graphically. Next,
Section 9 aims to discuss or show that there are overlapping conditions between the model we developed and the model developed by Ibrahim [
20]. These conditions are obtained by selecting certain parameters. Finally, the conclusion of this study is presented in
Section 10.
4. Local Stability of Each Equilibrium Point
The local stability of each equilibrium point for system (3) is explored with the help of its Jacobian matrix. If the real parts of all eigenvalues of the Jacobian matrix for each equilibrium point are nonpositive, then system (3) is locally stable toward the corresponding equilibrium point. Conversely, if the real part of any eigenvalue is positive, system (3) is not locally stable at the corresponding equilibrium point. Suppose the eigenvalues of the Jacobian matrix of an equilibrium point cannot be obtained explicitly. In that case, the help of the Routh–Hurwitz criterion is needed in determining the local stability of the equilibrium point. The local stability of the equilibrium points , , and are presented in Theorem 3, Theorem 4, and Theorem 5, respectively.
Theorem 3. For the model in (3), the equilibrium point is always a saddle.
Proof of Theorem 3. For equilibrium point
, the Jacobian matrix is
Furthermore, the eigenvalues obtained from the matrix are and . Since , the equilibrium point is always a saddle. □
Theorem 4. For the model in (3), the equilibrium point is always a saddle.
Proof of Theorem 4. For equilibrium point
, the Jacobian matrix is
Furthermore, the eigenvalues obtained from the matrix are and . Based on these two eigenvalues, the equilibrium point is a saddle. □
Theorem 5. For the model in (3), the equilibrium point is always asymptotically locally stable. Furthermore, the equilibrium point yields the following
- 1.
Semi-spiral sink or star sink, if ;
- 2.
Sink, if ;
- 3.
Spiral sink, if .
Proof of Theorem 5. Evaluation of the Jacobian matrix at equilibrium point
, namely
Therefore, the equilibrium point
is locally asymptotically stable. Furthermore, the value of
Equation (8) is zero if . Therefore, if Equation (8) is zero, the equilibrium point produces a semi-spiral sink or star sink. Then, Equation (8) is less than zero if . Therefore, if Equation (8) is less than zero, the equilibrium point generates a sink. Finally, Equation (8) is greater than zero if . Therefore, if Equation (6) is greater than zero, the equilibrium point produces a spiral sink. □
8. Numerical Simulations
This section presents numerical simulations of the results obtained in the previous sections. The software used for this numerical simulation was Maple 2019. All the codes are attached to the link listed in the
Supplementary Materials section. Based on the results in
Section 3, there are three equilibrium points, including (1) equilibrium point
(trivial), which is the point where the element value of the equilibrium point is zero, meaning that the population density of prey species
is extinct and there is no capture effort
; (2) equilibrium point
, which is the point where the population density of the prey
is equal to
while there is no capture effort
; (3) equilibrium point
, which is the point where the prey density
and capture effort
) exist. One of the three equilibrium points is always stable, namely equilibrium point
. Meanwhile, equilibrium points
and
are always saddle. Furthermore, equilibrium point
is globally asymptotically stable, and system (3) has no limit cycle around equilibrium point
.
Numerical simulations were explored to show the local stability results of the three equilibrium points in Theorems 3, 4, and 5. The values of each parameter were , , , and . Meanwhile, the value of was varied based on the three conditions that satisfy Theorem 5. The value of is obtained from the result of , which is . To fulfill the first, second, and third conditions of Theorem 6, parameter was greater than, equal to, and smaller than with values of ,, and 0.025.
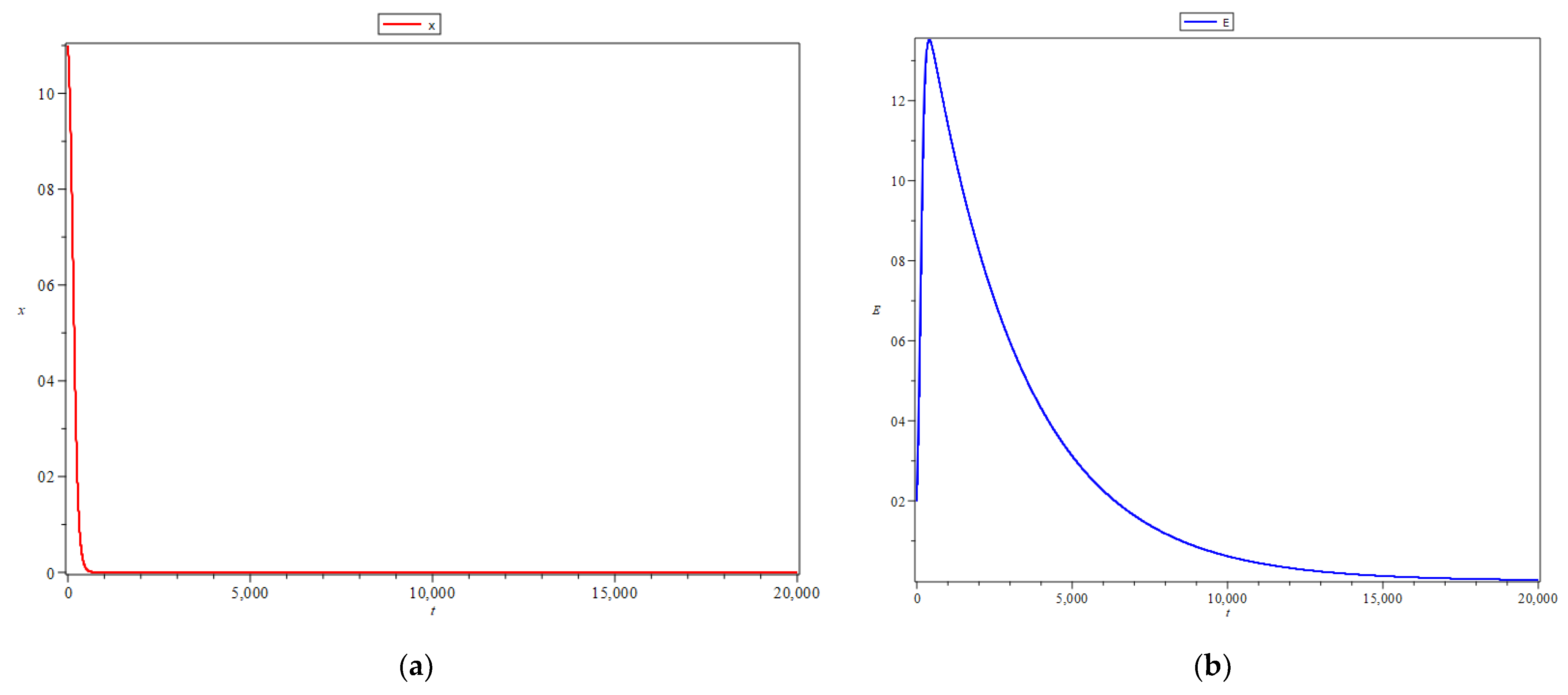
The numerical simulation result for
is presented in
Figure 1. This choice of parameter
is intended to show that equilibrium point
produces a semi-spiral sink or star sink.
Figure 1 shows that all initial values of
and
in the positive quadrant will lead to equilibrium point
. In
Figure 1, the blue line represents more clearly that for some initial points
, including
For , the points tend to equilibrium point
The numerical simulation result for
is presented in
Figure 2. The choice of parameter
is intended to show that equilibrium point
generates a sink.
Figure 2 shows that all initial values of
and
in the positive quadrant will lead to equilibrium point
. In
Figure 2, the blue line represents more clearly that for some initial points
, including
For , the points tend to equilibrium point
The numerical simulation result for
is presented in
Figure 3. The choice of parameter
is intended to show that equilibrium point
produces a spiral sink.
Figure 3 shows that all initial values of
and
in the positive quadrant will lead to equilibrium point
. In
Figure 3, the blue line represents more clearly that for some initial points
, including
For , the points tend to equilibrium point
Furthermore,
Figure 1,
Figure 2 and
Figure 3 show that all initial values of
and
around equilibrium points
,
, and
converge to the corresponding equilibrium point
. It shows that the system around the equilibrium points
and
is always a saddle. Consequently, the equilibrium points are globally stable since they are valid for initial values in the positive quadrant. In addition,
Figure 1,
Figure 2 and
Figure 3 also show that the equilibrium points are asymptotically globally stable, as shown by the absence of limit cycles.
9. Discussion
A predator–prey model considering implicit marine protected areas and critical biomass level was built and analyzed in this study. The model is presented in (3), which is a modification of the model from Ibrahim [
20] in (2). Ibrahim [
20], in his research, examined the boundedness of the solution of the model in (2). We also examined the boundedness of the solution of the model in (3). Furthermore, we examined the existence, uniqueness, and permanence of the solution of the model in (3), which was not studied by Ibrahim [
20].
The next discussion in this paper is to determine the local and global stability of the equilibrium points of model (3). The determination of stability begins with the determination of the equilibrium point of model (3). Based on the results obtained in
Section 3, there are three equilibrium points, namely
,
, and
. We found that the equilibrium points
and
of the models in (2) and (3) are the same:
and
, respectively. However, the equilibrium point
of the models in (2) and (3) can have the same and different values. The equilibrium point
of model (2) is
. Then, the equilibrium point
of model (3) is
. Furthermore, the equilibrium point
of the model in (3) always exists. Meanwhile, the equilibrium point
of the model in (2) will exist if
, which is stated in Ibrahim’s research [
20]. The following remark presents the conditions under which the equilibrium points
of models (2) and (3) are equal at
.
Remark 1. The equilibrium point of models (2) and (3) is the same if and only if .
Referring to Ibrahim’s research [
20], the parameter values
, and
Based on Remark 1, we obtain
. Therefore, the equilibrium point
is 300 tons of fish and one fishing trip. With these parameters, the numerical simulation results for models (2) and (3) are presented in
Figure 4 and
Figure 5, respectively. The steady state of model (3) is faster than the steady state of model (2) at
with initial values of
and
. In addition, we tried to change the value of
to a smaller value,
. As a result, we obtained a value of
. The simulation results for the new values of
and
are presented in
Figure 6 and
Figure 7.
Figure 6 and
Figure 7 show that the results for parameter
are opposite to the results for parameter
.
Figure 6 and
Figure 7 show that the steady state of model (2) is faster than the steady state of model (3) at
with initial values
and
.
After determining the equilibrium points of model (3), the next step was to determine the local and global stability of each of the equilibrium points. Based on our analysis, the local stability of and is always saddle, and is always asymptotically stable. This result is similar to the equilibrium points and of model (2), which are also always saddle, and is always asymptotically stable. In addition, equilibrium point is always globally stable, as proved using Lyapunov’s theorem. Furthermore, equilibrium point is globally asymptotically stable, and system (3) has no limit cycle around equilibrium point . The results obtained are also similar to those of the equilibrium point of model (2) in that the equilibrium point is asymptotically globally stable.
Figure 4,
Figure 5,
Figure 6 and
Figure 7 show that the same value of parameters for models (2) and (3) and
will result in the system stabilizing toward the same equilibrium point, which is at
. The following theorem shows that different parameter values of models (2) and (3) can also result in a stable system at any equilibrium point
.
Theorem 8. For any parameters , , , , and of models (2) and (3) with equilibrium points of models (2) and (3), respectively, i.e., and . Let , , , , and be the parameters , , , , and of model (i) for . The two equilibrium points have the same values, namely if and only if and with and .
Proof of Theorem 8. Suppose the two equilibrium points,
and
are equal. This means,
Based on the first equation in (11), we have
By elaborating the second equation of (11), we obtain
Based on (12), then (13) becomes
So, if , then and .
Then,
and
. We have
with
Hence, if (15) is satisfied, then the equilibrium points and will have the same value. □
Remark 2. For the parameter values , , and and let and be the parameters and of the model (i) for , the equilibrium point of model (2) and that of model (3) are equal if and only if and .
For model (2), Ibrahim [
20], in his research, assumed that
, i.e., 60% of the fish population in the sea is allowed to be harvested. The parameters
,
, and
As a result, the equilibrium point
. That is, there are 300,000 tons of fish and 920,370 fishing trips in a steady state. In this case, we assume that
,
and
values are the same as in Ibrahim’s study [
20]. Based on Remark 2,
and
. From the perspective of the model (3), equilibrium condition
can be achieved by applying the same marine reserve where 60% of the fish population is allowed to be harvested. Meanwhile, the threshold value of
, the ratio of the total fish population in the sea to the number of fishing trips, is
.
Figure 8 shows that the trajectories of model (2) and model (3) with these parameters spiral inward toward the same equilibrium point with 300,000 tons of fish and 920,370 fishing trips. This number of fishing trips is twice the number of fishing trips (effort) at maximum sustainable yield (MSY), which is 394,444 [
20]. In
Figure 8, the cyan arrow and red solid line represent the model in (2). Meanwhile, the gray arrow and blue solid line represent the model in (3).
10. Conclusions
This study constructs a predator–prey model considering marine protected areas and critical biomass levels. In this case, the critical biomass level is assumed to be proportional to the fishing effort . The model is presented in (3). The stability properties of the model are analyzed by determining the equilibrium points and the local stability by Jacobian linearization and global stability by the Lyapunov theorem of each equilibrium point.
Based on the stability analysis, there are three equilibrium points, , , and . Then, one of the equilibrium points, , is always stable. Meanwhile, equilibrium points and are always saddle. Furthermore, the global stability analysis of equilibrium point is analyzed by the Lyapunov theorem. As a result, equilibrium point is globally asymptotically stable. The presence or absence of a limit cycle is shown using the Bendixson–Dulac criterion. We found that system (3) has no limit cycles around equilibrium point .
In
Section 9, we discuss the model studied by Ibrahim [
20], namely the model in (2), alongside our model, namely the model in (3). We conclude that the equilibrium points
and
of models (2) and (3) have the same values and the same type of stability. Furthermore, the equilibrium point
of model (2) and that of model (3) also have the same type of stability. Then, we derive one theorem and two remarks regarding determining parameter values of models (2) and (3) that result in the same value of equilibrium point
. Finally, when 60% (
) of the marine fish population is allowed to be harvested and
, the number of fishing trips (effort) is twice that of fishing trips at maximum sustainable yield (MSY).
So far, we have only focused on showing that models (2) and (3) have overlapping conditions, especially the similarity of the value of equilibrium point that occurs under certain conditions. Furthermore, studies on equilibrium points and are also interesting to explore. For example, if possible, it would be better to show the regions of attraction and repulsion for and in the region and . In addition, it would be interesting to investigate further the possibility that model (3) provides some types of dynamics that cannot be obtained in model (2). A comparison of the application of models (2) and (3) to natural observational data would also be interesting. This comparison would show which model is better for real-world problems.
The results above are deduced from the specific assumptions used in the construction of the model. It would be interesting to explore the robustness of the results for other ecological issues that are worthy to be taken into account in developing a more realistic model, such as different levels of intraspecific competition [
28], the Allee effect [
29,
30], fear effect [
31,
32], and a coupled growth function caused by a metapopulation structure [
33], which may alter the result in this study. These are being investigated by the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







