1. Introduction
The world has long been confronted with hazads of climate change caused by global warming. China is taking pragmatic actions facing the challenges brought by climate change. At the general debate of the 75th Session of the United Nations General Assembly on 22 September 2020, President Xi Jinping announced that China would scale up its NDCs (Nationally Determined Contributions) by adopting more vigorous policies and measures, strive to peak emissions before 2030, and achieve carbon neutrality before 2060.
Forests’ annual carbon sequestration accounts for about 2/3rd of the whole terrestrial system, which is the main body of the terrestrial ecosystem. According to the China Forest Resources Report, by 2018, China’s forest coverage rate was 22.96%, forest area was 220 million , forest stock volume was 17.56 billion , and total carbon storage was 91.86 billion tons. From 1990 to 2020, China’s forest carbon sink capacity witnessed an escalation from 185.5 to 321.4 . A vigorous increase of forest carbon sink has become a top priority to achieve the carbon peaking and carbon neutrality goals. Effective control of forest pests and diseases can prolong the growth cycle of trees and increase the forest carbon sink, which is of great significance for the realization of China’s “30·60” goal.
Pine wilt disease (PWD) is a devastating forest disease caused by pine wood nematode (PWN). The PWD is a multipartite system involving intimate relationships between the pathogen, PWN, Bursaphelenchus xylophilus (Steiner & Buhrer) Nickle, its vectors, and symbiotic microorganisms [
1]. It is mainly transmitted by
Monochamus alternatus (
M. alternatus) in Asia [
1], which spreads rapidly and kills trees quickly. Through consulting relevant data, PWN is thought to have originated in North America [
2], then gradually invaded other countries, such as Japan [
3], Korea [
4], China [
5], Mexico [
6], and Spain [
7]. How to effectively control the occurrence and spread of PWD has become the focus and frontier topic of researchers. Ecologists have made remarkable progress in many aspects. Kim et al. [
8] used recombinant BxPrx as an antigen to generate a novel antibody that can be used to quickly and accurately determine PWD. Ding et al. [
9] improved the genome sequence of PWN and explained the interaction between PWD and pine trees. Palomares-Rius et al. [
10] determined a gene set affected by genomic variation finding that the level of genomic diversity of PWN was related to its phenotypic variability, including variations in pathogenicity and ecological traits. Presently, global strategies for PWD prevention and control encompass chemical control, physical control, biological control and biomimetic technology, with avermectin (AVM) as a predominant insecticidal agent [
11]. Lee et al. [
12] conducted comparative analyses on 16 avermectin benzoate formulations against PWD to support disease control. Alvarez et al. [
13] engineered diverse trap designs assessing their efficacy in maximizing the attraction and retention of live insects through field experiments and comparative modeling. Mannaa et al. [
14] found that treatment with resistance-induced chemical inducers MeSA and ASM significantly reduced the severity of PWD, providing new ideas for its prevention and treatment.
While ecologists have largely concentrated on the biological structure, distribution and control factors of PWN, attention to the dynamic characteristics of PWD transmission system remains scant. In recent years, mathematicians have established various models to predict the occurrence trend of PWD. Shi and Song [
15] investigated the dynamical behavior of PWD by incorporating a standard incidence rate and the threshold value of the relative basic reproductive number
which determined the spread of infection has been worked out. Ozair [
16] discussed the global stability of PWD by considering the nonlinear incidence rate with the horizontal transmission in the model. Khan et al. [
17] introduced a mathematical model that described the dynamics of PWD by presenting the stability analysis of the disease-free and endemic equilibria base on basic reproduction number
, and an optimal control strategy was formulated by adding control variables related to time to the model. Subsequent work by Khan et al. [
18] continued this line of inquiry by exploring the effect of asymptomatic carriers of PWD and further elaborating on the optimal control strategies in 2020.
As we know from the literature, most of the literature studied the occurrence of PWD in its natural state or the effects of prevention and control on PWD infectivity, with limited examination of time-delay in the control process. Therefore, it is feasible to propose a model that can comprehensively show intensity and time-delay of disease control. Based on the infectious disease model, we divide
M. alternatus into susceptible
M. alternatus (not carrying PWN) and infected
M. alternatus (carrying PWN). After the outbreak of PWD in a forest area, we usually take measures to protect pine trees and kill
M. alternatus. However, given the extensive adaptability of PWN and the rapid spread of PWD, there is a certain time delay of the control to take effect (that is, the infection rate of
M. alternatus begins to decline). We use delay differential equations to describe the dynamic changes in the insect-vector populations system more truly and accurately. Delay differential equations are used to describe the development systems that depend on both the current state and the past state and have been widely used in many fields. In the study of the bifurcation phenomenon, it is very important to derive the bifurcation normal form of differential equations. Nayfeh [
19] proposed the method of multiple time scales (MTS) to solve the problem of nonlinear vibration and gave the calculation process of Hopf bifurcation normal form of delay differential equations by MTS in 2008 [
20]. Later, many scholars studied the stability and bifurcation theory of various differential equations [
21,
22,
23]. Based on this background, we establish a two-dimensional differential equation model with time delay to discuss the stability and bifurcation phenomenon of PWD infection-control system to predict the occurrence of PWD, and provide theoretical support for the prevention and control of PWD.
The rest of the content is arranged as follows. In
Section 2, we first build a differential equation model with time delay based on the epidemic model among the medium insects. In
Section 3, we analyze the existence and stability of equilibrium and the existence of Hopf bifurcation for the model with time delay. In
Section 4, we derive the normal form of the Hopf bifurcation by using MTS and analyze the stability of the periodic solution of the Hopf bifurcation. In
Section 5, we discuss and analyze the unknown parameters in the model and then present numerical simulations to verify the correctness of the theoretical analysis. Finally, the conclusion is drawn in
Section 6.
2. Mathematical Modeling
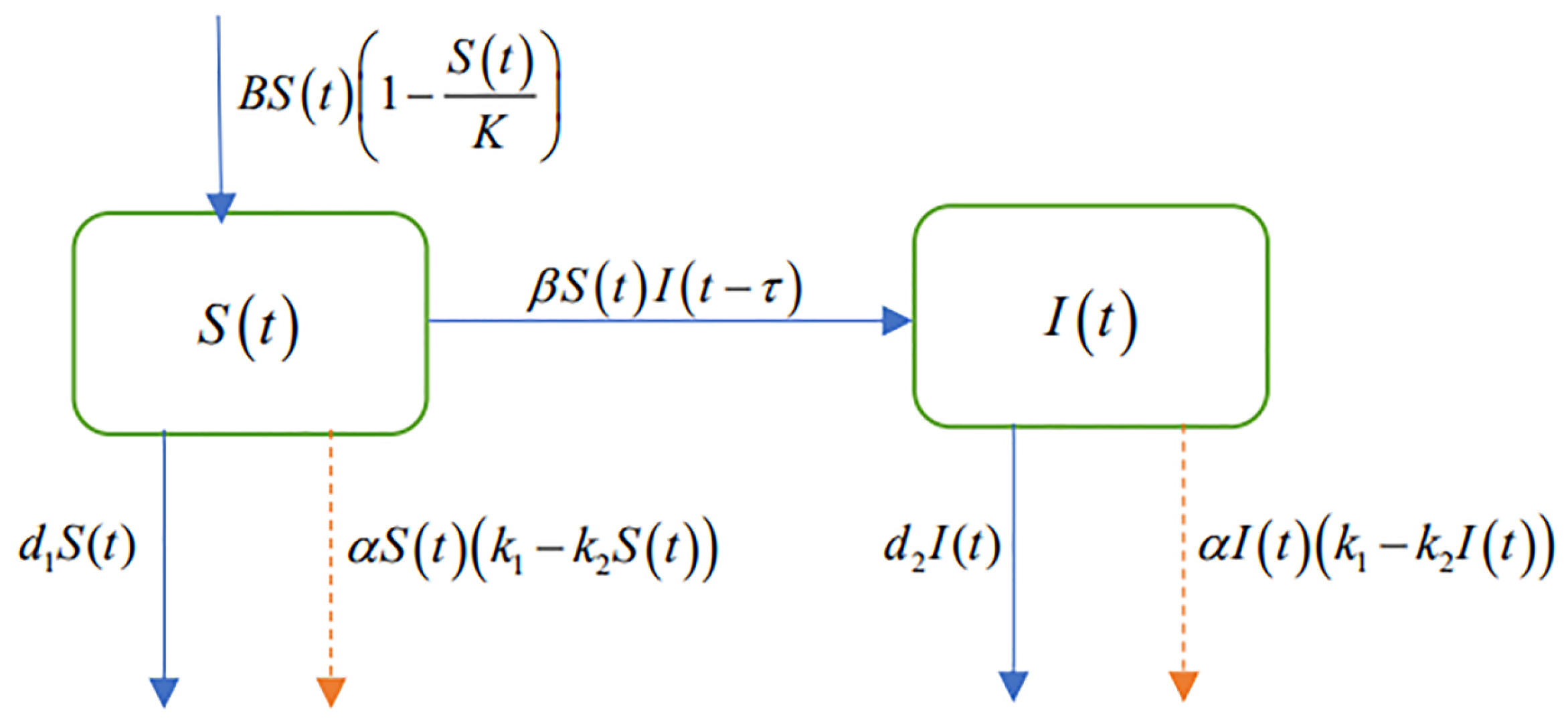
After the outbreak of PWD in an area, we take control measures (such as nematicide injection and vaccination) to protect pine trees. The transmission of PWN is not reliant upon direct contact between M. alternatus, but hinges on the process of infected M. alternatus transmitting PWN to healthy pine trees during feeding and oviposition. Newly formed adult M. alternatus remain within the pupation chamber prior to emergence, where they become infected by PWN from deceased host tree wood, then they carry PWN to continue to infect other healthy pine trees after emergence. Upon implementation of nematicide injection or vaccination, the quantity of PWN in pine trees diminishes, leading to a concomitant decline in infection rate. Therefore, taking control measures on trees can effectively reduce the infection rate of M. alternatus. Moreover, measures such as insecticidal spraying insecticides can kill M. alternatus, which also reduces the rate of infection. However, the transmission speed of PWN is very fast when the forest is in the outbreak period of PWD, so in the early stage of control, the transmission efficiency of PWN may be higher than that of control. Consequently, there is a time delay between taking control measures and the beginning of the decline in the infection rate of M. alternatus, so there is a certain time delay in the effective of prevention and control. Since not every time infected M. alternatus can “feed” PWN, we introduce an infection coefficient. We suppose that the infection coefficient of PWD is reduced to after adopting control measures. Other influencing factors in the infection-control system are analyzed below.
It is assumed that
M. alternatus are divided into susceptible
M. alternatus which did not carry PWN and infected
M. alternatus which did carry PWN at a certain time in a PWD epidemic area. For the input of
M. alternatus, the born
M. alternatus and the dead
M. alternatus are mainly considered. Because the resources are limited, it is more realistic for us to use the logistic function to describe the growth rate of the
M. alternatus. We assume the natural mortality of susceptible and infected
M. alternatus as
,
, respectively. In the process of the spread of PWD, prevention and control can increase the mortality rate of
M. alternatus, thereby inhibiting the spread of the disease. With the progress of control, the mortality rate of
M. alternatus will tend to be saturated. Therefore, the number of
M. alternatus killed by artificial control grows following nonlinear logistic growth, and the mortality rate of
M. alternatus is related to the intensity of control, with the increase of control efforts, the mortality rate also increases nonlinearly, so we use
to describe its linear part and
to describe its nonlinear part, where
represents control measures efficiency of PWD. Adding the nonlinear part better reflects the saturation effect of artificial prevention and control, which is more consistent with reality. To better study the impact of prevention and control on PWD infection-control system, we present the variable relationships shown in
Figure 1.
According to
Figure 1, we can construct the following delayed differential equation model:
where
and
are the variables;
B,
,
,
,
,
,
and
K are the positive constants; and
is the time delay of disease control to take effect. The specific descriptions are given in
Table 1.
For convenience, we denote that
,
,
; then, model (
1) becomes:
Due to the wide distribution and strong concealment of M. alternatus, we believe that the number of births of susceptible M. alternatus is always greater than the number of deaths, that is, , which is also consistent with the data we found in the later parameter analysis. Moreover, we believe that the intensity of prevention and control against M. alternatus will be change within 0∼1, that is, the maximum value of is 100%.
Then, we prove that the solution of system (
2) is nonnegative under positive initial conditions.
The initial condition of system (
2) is
, where
,
,
is a continuous function mapping from
to
in Banach space, for system (
2),
.
Theorem 1. If , , , then the solution of system (2) , is nonnegative for . Proof. Assume that the system (
2) in the nonnegative initial function
,
, the solution
is not nonnegative when
, then there must be the first time
, such that
,
. According to the first equation of system (
2), we can obtain
, contradicting with
at this time.
Similarly, assuming that the solution of system (
2)
is not nonnegative when
in the case of nonnegative initial function
,
, then there must be a first time
, such that
,
,
, according to the second equation of system (
2):
, note the parameter
, so
, contradicting with
at this time.
In summary, when
, the solutions
and
of system (
2) are still nonnegative for nonnegative initial functions. □
4. Normal Form of Hopf Bifurcation
In
Section 3, we have shown that when
, the equilibrium
is locally asymptotically stable for any
; when
, the bifurcating periodic solution near the equilibrium
is unstable by Theorem 2. Thus, we only care about the stability of bifurcating periodic solution near the positive equilibrium
. In order to be more realistic, we focus on the delay between taking control measures and the beginning of control to take effect. Therefore, we consider the time-delay
as a bifurcation parameter and denote the critical value
, where
is given in Equation (
14). When
, Equation (
13) has a pair of pure imaginary roots
. Therefore, system (
2) undergoes a Hopf bifurcation near equilibrium
. In this section, we derive the normal form of Hopf bifurcation for the system (
2) by using the multiple time scales method.
In order to normalize the delay, we first re-scale the time
t by using
, then translate the equilibrium
to the origin, so system (
2) is transformed into:
Equation (
15) can also be written as:
where
and
Let
h be eigenvector corresponding to eigenvalue
of linearized system of Equation (
16), and
be the eigenvector corresponding to eigenvalue
of adjoint matrix of linearized system of Equation (
16), satisfying:
By calculating, we have:
where
.
We suppose the solution of Equation (
16) is as follows:
where
The derivative with respect to
t is transformed:
where
is differential operator, and:
Using a Taylor series expansion of
, we obtain: that
where
.
As we stated,
is the bifurcation parameter, and
, where
is the Hopf bifurcation critical value,
is perturbation parameter, and
is dimensionless scale parameter. Substituting Equations (
19)–(
22) into Equation (
16) and balancing the coefficients before
on both sides of the equation, the following expression is obtained:
Thus, Equation (
23) has the following solution form:
The expression of the coefficient before
is as follows:
Substituting Equation (
24) into the right-hand side of Equation (
25), and the coefficient vector of
is denoted by
. According to the solvability condition
, the expression of
is obtained as follows:
where
.
Since
is a disturbance parameter, we only consider its effect on the linear part. Therefore, we ignore the part containing
in the higher order. We suppose the solutions of Equation (
25) are given as follows:
where
where
are given in Equation (
18) and
The expression of the coefficient before
is:
Next, substituting solution (
24) and (
27) into Equation (
29), and with the coefficient vector of
noted as
, by solvability condition, we have
. Note that
is a disturbance parameter, and
has little influence for small unfolding parameter, and thus, we can ignore the
, then the expression of
can be obtained as follows:
where
where
are given in Equation (
18), and
are given in Equation (
28).
Let
, then, the deduced third-order normal form of Hopf bifurcation of system (
2) is:
where
M is given in (
26) and
H is given in (
30).
Substituting
into Equation (
31), the following normal form of Hopf bifurcation in polar coordinates is obtained:
According to the normal form of Hopf bifurcation in polar coordinates, we only need to consider the first equation in system (
32). Thus, the following theorem holds:
Theorem 4. For the system (32), when , there is a semitrivial fixed point , and system (2) has periodic solution. - (1)
If , then the periodic solution reduced on the center manifold is unstable.
- (2)
If , then the periodic solution reduced on the center manifold is stable.
6. Conclusions
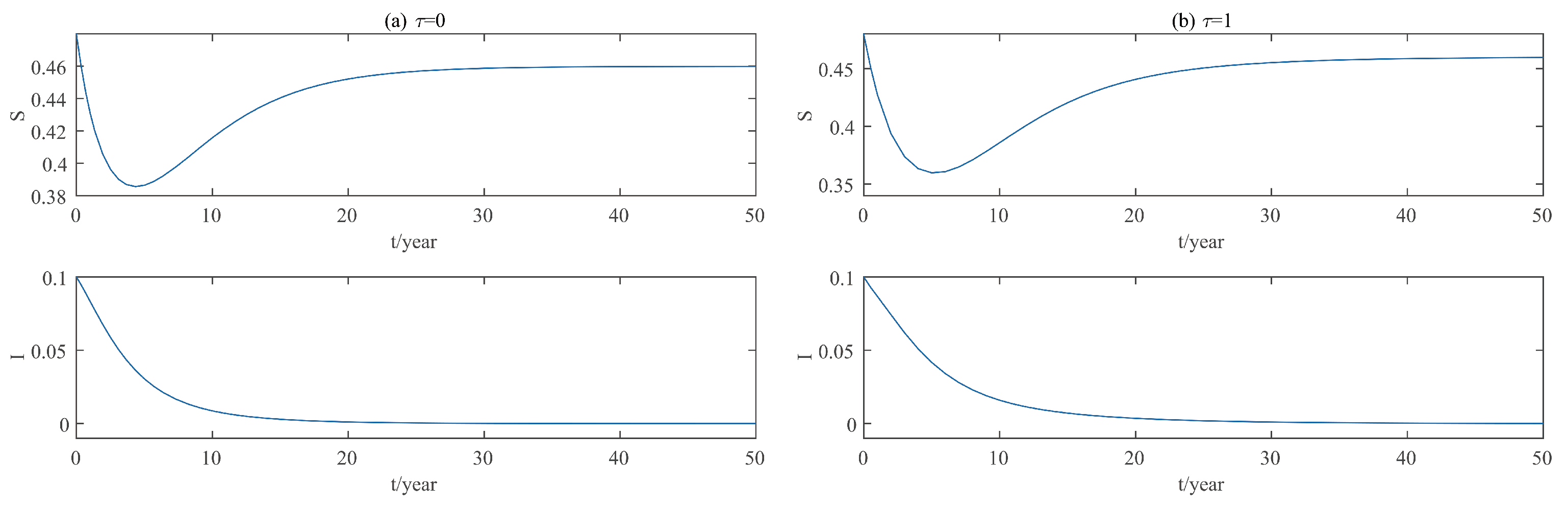
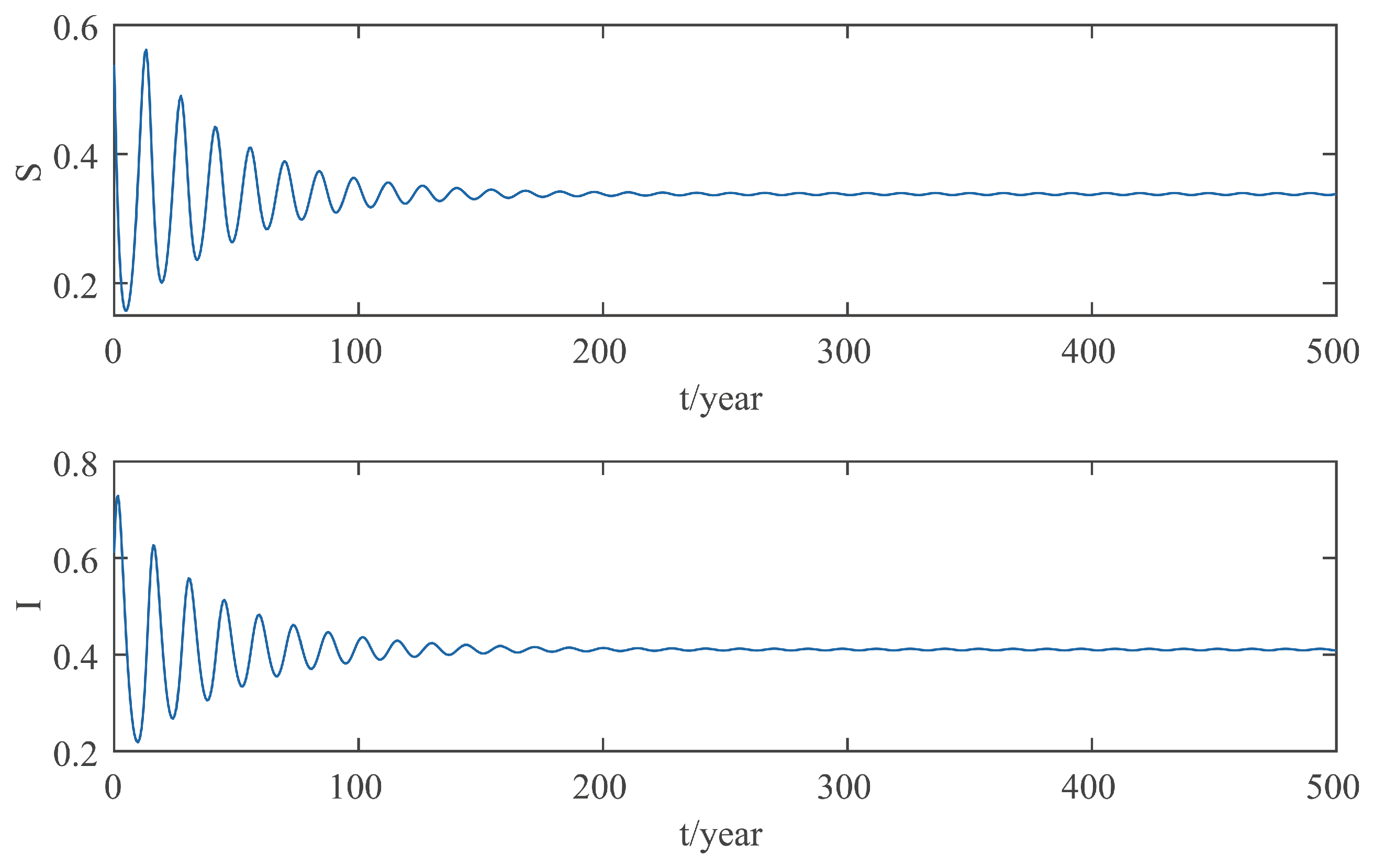
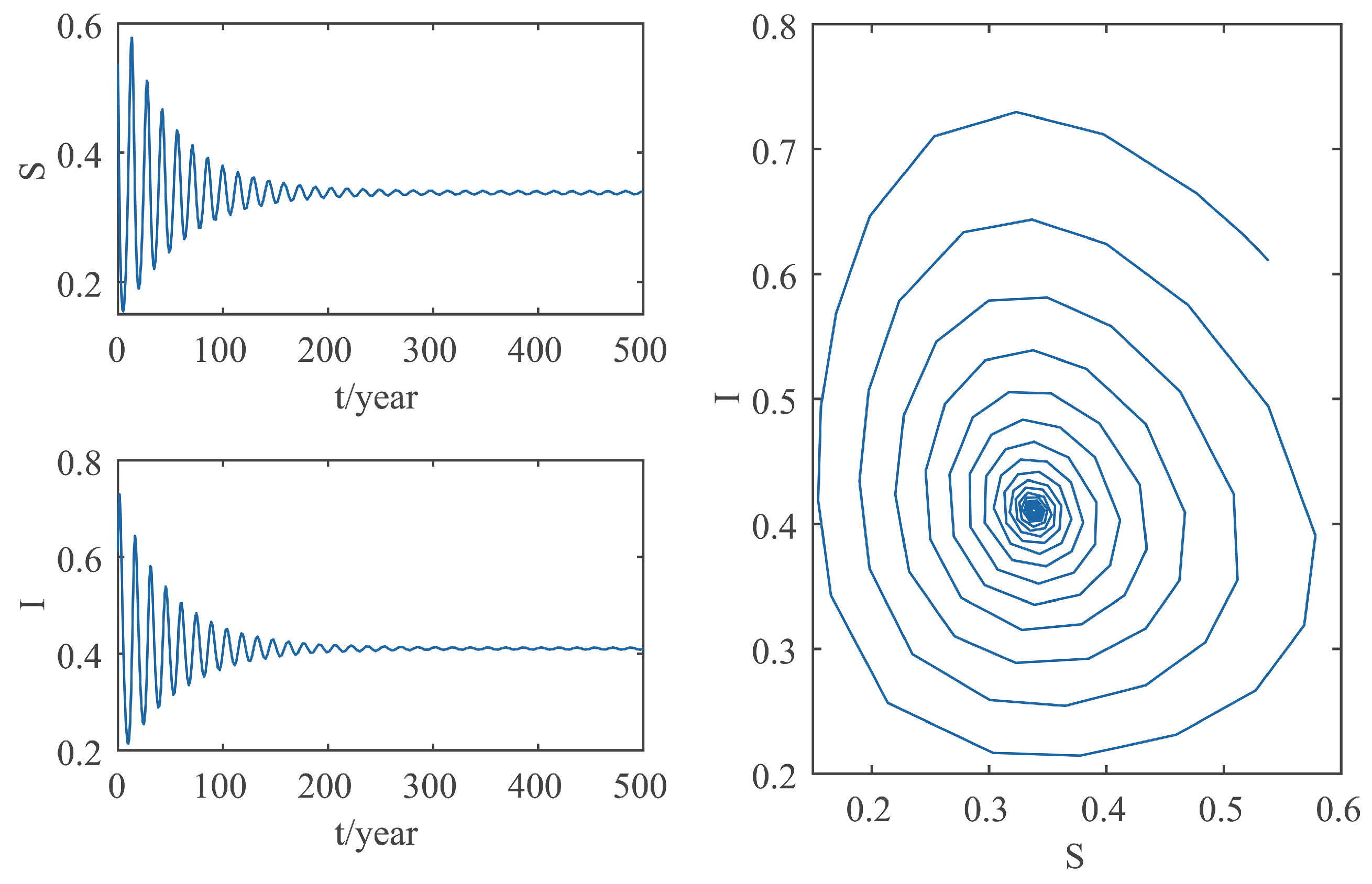
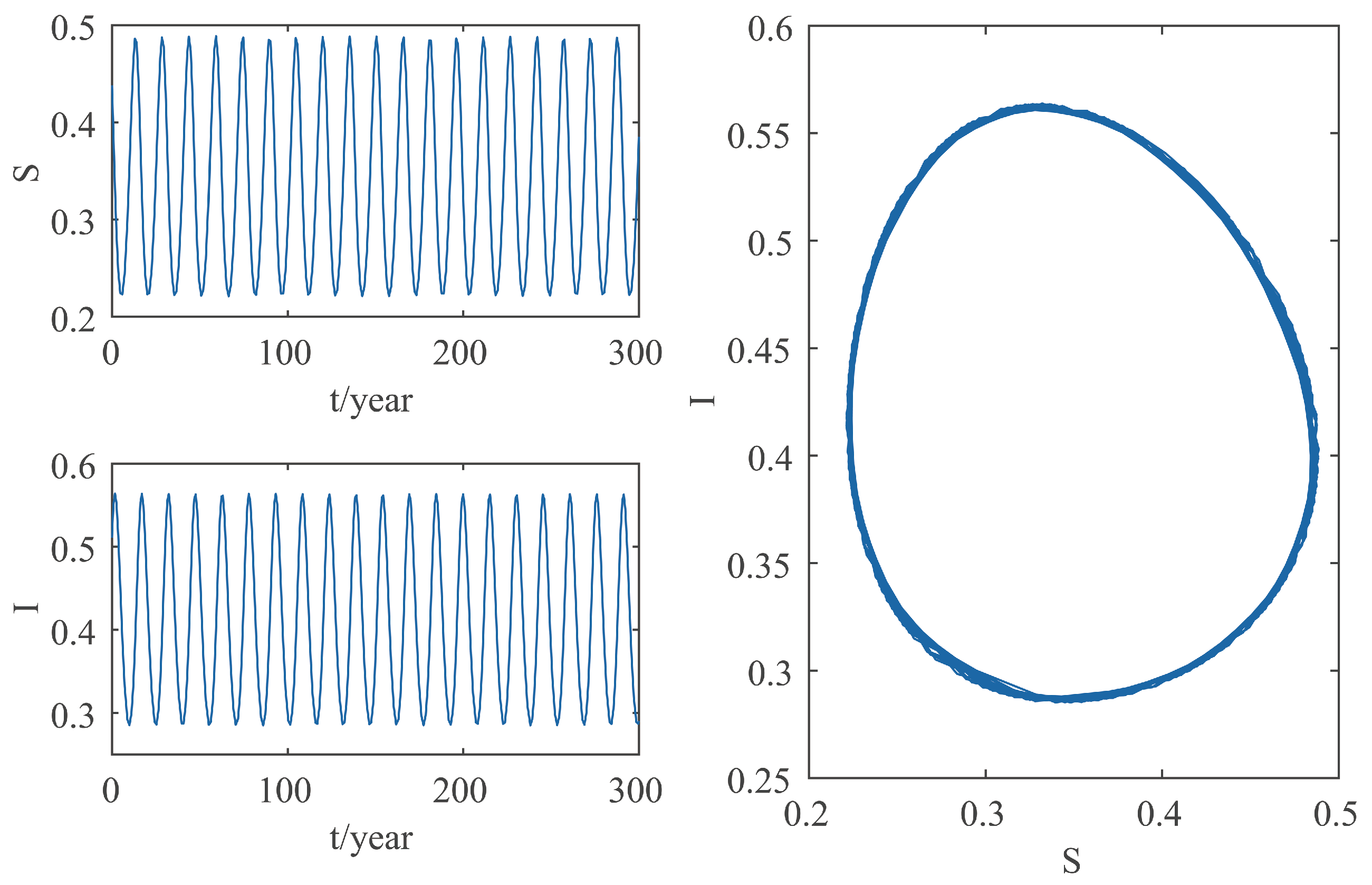
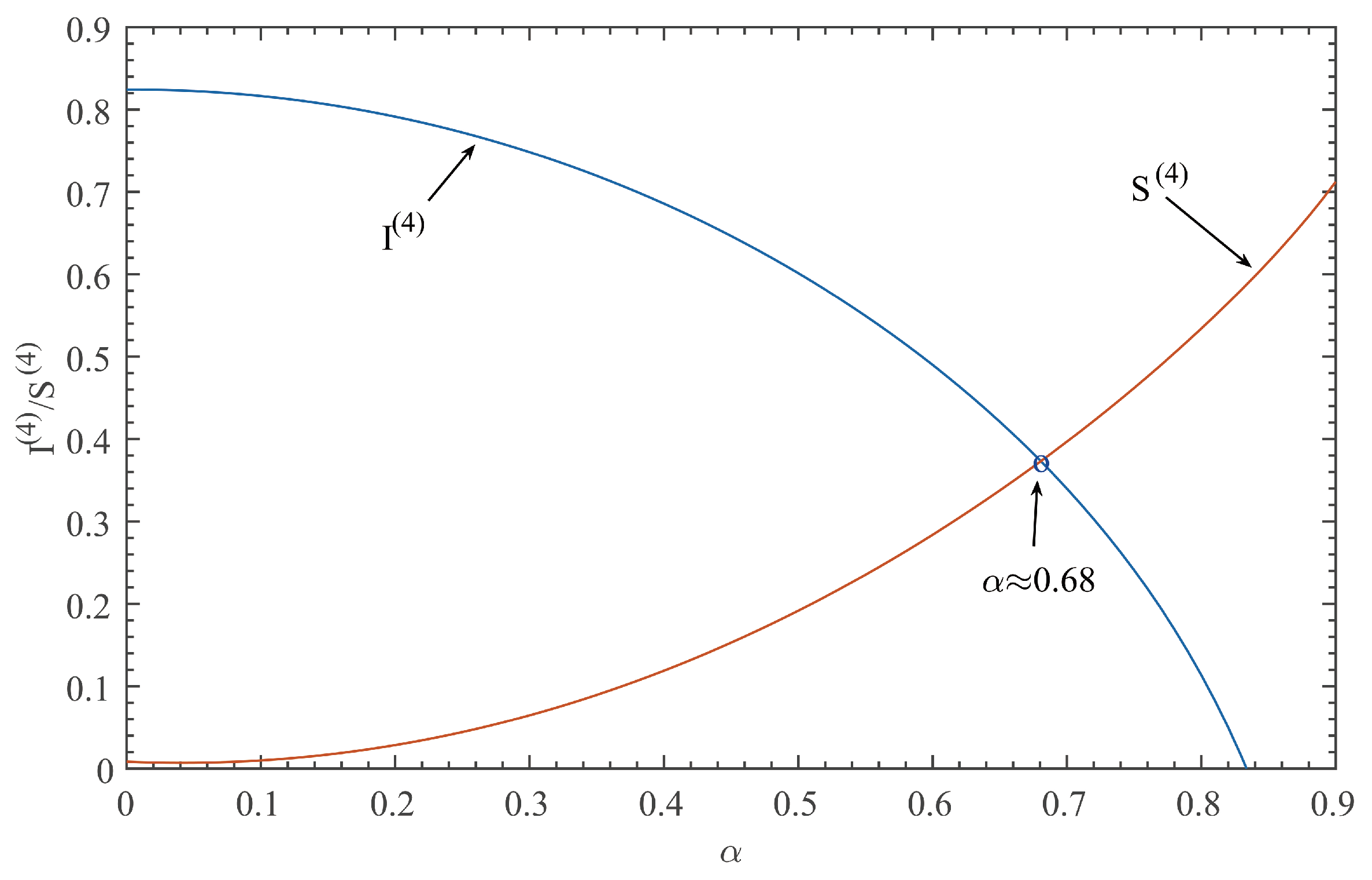
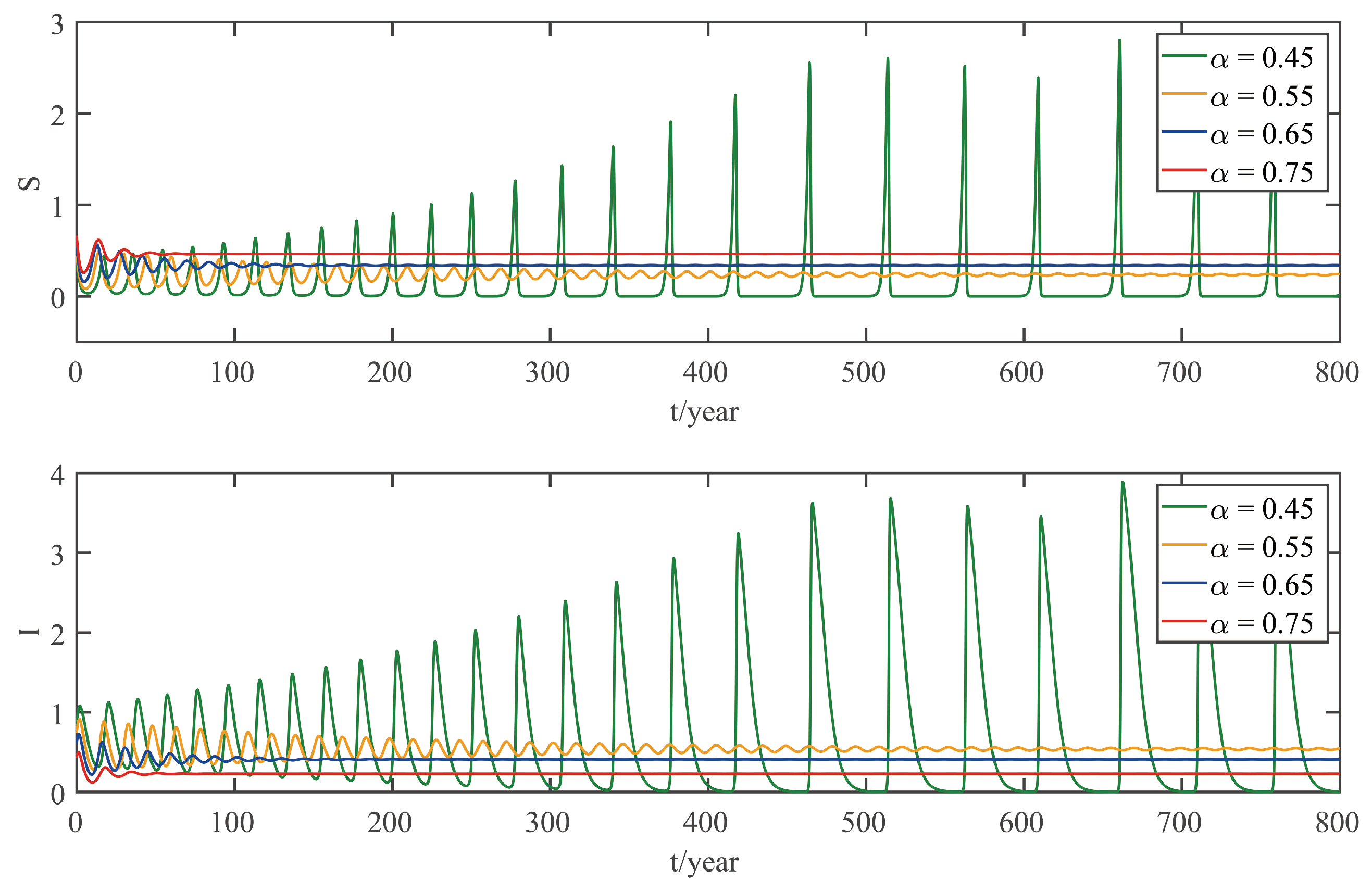
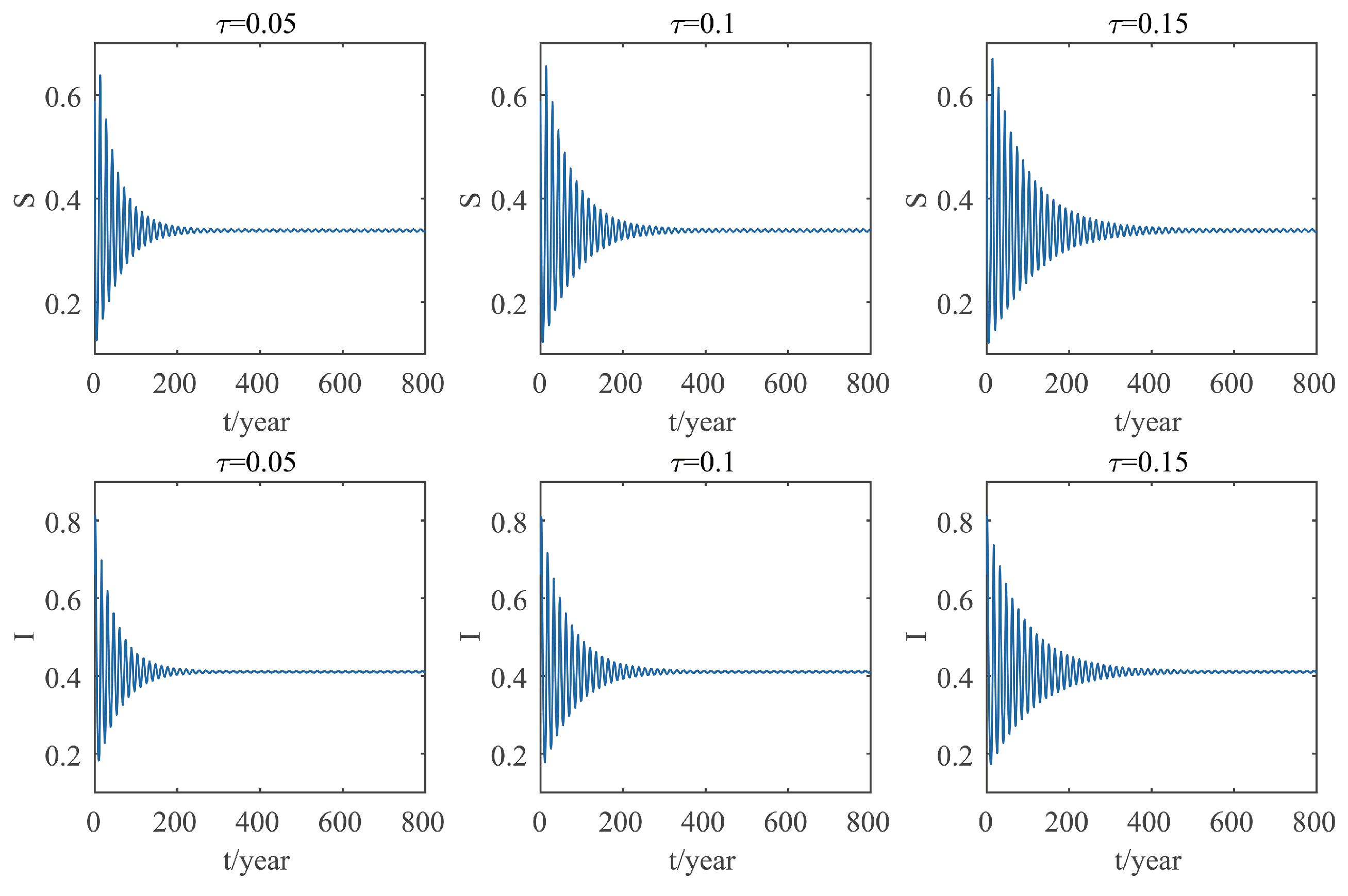
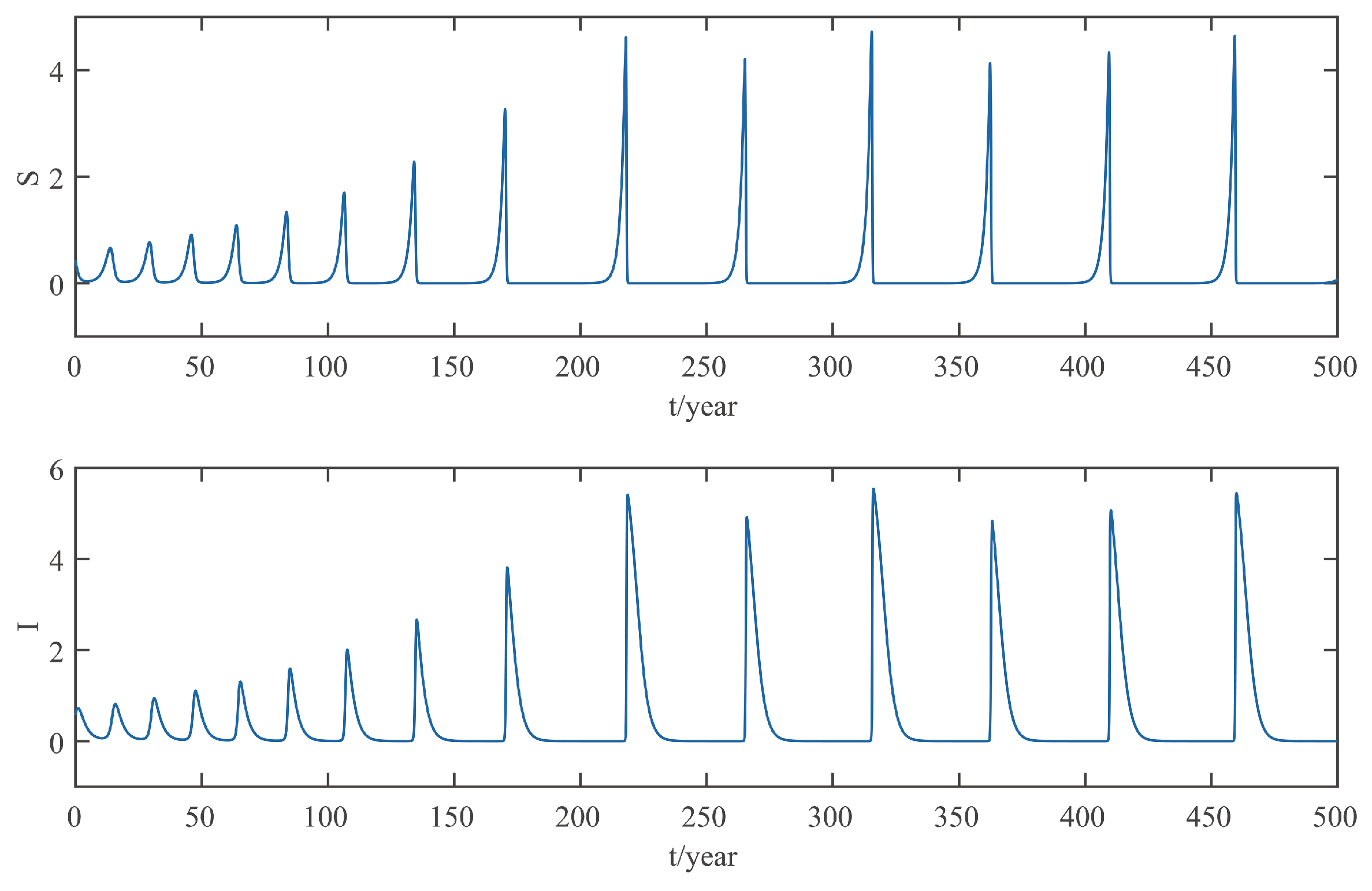
This paper focuses on the dynamics of the insect-vector populations based on SI epidemic model. By dividing M. alternatus into susceptible and infected, we have constructed a two-dimensional delay differential equation model considering the control intensity and the time delay for control to take effect. After that, we have analyzed the existence and stability of the equilibrium and the existence of Hopf bifurcation, and derived the normal form of Hopf bifurcation by using a multiple time scales method. Finally, by selecting scientific parameters for numerical simulation, the results of our theoretical analysis have been verified. Numerical analysis shows that when the intensity of control is large (obviously, if the intensity of control is large, then the environmental carrying capacity of M. alternatus will decrease accordingly), the disease-free equilibrium is always stable; when the intensity of control is 55%∼75%, the disease-free equilibrium is unstable for any , and oppositely, the positive equilibrium is stable before the critical time delay , and the system will occur stable Hopf bifurcation periodic solution near equilibrium . If the environmental carrying capacity of M. alternatus is large, the disease-free equilibrium does not exist and the positive equilibrium cannot reach stability, which provides a theoretical support for the prevention and control of PWD. However, in fact, there are many difficulties in the disease control. For example, the effect of control is affected by many factors, so it is unrealistic for the intensity of disease control to remain constant. In the process of modeling, we assume that the parameters are constant; in reality, the parameters are changing over time. However, in general, the stability of our model is consistent with the reality. Based on the stability analysis, more effective measures can be taken to reduce the damage caused by PWD.
In addition, our numerical analysis also shows that the number of infected M. alternatus decreases with the control intensity increasing, and the time for the system to reach stability increases with the time delay increasing. Therefore, it is important to increase the control intensity and shorten the time delay of control to take effect. Here, we suggest that in the process of prevention and control, we can choose combined measures to increase control intensity. Meanwhile, we suggest strengthening the monitoring of trees to take measures on trees as soon as possible to shorten the time delay. Moreover, when the the system eventually fails to reach stability, the disease outbreak shows apparent periodicity. In this way, we can better prevent the outbreak of PWD according to some rules, which can prolong the growth cycle of trees and reduce the loss of forest resources, and thus, improve the carbon sink capacity of forests and accelerate the realization of the goal of “emission peak and carbon neutrality”, so as to build a modernized country in which humanity and nature coexist in harmony.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}