
Cross-Diffusion-Induced Turing Instability in a Two-Prey One-Predator System


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Approach
2.2. A Model of a Two-Prey One-Predator Ecosystem and Its Parameters
- and are the population densities of three species.
- is a bounded domain in with a smooth boundary .
- Vector is the unit outward normal to .
- Coefficient is the diffusion rate of the i-th species. This diffusion term represents a simple Brownian-type motion of particle dispersal.
- is the cross-diffusion rate of the i-th species. It is necessary to note that the cross-diffusion coefficient may be positive or negative. The positive cross-diffusion coefficient represents that one species tends to move in the direction of a lower concentration of another species. On the contrary, the negative cross-diffusion coefficient denotes the population flux of one species in the direction of the higher concentration of another species. For instance, the predator diffuses with fluxAs , the part of the flux is directed toward the decreasing population density of the prey . Here, the cross-diffusion term presents the tendency of predators to avoid group defense by a large number of prey, i.e., the predator diffuses in the direction of the lower concentration of the prey species. More biological background can be found in [26,27,28].
3. Main Results
3.1. Stability of the Positive Equilibrium Solution of the ODE System
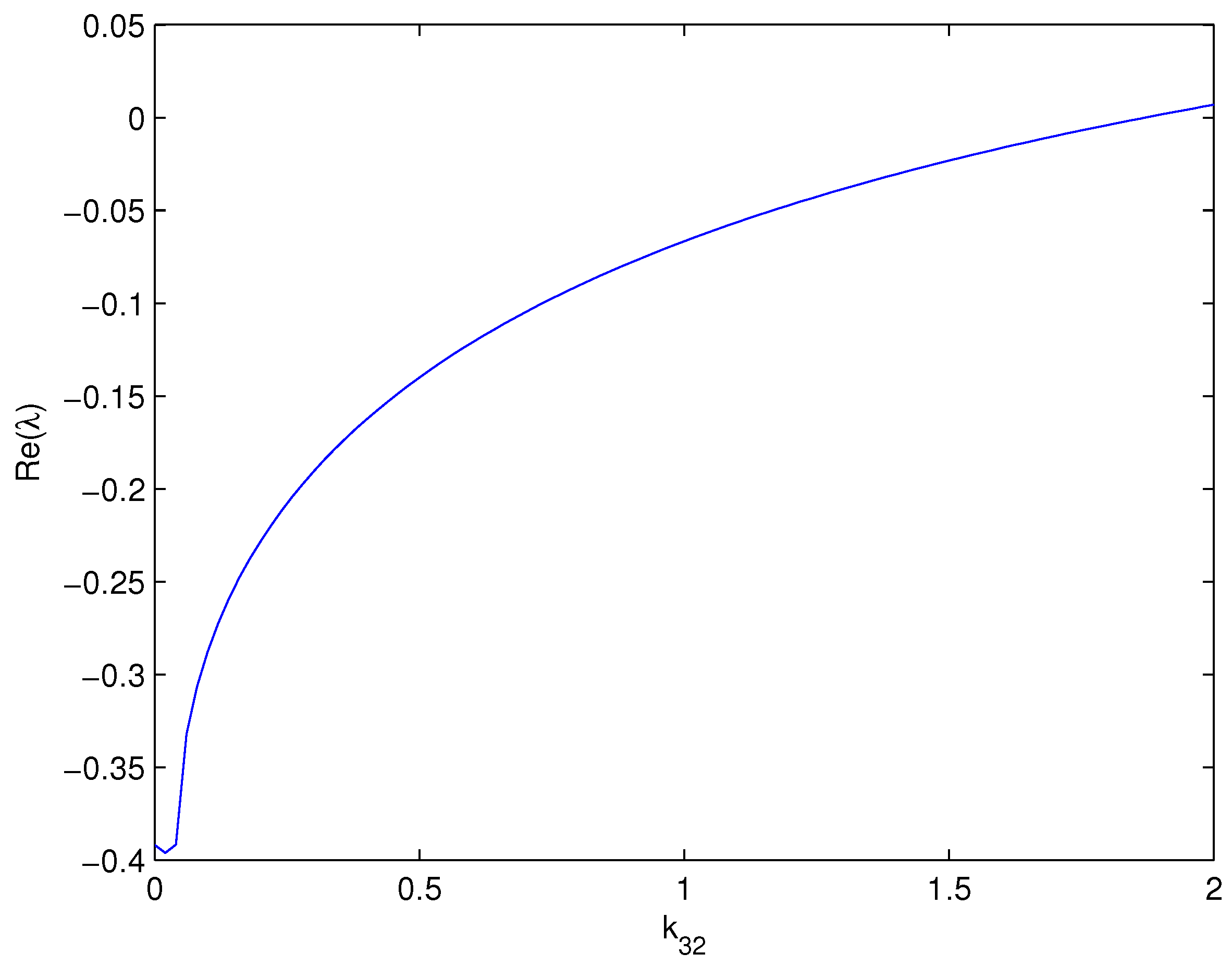
3.2. Effects of Cross-Diffusion on Turing Instability
- Suppose that . Consider as the variation parameter; then, there exists a positive constant such that when , the equilibrium is linearly unstable for some domain Ω.
- (1)
- (2)
- if
- (3)
- if
- (1)
- ,
- (2)
- ,
- (3)
- .
- Biological interpretation: In our model, the third species preys on the first and second. The positive steady state of the model can be broken by the reaction–diffusion among two species in the model.
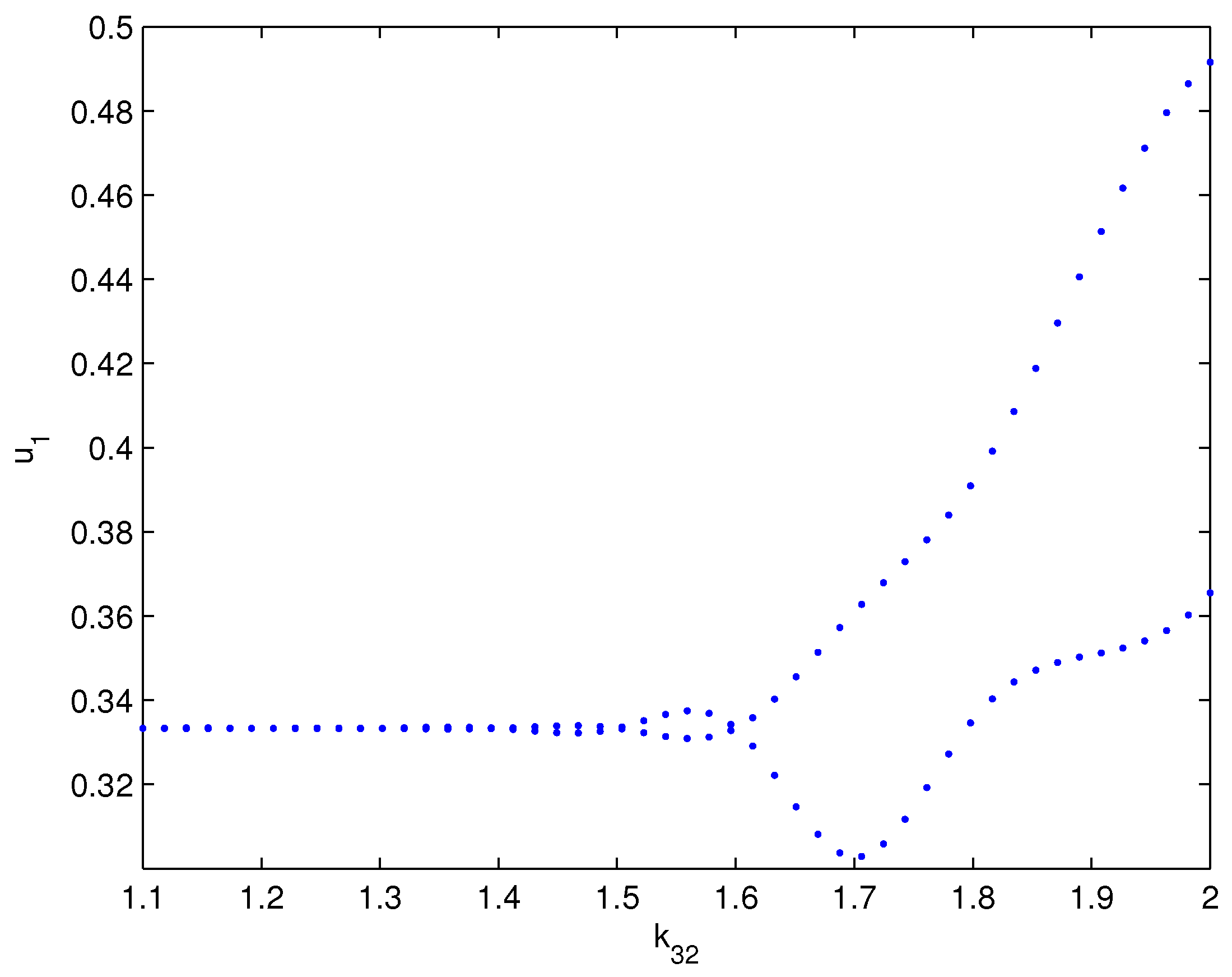
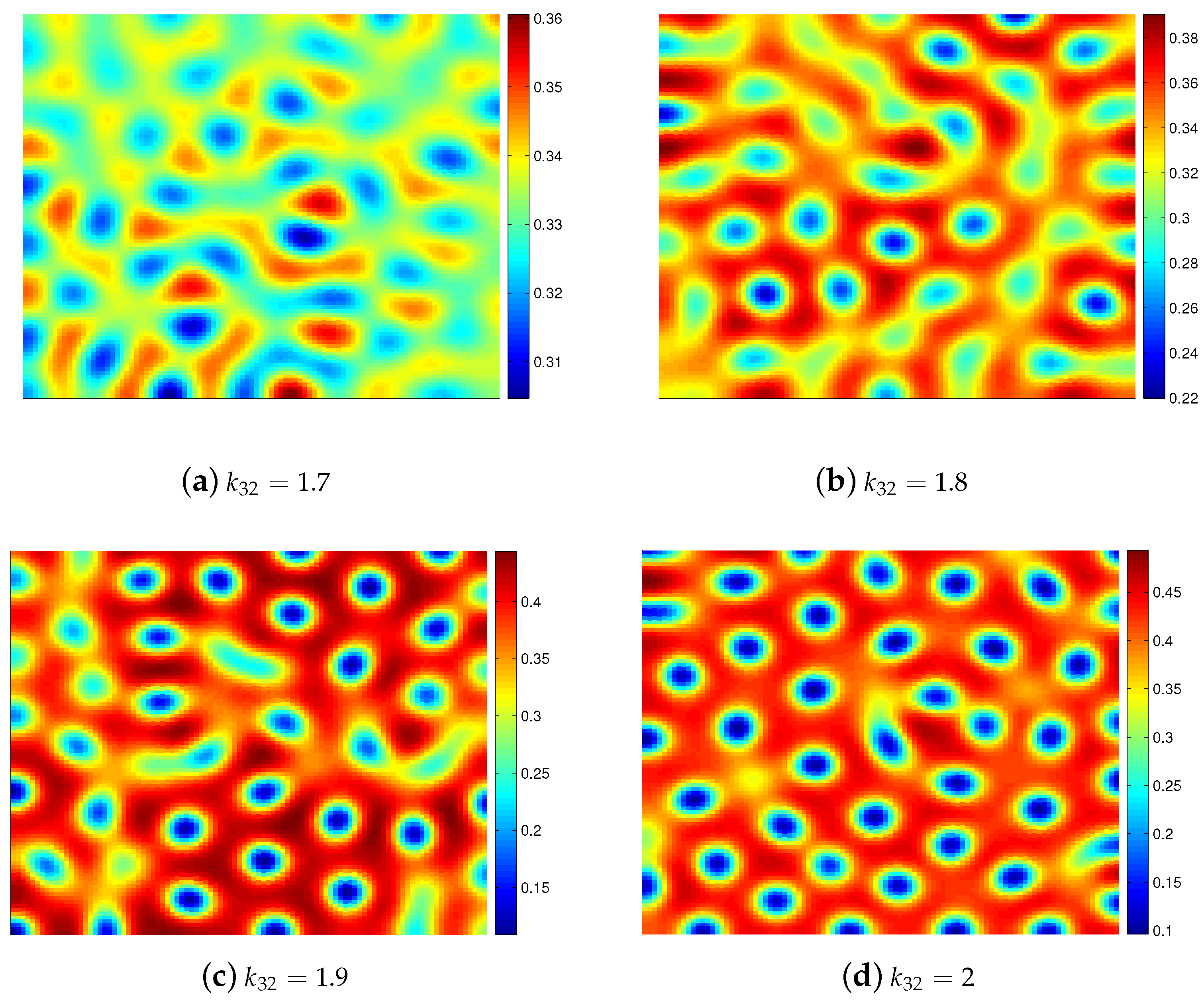
4. Numerical Simulations
5. Conclusions
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turing, A.M. The chemical basis of morphogenesis. Philos. Trans. R. Soc. Lond. Ser. B 1952, 237, 37–72. [Google Scholar]
- Cintra, W.; dos Santos, C.A.; Zhou, J.Z. Coexistence states of a Holling type II predator-prey system with self and cross-diffusion terms. Discret. Contin. Dyn. Syst. Ser. B 2022, 27, 3913–3931. [Google Scholar] [CrossRef]
- Farshid, M.; Jalilian, Y. Steady state bifurcation in a cross diffusion prey-predator model. Comput. Methods Differ. Equ. 2023, 11, 254–262. [Google Scholar] [CrossRef]
- Kuto, K. Stability of steady-state solutions to a prey-predator system with cross-diffusion. J. Differ. Equ. 2004, 197, 293–314. [Google Scholar] [CrossRef]
- Kuto, K.; Yamada, Y. Multiple coexistence states for a prey-predator system with cross-diffusion. J. Differ. Equ. 2004, 197, 315–348. [Google Scholar] [CrossRef]
- Li, Q.; He, J.F. Pattern formation in a ratio-dependent predator-prey model with cross diffusion. Electron. Res. Arch. 2023, 31, 1106–1118. [Google Scholar] [CrossRef]
- Ling, Z.; Zhang, L.; Lin, Z.G. Turing pattern formation in a predator-prey system with cross diffusion. Appl. Math. Model. 2014, 38, 5022–5032. [Google Scholar] [CrossRef]
- Ma, L.; Wang, H.T.; Gao, J.P. Dynamics of two-species Holling type-II predator-prey system with cross-diffusion. J. Differ. Equ. 2023, 365, 591–635. [Google Scholar] [CrossRef]
- Peng, R.; Wang, M.X.; Yang, G.Y. Stationary patterns of the Holling-Tanner prey-predator model with diffusion and cross-diffusion. Appl. Math. Comput. 2008, 196, 570–577. [Google Scholar] [CrossRef]
- Tao, Y.S.; Winkler, M. Existence theory and qualitative analysis for a fully cross-diffusive predator-prey system. SIAM J. Math. Anal. 2022, 54, 4806–4864. [Google Scholar] [CrossRef]
- Wang, M.X. Stationary patterns caused by cross-diffusion for a three-species prey-predator model. Comput. Math. Appl. 2006, 52, 707–720. [Google Scholar] [CrossRef]
- Xie, Z.F. Cross-diffusion induced Turing instability for a three species food chain model. J. Math. Anal. Appl. 2012, 388, 539–547. [Google Scholar] [CrossRef]
- Zhu, M.; Li, J.; Lian, X. Pattern Dynamics of Cross Diffusion Predator–Prey System with Strong Allee Effect and Hunting Cooperation. Mathematics 2022, 10, 3171. [Google Scholar] [CrossRef]
- Kerner, E.H. Further considerations on the statistical mechanics of biological associations. Bull. Math. Biophys. 1959, 21, 217–255. [Google Scholar] [CrossRef]
- Shigesada, N.; Kawasaki, K.; Teramoto, E. Spatial segregation of interacting species. J. Theoret. Biol. 1979, 79, 83–99. [Google Scholar] [CrossRef]
- Prokopev, S.; Lyubimova, T.; Mialdun, A.; Shevtsova, V. A ternary mixture at the border of Soret separation stability. Phys. Chem. Chem. Phys. 2021, 23, 8466–8477. [Google Scholar] [CrossRef] [PubMed]
- Šeta, B.; Errarte, A.; Ryzhkov, I.I.; Bou-Ali, M.M.; Shevtsova, V. Oscillatory instability caused by the interplay of Soret effect and cross-diffusion. Phys. Fluids 2023, 35, 021702. [Google Scholar] [CrossRef]
- Vanag, V.K.; Epstein, I.R. Cross-diffusion and pattern formation in reaction–diffusion systems. Phys. Chem. Chem. Phys. 2009, 11, 897–912. [Google Scholar] [CrossRef]
- Jorné, J. The diffusive Lotka-Volterra oscillating system. J. Theor. Biol. 1977, 65, 133–139. [Google Scholar] [CrossRef]
- Kersner, R.; Klincsik, M.; Zhanuzakova, D. A competition system with nonlinear cross-diffusion: Exact periodic patterns. Rev. R. Acad. Cienc. Exactas Fís. Nat. Ser. A Mat. 2022, 116, 187. [Google Scholar] [CrossRef]
- Matano, H.; Mimura, M. Pattern formation in competion-diffusion systems in nonconvex domains. Publ. Res. Inst. Math. Sci. 1983, 19, 1049–1079. [Google Scholar] [CrossRef]
- Gurtin, M.E. Some mathematical models for population dynamics that lead to segregation. Quart. Appl. Math. 1974, 32, 1–9. [Google Scholar] [CrossRef]
- Dhariwal, G.; Jüngel, A.; Zamponi, N. Global martingale solutions for a stochastic population cross-diffusion system. Stoch. Process Their Appl. 2019, 129, 3792–3820. [Google Scholar] [CrossRef]
- Yamada, Y. Global solutions for quasilinear parabolic systems with cross-diffusion effects. Nonlinear Anal. 1995, 24, 1395–1412. [Google Scholar] [CrossRef]
- Elettreby, M.F. Two-prey one-predator model. Chaos Solitons Fractals 2009, 39, 2018–2027. [Google Scholar] [CrossRef]
- Cantrell, R.S.; Cosner, C. Spatial Ecology via Reaction-Diffusion Equations; John Wiley & Sons, Ltd.: Chichester, UK, 2003. [Google Scholar]
- Lou, Y.; Ni, W.M. Diffusion, self-diffusion and cross-diffusion. J. Differ. Equ. 1996, 131, 79–131. [Google Scholar] [CrossRef]
- Okubo, A. Diffusion and Ecological Problems: Mathematical Models; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1980. [Google Scholar]
- Hale, J.K. Ordinary Differential Equations; Krieger: Malabar, FL, USA, 1980. [Google Scholar]
- Ermentrout, B. Stripes or spots? Non-linear effects in bifurcation of reaction-diffusion equations on the square. Proc. R. Soc. Lond. A 1991, 434, 413–417. [Google Scholar] [CrossRef]
- Guin, L.N.; Haque, M.; Mandal, P.K. The spatial patterns through diffusion-driven instability in a predator-prey model. Appl. Math. Model. 2012, 36, 1825–1841. [Google Scholar] [CrossRef]
- Sun, G.Q.; Jin, Z.; Li, L.; Haque, M.; Li, B.-L. Spatial patterns of a predator-prey model with cross diffusion. Nonlinear Dynam. 2012, 69, 1631–1638. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Chen, Y.; Zhou, Y. Cross-Diffusion-Induced Turing Instability in a Two-Prey One-Predator System. Mathematics 2023, 11, 2411. https://doi.org/10.3390/math11112411
Yu Y, Chen Y, Zhou Y. Cross-Diffusion-Induced Turing Instability in a Two-Prey One-Predator System. Mathematics. 2023; 11(11):2411. https://doi.org/10.3390/math11112411
Chicago/Turabian StyleYu, Ying, Yahui Chen, and You Zhou. 2023. "Cross-Diffusion-Induced Turing Instability in a Two-Prey One-Predator System" Mathematics 11, no. 11: 2411. https://doi.org/10.3390/math11112411