
A Study on the Low-Power Operation of the Spike Neural Network Using the Sensory Adaptation Method

Abstract
:1. Introduction
- We propose the sensory adaptation, which is closer to a biological mechanism than the existing frequency adaptation methods, and apply it to SNNs to develop SNNs with more biological meaningfulness.
- We show that improved SNNs have more biological meaningfulness and generate only a smaller number of firing spikes while maintaining accuracy, thereby demonstrating that the corresponding SNNs operate in lower power.
- We mathematically demonstrate the significance of the proposed method and develop a dedicated SNN simulator that can verify its superiority.
2. Spiking Neural Networks: A Preliminary
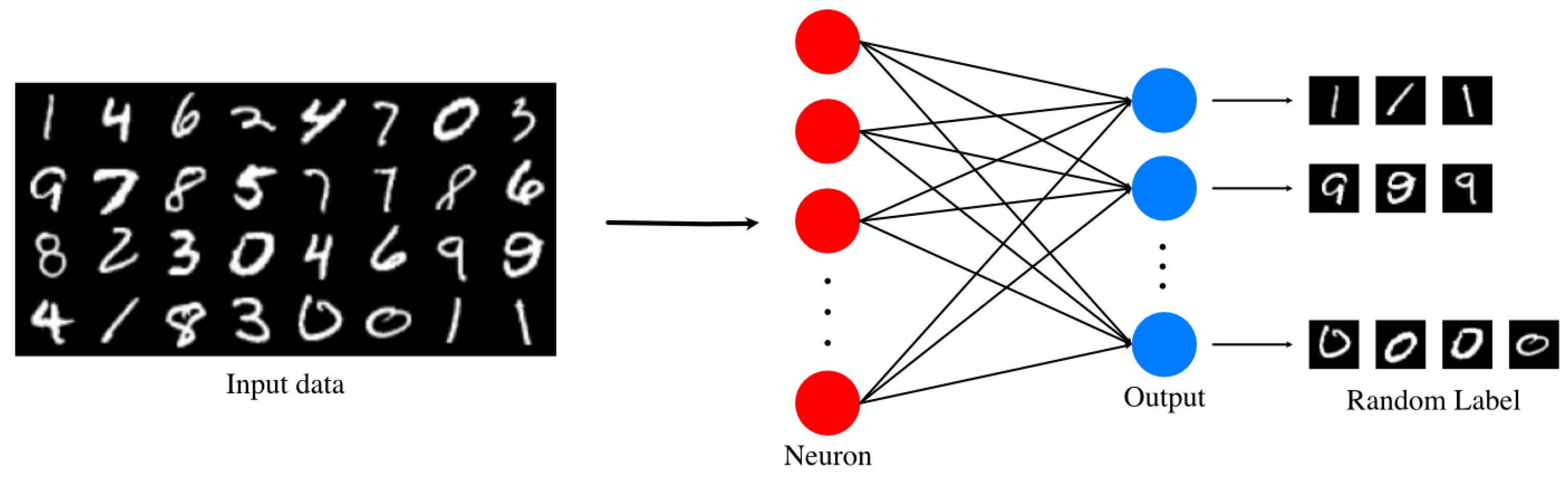
2.1. Unsupervised SNN
2.2. Neuron Model and Learning Method
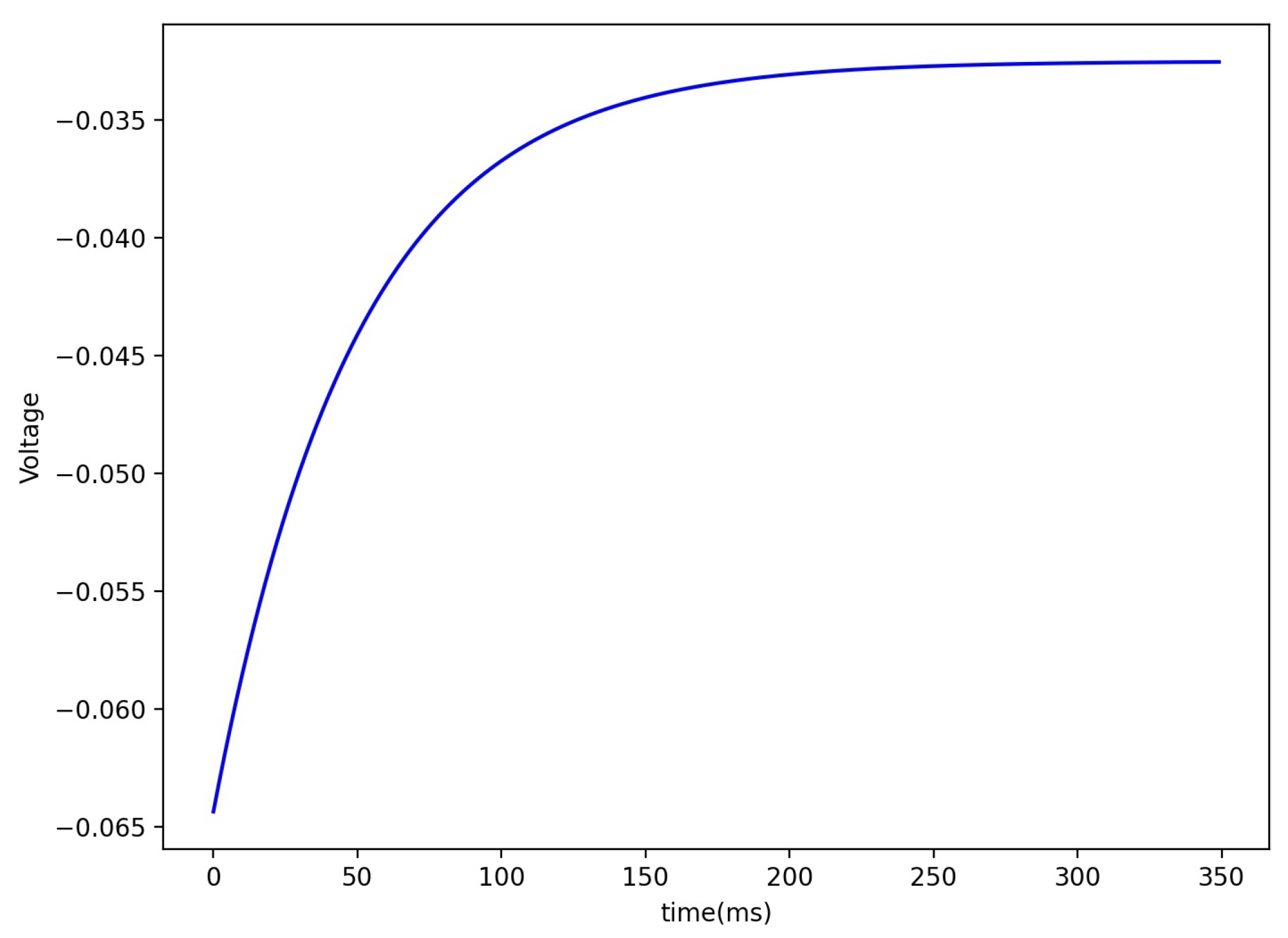
2.3. Spike Frequency Adaptation
3. SNN with Sensory Adaptation: The Proposed Method
- Sensitivity in the sensory adaptation is a completely different concept from discussed in Section 2, where represents the potential threshold of the neuron, and the sensitivity refers to the sensitivity of the receptor that the neuron receives the stimulus.
- in the proposed SNN with sensory adaptation does not change in any case.
3.1. Neuron Model and Network Architecture
3.2. Sensory Adaptation
4. Development of the SNN Simulator
5. Results
5.1. Experimental Works
5.2. Analysis
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Davies, M. Advancing Neuromorphic Computing from promise to Competitive technology. In Proceedings of the Neuro-Inspired Computational Elements Workshop (NICE), Albany, NY, USA, 26–28 March 2019. [Google Scholar]
- Merolla, P.A.; Arthur, J.V.; Alvarez-Icaza, R.; Cassidy, A.S.; Sawada, J.; Akopyan, F.; Jackson, B.L.; Imam, N.; Guo, C.; Nakamura, Y.; et al. A million spiking-neuron integrated circuit with a scalable communication network and interface. Science 2014, 345, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, B.V.; Gao, P.; McQuinn, E.; Choudhary, S.; Chandrasekaran, A.R.; Bussat, J.M.; Alvarez-Icaza, R.; Arthur, J.V.; Merolla, P.A.; Boahen, K. Neurogrid: A Mixed-Analog-Digital Multichip System for Large-Scale Neural Simulations. Proc. IEEE 2014, 102, 699–716. [Google Scholar] [CrossRef]
- Akopyan, F.; Sawada, J.; Cassidy, A.; Alvarez-Icaza, R.; Arthur, J.; Merolla, P.; Imam, N.; Nakamura, Y.; Datta, P.; Nam, G.J.; et al. TrueNorth: Design and Tool Flow of a 65 mW 1 Million Neuron Programmable Neurosynaptic Chip. IEEE Trans. Comput.-Aided Des. Integr. Circuits Syst. 2015, 34, 1537–1557. [Google Scholar] [CrossRef]
- Davies, M.; Srinivasa, N.; Lin, T.H.; Chinya, G.; Cao, Y.; Choday, S.H.; Dimou, G.; Joshi, P.; Imam, N.; Jain, S.; et al. Loihi: A Neuromorphic Manycore Processor with On-Chip Learning. IEEE Micro 2018, 38, 82–99. [Google Scholar] [CrossRef]
- Davies, M.; Wild, A.; Orchard, G.; Sandamirskaya, Y.; Guerra, G.A.F.; Joshi, P.; Plank, P.; Risbud, S.R. Advancing Neuromorphic Computing with Loihi: A Survey of Results and Outlook. Proc. IEEE 2021, 109, 911–934. [Google Scholar] [CrossRef]
- Khodaverdian, Z.; Sadr, H.; Edalatpanah, S.A. A Shallow Deep Neural Network for Selection of Migration Candidate Virtual Machines to Reduce Energy Consumption. In Proceedings of the 2021 7th International Conference on Web Research (ICWR), Tehran, Iran, 19–20 May 2021; pp. 191–196. [Google Scholar]
- Khodaverdian, Z.; Sadr, H.; Edalatpanah, S.A.; Solimandarabi, M.N. Combination of Convolutional Neural Network and Gated Recurrent Unit for Energy Aware Resource Allocation. arXiv 2021, arXiv:2106.12178. [Google Scholar]
- Shukla, R.; Khalilian, B.; Partouvi, S. Academic progress monitoring through neural network. Big Data Comput. Visions 2021, 1, 1–6. [Google Scholar]
- Peykani, P.; Eshghi, F.; Jandaghian, A.; Farrokhi-Asl, H.; Tondnevis, F. Estimating cash in bank branches by time series and neural network approaches. Big Data Comput. Visions 2021, 1, 170–178. [Google Scholar]
- Izhikevich, E. Simple model of spiking neurons. IEEE Trans. Neural Netw. 2003, 14, 1569–1572. [Google Scholar] [CrossRef] [Green Version]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500. [Google Scholar] [CrossRef]
- Morris, C.; Lecar, H. Voltage oscillations in the barnacle giant muscle fiber. Biophys. J. 1981, 35, 193–213. [Google Scholar] [CrossRef] [Green Version]
- Izhikevich, E. Which model to use for cortical spiking neurons? IEEE Trans. Neural Netw. 2004, 15, 1063–1070. [Google Scholar] [CrossRef]
- Peron, S.P.; Gabbiani, F. Role of spike-frequency adaptation in shaping neuronal response to dynamic stimuli. Biological cybernetics. Biol. Cybern. 2009, 100, 505–520. [Google Scholar] [CrossRef]
- Diehl, P.; Cook, M. Unsupervised learning of digit recognition using spike-timing-dependent plasticity. Front. Comput. Neurosci. 2015, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.R.; Alexiades, J.M.; Rajendran, B. Learning and real-time classification of hand-written digits with spiking neural networks. In Proceedings of the 2017 24th IEEE International Conference on Electronics, Circuits and Systems (ICECS), Tehran, Iran, 19–20 May 2017. [Google Scholar]
- Daqi Liu, S.Y. Fast unsupervised learning for visual pattern recognition using spike timing dependent plasticity. Neurocomputing 2017, 249, 212–224. [Google Scholar]
- Kang, T.; Oh, K.I.; Lee, J.J.; Kim, S.E.; Kim, S.E.; Lee, W.; Oh, W. Spiking Neural Networks-Inspired Signal Detection Based on Measured Body Channel Response. IEEE Trans. Instrum. Meas. 2022, 71, 1–16. [Google Scholar] [CrossRef]
- Kang, T.; Hwang, J.H.; Kim, H.; Kim, S.E.; Oh, K.I.; Lee, J.J.; Park, H.I.; Kim, S.E.; Oh, W.; Lee, W. Measurement and Evaluation of Electric Signal Transmission Through Human Body by Channel Modeling, System Design, and Implementation. IEEE Trans. Instrum. Meas. 2021, 70, 1–14. [Google Scholar] [CrossRef]
- Barbier, T.; Teulière, C.; Triesch, J. Spike timing-based unsupervised learning of orientation, disparity, and motion representations in a spiking neural network. In Proceedings of the 2021 IEEE/CVF Conference on Computer Vision and Pattern Recognition Workshops (CVPRW), Nashville, TN, USA, 19–25 June 2021; pp. 1377–1386. [Google Scholar]
- Kheradpisheh, S.R.; Ganjtabesh, M.; Thorpe, S.J.; Masquelier, T. STDP-based spiking deep convolutional neural networks for object recognition. Neural Netw. 2018, 99, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Ha, G.E.; Lee, J.; Kwak, H.; Song, K.; Kwon, J.; Jung, S.Y.; Hong, J.; Chang, G.E.; Hwang, E.M.; Shin, H.S.; et al. The Ca2+-activated chloride channel anoctamin-2 mediates spike-frequency adaptation and regulates sensory transmission in thalamocortical neurons. Nat. Commun. 2016, 7, 13791. [Google Scholar] [CrossRef]
- Morrison, A.; Aertsen, A.; Diesmann, M. Spike-Timing-Dependent Plasticity in Balanced Random Networks. Neural Comput. 2007, 19, 1437–1467. [Google Scholar] [CrossRef]
- Nessler, B.; Pfeiffer, M.; Buesing, L.; Maass, W. Bayesian Computation Emerges in Generic Cortical Microcircuits through Spike-Timing-Dependent Plasticity. PLoS Comput. Biol. 2013, 9, e1003037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfister, J.P.; Gerstner, W. Triplets of Spikes in a Model of Spike Timing-Dependent Plasticity. J. Neurosci. 2006, 26, 9673–9682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.; Miller, K.D.; Abbott, L.F. Competitive Hebbian learning through spike-timing-dependent synaptic plasticity. Nat. Neurosci. 2000, 3, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Bi, G.q.; Poo, M.m. Synaptic Modifications in Cultured Hippocampal Neurons: Dependence on Spike Timing, Synaptic Strength, and Postsynaptic Cell Type. J. Neurosci. 1998, 18, 10464–10472. [Google Scholar] [CrossRef] [Green Version]
- Doborjeh, M.; Doborjeh, Z.; Merkin, A.; Bahrami, H.; Sumich, A.; Krishnamurthi, R.; Medvedev, O.N.; Crook-Rumsey, M.; Morgan, C.; Kirk, I.; et al. Personalised predictive modelling with brain-inspired spiking neural networks of longitudinal MRI neuroimaging data and the case study of dementia. Neural Netw. 2021, 144, 522–539. [Google Scholar] [CrossRef]
- Guo, S.; Wang, L.; Wang, S.; Deng, Y.; Yang, Z.; Li, S.; Xie, Z.; Dou, Q. A Systolic SNN Inference Accelerator and Its Co-Optimized Software Framework. In Proceedings of the 2019 on Great Lakes Symposium on VLSI, Tysons Corner, VA, USA, 9–11 May 2019; pp. 63–68. [Google Scholar]
- Li, S.; Zhang, Z.; Mao, R.; Xiao, J.; Chang, L.; Zhou, J. A Fast and Energy-Efficient SNN Processor With Adaptive Clock/Event-Driven Computation Scheme and Online Learning. IEEE Trans. Circuits Syst. Regul. Pap. 2021, 68, 1543–1552. [Google Scholar] [CrossRef]
- Querlioz, D.; Bichler, O.; Dollfus, P.; Gamrat, C. Immunity to Device Variations in a Spiking Neural Network with Memristive Nanodevices. IEEE Trans. Nanotechnol. 2013, 12, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Kang, T.; Lee, J.J.; Han, K.; Kim, J.; Pedram, M. TEI-ULP: Exploiting Body Biasing to Improve the TEI-Aware Ultralow Power Methods. IEEE Trans. Comput.-Aided Des. Integr. Circuits Syst. 2019, 38, 1758–1770. [Google Scholar] [CrossRef]
- Han, K.; Lee, S.; Lee, J.J.; Lee, W.; Pedram, M. TIP: A Temperature Effect Inversion-Aware Ultra-Low Power System-on-Chip Platform. In Proceedings of the 2019 IEEE/ACM International Symposium on Low Power Electronics and Design (ISLPED), Lausanne, Switzerland, 29–31 July 2019; pp. 1–6. [Google Scholar]
- Han, K.; Lee, S.; Oh, K.I.; Bae, Y.; Jang, H.; Lee, J.J.; Lee, W.; Pedram, M. Developing TEI-Aware Ultralow-Power SoC Platforms for IoT End Nodes. IEEE Internet Things J. 2021, 8, 4642–4656. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description |
|---|---|
| resting membrane potential | |
| excitatory postsynaptic potential | |
| inhibitory postsynaptic potential | |
| excitatory conductance time constant | |
| inhibitory conductance time constant | |
| the conductance associated with the excitatory neuron | |
| the conductance associated with the inhibitory neuron |
| Param. | Value | Param. | Value |
|---|---|---|---|
| 0.065 V | 0.25 V | ||
| 0.06 V | 100 ms | ||
| 0.08 V | 10 ms | ||
| 0.075 V | 1 ms | ||
| 0.055 V | 2 ms | ||
| 0V | 1 nS |
| Accuracy | Number of Fired Spikes | |
|---|---|---|
| Conventional method | 80.52% | 446,722 |
| Proposed method | 83.56% | 242,904 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, M.; Kang, T.; Lee, J.-J.; Lee, W. A Study on the Low-Power Operation of the Spike Neural Network Using the Sensory Adaptation Method. Mathematics 2022, 10, 4191. https://doi.org/10.3390/math10224191
Jeon M, Kang T, Lee J-J, Lee W. A Study on the Low-Power Operation of the Spike Neural Network Using the Sensory Adaptation Method. Mathematics. 2022; 10(22):4191. https://doi.org/10.3390/math10224191
Chicago/Turabian StyleJeon, Mingi, Taewook Kang, Jae-Jin Lee, and Woojoo Lee. 2022. "A Study on the Low-Power Operation of the Spike Neural Network Using the Sensory Adaptation Method" Mathematics 10, no. 22: 4191. https://doi.org/10.3390/math10224191