

Role of Glucose Risk Factors on Human Breast Cancer: A Nonlinear Dynamical Model Evaluation

Abstract
:1. Introduction
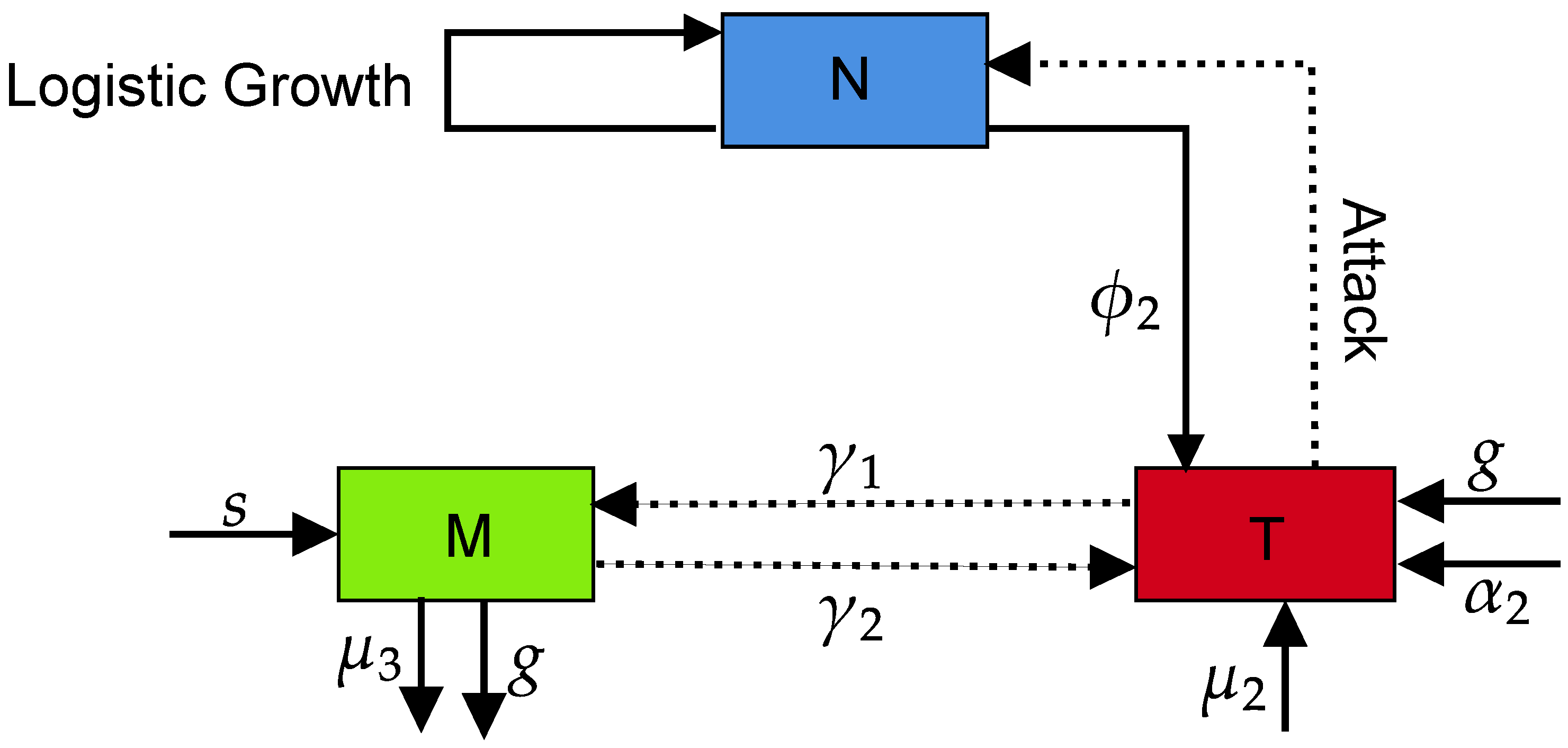
2. Formulation of Nonlinear Dynamical Model for Breast Cancer
- The category of host (normal) epithelial cells () at any time (t) is composed of the breast tissues. The life cycle of follows the logistic growth , where and are the corresponding growth and death rate of the normal breast cells, respectively.
- The category of tumor cells () and their concentration is entirely different from the normal cells. The cell divisions in this class occur rapidly in an uncontrolled manner. The natural death of tumor cells has no biological meaning; however, their growth can be inhibited due to some factors such as immune suppression. The growth of tumor cells is described by , wherein and are the corresponding growth and inhibition rate of the breast tumor cells, respectively.
- The reasons for the appearance of the tumor cells in tissues are usually unknown. However, many studies argued that the occurrence of tumors may be due to various internal and external factors [23]. The factors such as cell damage, some hormones, immune cell weakness and others are classified as internal types. Conversely, the external factors include lifestyle, sleeping habits, stress, and exercise activity. These factors may transform a normal cell into a tumor cell. It is further assumed that the normal cells compete strongly with the tumor cells to gain space and energy resources in a tiny volume, justifying our acceptance of the competition model proposed by Mufudza et al. [9]. In essence, the interaction between tumor and normal cells can be represented as , where is the competition-induced death rate of the normal cells.
- Following the recommendation of [7], wherein due to the competition between tumor and normal cells, the former ones (tumor) grow rapidly at the expense of later ones (normal), one can write . Herein, denotes the competition-induced growth rate of the tumor cells.
- The human immune system is mainly responsible for the protection of the body from the growth of tumor cells, which are generated and die on daily basis [24,25]. The immune cells () in the absence of a tumor can be written as , where s and are the corresponding growth and natural death rates of the immune cells.
- The activity status of the tumors can boost the human immune system. A positive nonlinear growth term of the form , where and are the respective immune response and threshold rates (inversely proportional to the steepness of the immune response curve) related to the immune cells, can be used to represent such activity status.
- Both cells (tumor and immune cells) can fight with others, but immune cells have the capacity to fight with the foreign cells only. Hence, the tumor and immune cells might be reduced during the interaction process. We denote the competition via and , wherein is the reduction of the tumor cells by the immune cells action and is the decrease of the immune cells by the tumor cells. It is worth noting that the ability of the immune cells to invite the fringe can be affected by various factors such as excess blood glucose rate, impacting the immune system efficacy [20,21] written as .
3. Model Analysis
3.1. Boundedness and Positive Invariance
3.2. Existence of Equilibrium Points
3.2.1. Tumor-Free Equilibrium :
3.2.2. Type 1 Dead Equilibrium
3.2.3. Type 2 Dead Equilibrium ():
3.2.4. Coexisting Equilibrium :
- For , we achieve one real root and two imaginary roots.
- For , one achieves three real roots.
- For and , we obtain one simple real root with a multiplicity of three.
- For and , one obtains thee real roots with single and double multiplicity.
3.3. Local Stability of Equilibrium
- Asymptotically stable (sink) if
- Unstable (saddle) if
- Nonhyperbolic if .
4. Bifurcation
4.1. Zero Bifurcation
- (A1)
- The Jacobian matrix = has a simple one zero eigenvalue with eigenvector υ, and has an eigenvector w corresponding to the eigenvalue .
- (A2)
- has l eigenvalues with negative real parts and eigenvalues with positive real parts.
- (A3)
- (A4)
- (A5)
4.2. Hopf Bifurcation
5. Numerical Simulation of the Proposed Model
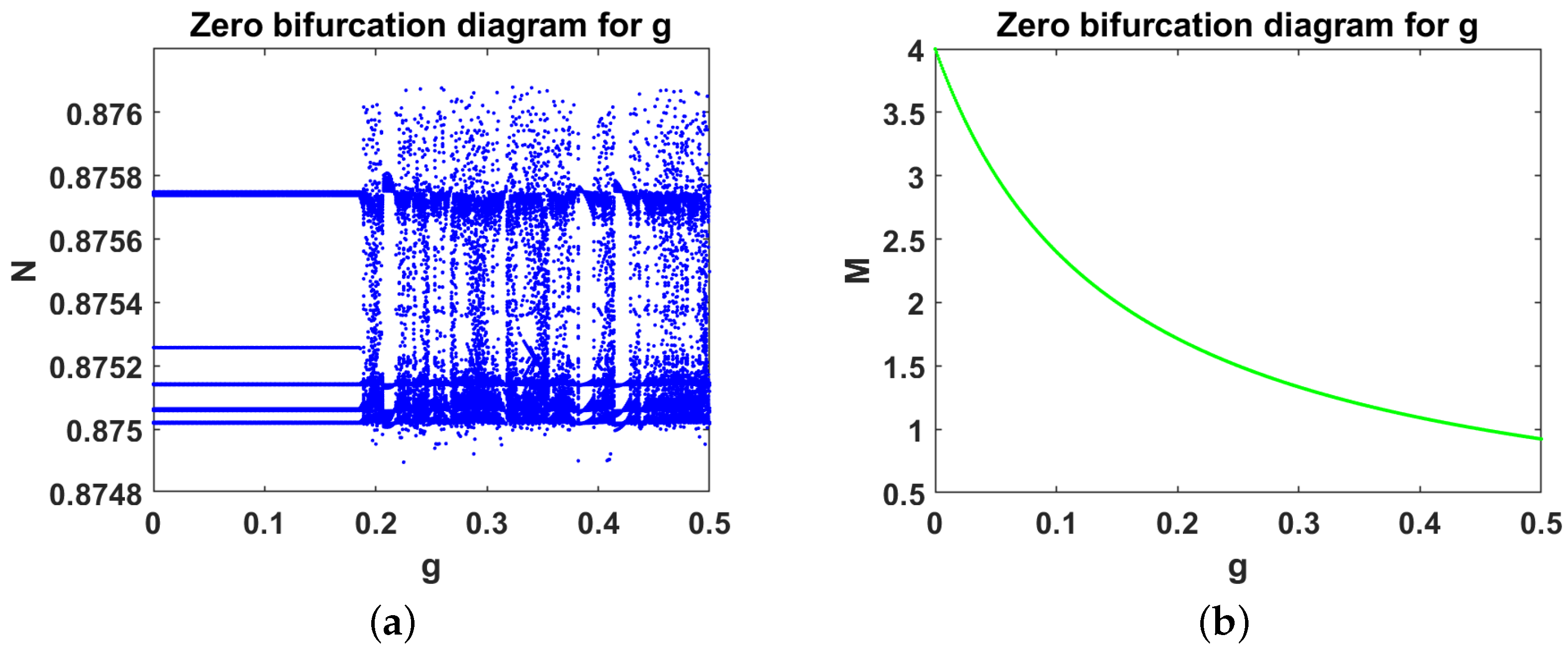
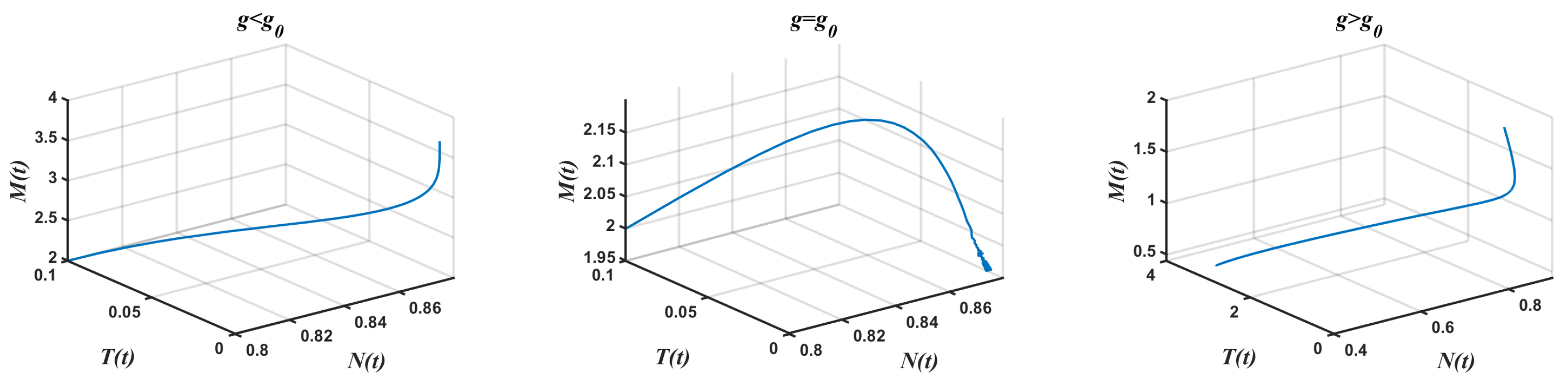
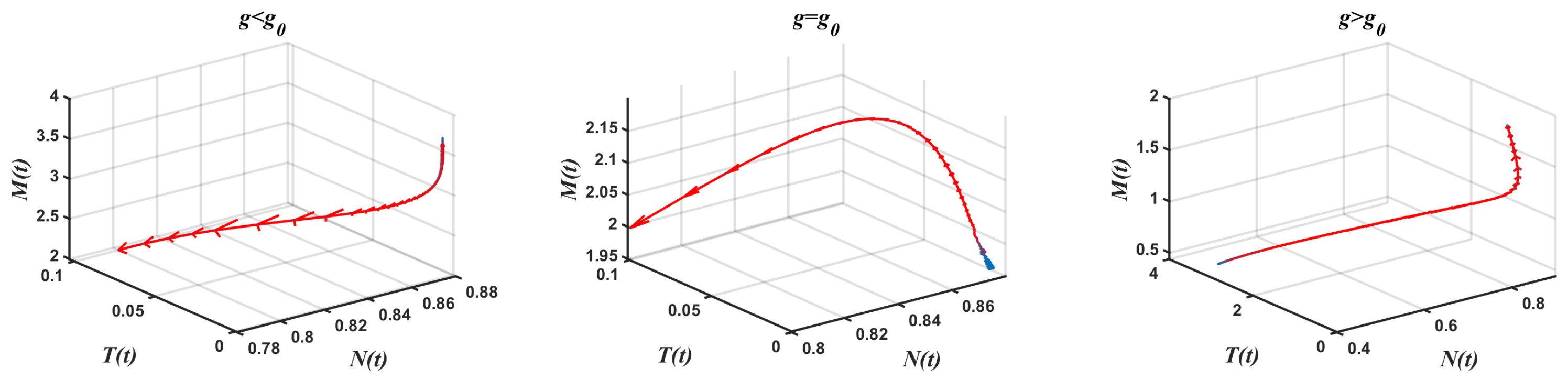
- Case 1.
- Complete absence of tumor cells We took and varied g. Theorem 6 was used to obtain . Equation (16) was used to obtain with , thereby achieving a free equilibrium point . The associated Jacobian matrix produced a zero eigenvalue and two negative eigenvalues Theorem 6 was used in the model Equation (1) to obtain a generic saddle-node bifurcation for Interestingly, changed its stability via the transcritical bifurcation when g crossed the critical value
- Case 2.
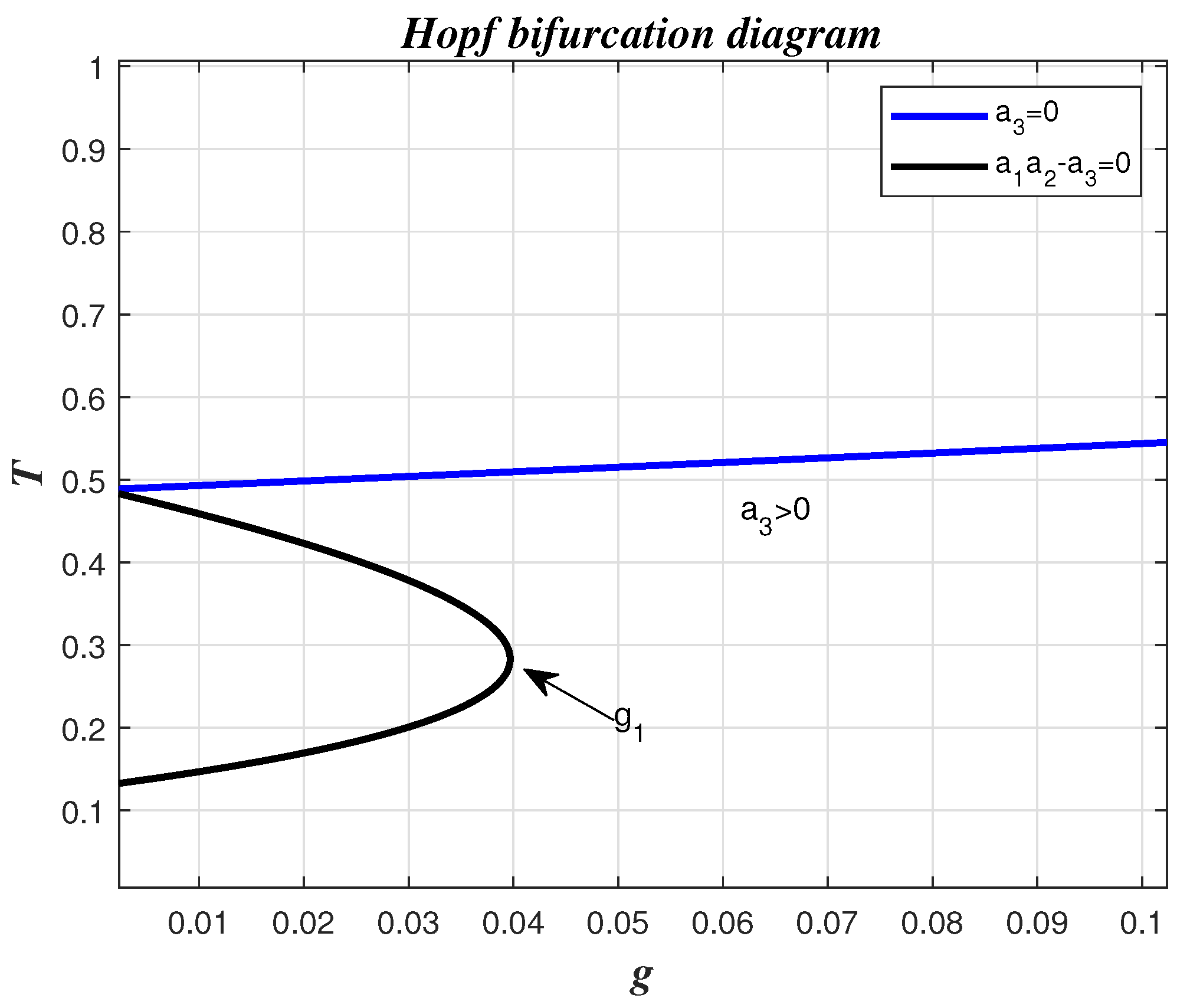
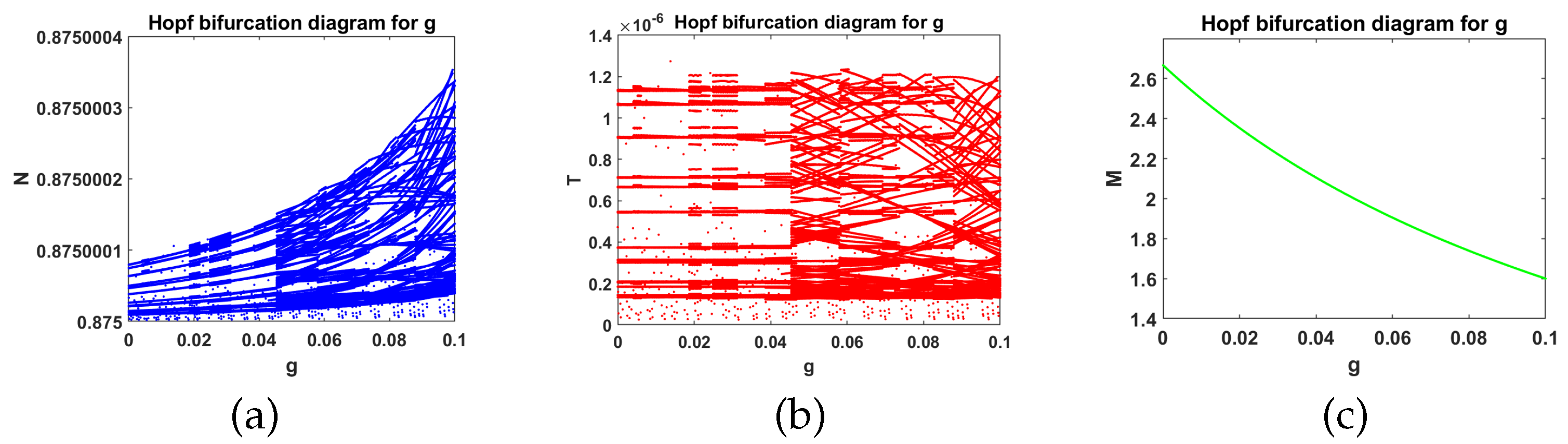
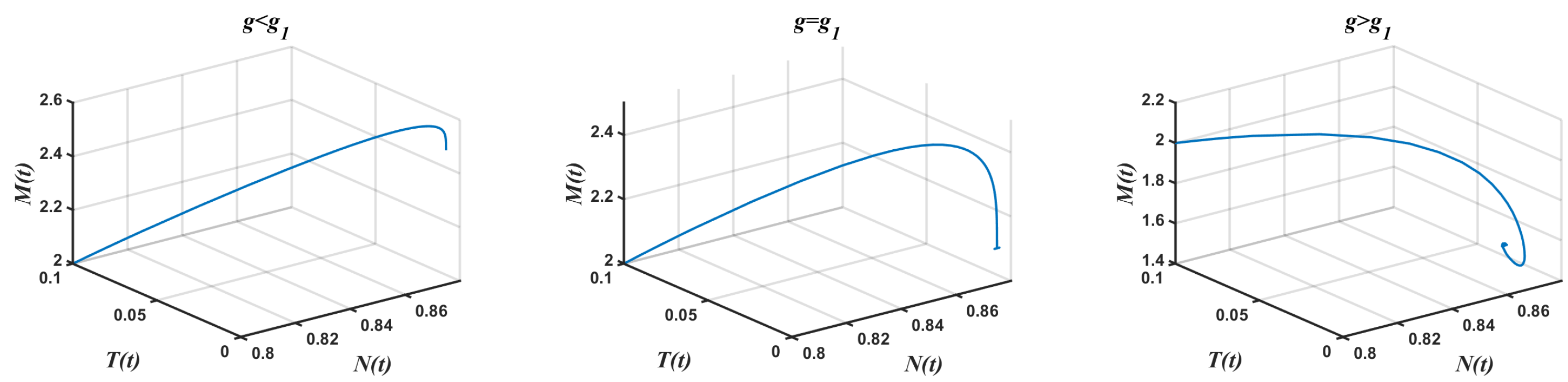
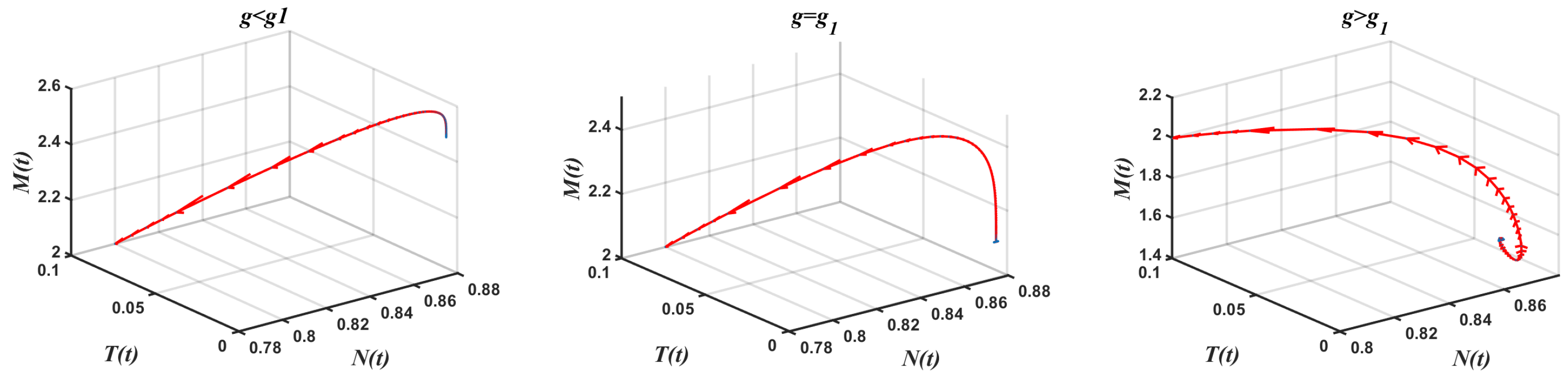
- In the presence of tumor cells In this case, the simulation is carried out by taking , , and other parameters have the same fixed values as indicated in Table 1. According to Theorem 9, had a pair of pure imaginary eigenvalues on the imaginary axis, and other roots lie on the left half-plane when the conditions given by Equation (33) are violated. The achieved critical Hopf bifurcation satisfied Equation (34) at . In addition, the coexistence equilibrium point was obtained. The Jacobian matrix had a negative real eigenvalue with a pair of pure imaginary eigenvalues According to Theorem 9, the model Equation (1) produced a Hopf bifurcation when Furthermore, changed its stability via the Hopf bifurcation when g crossed the critical parameterFigure 5 illustrates the Hopf bifurcation diagram for The area under the blue line satisfied the solutions of the inequalities and given in Equation (34). The critical value of was positioned on the black line inside the region in which in Equation (34) was satisfied. In brief, the emergence of Hopf bifurcation at for was clearly evidenced.
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dolatkhah, M.; Hashemzadeh, N.; Barar, J.; Adibkia, K.; Aghanejad, A.; Barzegar-Jalali, M.; Omidi, Y. Graphene-based multifunctional nanosystems for simultaneous detection and treatment of breast cancer. Colloids Surf. B Biointerfaces 2020, 193, 111104. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.M.; Hussain, F. Higher glucose enhances breast cancer cell aggressiveness. Nutr. Cancer 2020, 72, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, A.Y.; Spolador, L.H.; de Sousa Pereira, N.; Bocchi, M.; Fernandes, C.Y.; de Oliveira Pinsetta, M.; Munuera, M.D.; Moretto, S.L.; Fuzinatto, I.M.; de Castro, V.D.; et al. Breast cancer and nutrition: Interplay between diet and cancer prevention. Biosaúde 2020, 21, 87–99. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ku-Carrillo, R.A.; Delgadillo, S.E.; Chen-Charpentier, B.M. A mathematical model for the effect of obesity on cancer growth and on the immune system response. Appl. Math. Model. 2016, 40, 4908–4920. [Google Scholar] [CrossRef]
- Dehingia, K.; Yao, S.W.; Sadri, K.; Das, A.; Sarmah, H.K.; Zeb, A.; Inc, M. A study on cancer-obesity-treatment model with quadratic optimal control approach for better outcomes. Results Phys. 2022, 42, 105963. [Google Scholar] [CrossRef]
- Alharbi, S.A.; Rambely, A.S. Dynamic behaviour and stabilisation to boost the immune system by complex interaction between tumour cells and vitamins intervention. Adv. Differ. Equ. 2020, 2020, 412. [Google Scholar] [CrossRef]
- Admon, M.R.; Maan, N. Modelling tumor growth with immune response and drug using ordinary differential equations. Jurnal Teknologi 2017, 79, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Mufudza, C.; Sorofa, W.; Chiyaka, E.T. Assessing the effects of estrogen on the dynamics of breast cancer. Comput. Math. Methods Med. 2012, 2012, 473572. [Google Scholar] [CrossRef] [Green Version]
- Oke, S.I.; Matadi, M.B.; Xulu, S.S. Optimal control analysis of a mathematical model for breast cancer. Math. Comput. Appl. 2018, 23, 21. [Google Scholar]
- Debbouche, N.; Ouannas, A.; Grassi, G.; Al-Hussein, A.B.A.; Tahir, F.R.; Saad, K.M.; Jahanshahi, H.; Aly, A.A. Chaos in Cancer Tumor Growth Model with Commensurate and Incommensurate Fractional-Order Derivatives. Comput. Math. Methods Med. 2022, 2022, 5227503. [Google Scholar] [CrossRef] [PubMed]
- Abd-Rabo, M.A.; Zakarya, M.; Alderremy, A.A.; Aly, S. Dynamical analysis of tumor model with obesity and immunosuppression. Alex. Eng. J. 2022, 61, 10897–10911. [Google Scholar] [CrossRef]
- Fadaka, A.; Ajiboye, B.; Ojo, O.; Adewale, O.; Olayide, I.; Emuowhochere, R. Biology of glucose metabolization in cancer cells. J. Oncol. Sci. 2017, 3, 45–51. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.C.; Hsu, L.L.; Wang, S.F.; Hsu, C.Y.; Lee, H.C.; Tseng, L.M. ROS Mediate xCT-Dependent Cell Death in Human Breast Cancer Cells under Glucose Deprivation. Cells 2020, 9, 1598. [Google Scholar] [CrossRef]
- Wardi, L.; Alaaeddine, N.; Raad, I.; Sarkis, R.; Serhal, R.; Khalil, C.; Hilal, G. Glucose restriction decreases telomerase activity and enhances its inhibitor response on breast cancer cells: Possible extra-telomerase role of BIBR 1532. Cancer Cell Int. 2014, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Krętowski, R.; Borzym-Kluczyk, M.; Stypułkowska, A.; Brańska-Januszewska, J.; Ostrowska, H.; Cechowska-Pasko, M. Low glucose dependent decrease of apoptosis and induction of autophagy in breast cancer MCF-7 cells. Mol. Cell. Biochem. 2016, 417, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, A.M.; Martel, F. Targeting glucose transporters for breast cancer therapy: The effect of natural and synthetic compounds. Cancers 2020, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Sun, Y.; Rong, X.; Bai, L. High glucose promotes breast cancer proliferation and metastasis by impairing angiotensinogen expression. Biosci. Rep. 2019, 39, BSR20190436. [Google Scholar] [CrossRef] [Green Version]
- Shomali, N.; Mahmoudi, J.; Mahmoodpoor, A.; Zamiri, R.E.; Akbari, M.; Xu, H.; Shotorbani, S.S. Harmful effects of high amounts of glucose on the immune system: An updated review. Biotechnol. Appl. Biochem. 2020, 68, 404–410. [Google Scholar] [CrossRef]
- Von Ah Morano, A.E.; Dorneles, G.P.; Peres, A.; Lira, F.S. The role of glucose homeostasis on immune function in response to exercise: The impact of low or higher energetic conditions. J. Cell. Physiol. 2020, 235, 3169–3188. [Google Scholar] [CrossRef]
- O’Mahony, F.; Raz, I.M.; Pedram, A.; Harvey, B.J.; Levin, E.R. Estrogen modulates metabolic pathway adaptation to available glucose in breast cancer cells. Mol. Endocrinol. 2012, 26, 2058–2070. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.S.; Zhao, Z.; Yang, Z.N.; Xu, F.; Lu, H.J.; Zhu, Z.Y.; Shi, W.; Jiang, J.; Yao, P.P.; Zhu, H.P. Risk factors and preventions of breast cancer. Int. J. Biol. Sci. 2017, 13, 1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, G.M.; Hausman, R.E.; Hausman, R.E. The Cell: A Molecular Approach; ASM Press: Washington, DC, USA, 2007; Volume 4, pp. 649–656. [Google Scholar]
- Murphy, K.M.; Weaver, C. Janeway’s Immunobiology: Ninth International Student Edition; Garland Science, Taylor & Francis Group, LLC: New York, NY, USA, 2017. [Google Scholar]
- Bronshtein, I.N.; Semendyayev, K.A. Handbook of Mathematics; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Tilekar, K.; Upadhyay, N.; Iancu, C.V.; Pokrovsky, V.; Choe, J.Y.; Ramaa, C.S. Power of two: Combination of therapeutic approaches involving glucose transporter (GLUT) inhibitors to combat cancer. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2020, 1874, 188457. [Google Scholar] [CrossRef] [PubMed]
- Pliszka, M.; Szablewski, L. Glucose transporters as a target for anticancer therapy. Cancers 2021, 13, 4184. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhu, J.; Yu, S.J.; Ma, H.L.; Chen, J.; Ding, X.F.; Chen, G.; Liang, Y.; Zhang, Q. Sodium-glucose co-transporter-2 (SGLT-2) inhibition reduces glucose uptake to induce breast cancer cell growth arrest through AMPK/mTOR pathway. Biomed. Pharmacother. 2020, 132, 110821. [Google Scholar] [CrossRef] [PubMed]
- Stefano, B.; Desmarchelier, D. Local bifurcations of three and four-dimensional systems: A tractable characterization with economic applications. Math. Soc. Sci. 2019, 97, 38–50. [Google Scholar]
- Cardin, P.T.; Llibre, J. Transcritical and zero-Hopf bifurcations in the Genesio system. Nonlinear Dyn. 2017, 88, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Weber, A. Deciding Hopf bifurcations by quantifier elimination in a software-component architecture. J. Symb. 2000, 30, 161–179. [Google Scholar]
- Hong, H.; Liska, R.; Steinberg, S. Testing stability by quantifier elimination. J. Symb. Comput. 1997, 24, 161–187. [Google Scholar] [CrossRef]
- Hatami, M.; Akbari, M.E.; Abdollahi, M.; Ajami, M.; Jamshidinaeini, Y.; Davoodi, S.H. The relationship between intake of macronutrients and vitamins involved in one carbon metabolism with breast cancer risk. Sci. Inf. Database 2017, 75, 56–64. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Unit |
|---|---|---|
| 0.7 | day | |
| 0.8 | day | |
| 0.1 | day | |
| 0.98 | day | |
| 0.4 | day | |
| 0.8 | day | |
| 0.3 | day | |
| 0.3 | day | |
| 0.29 | day | |
| 0.15 | day |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alblowy, A.H.; Maan, N.; Alharbi, S.A. Role of Glucose Risk Factors on Human Breast Cancer: A Nonlinear Dynamical Model Evaluation. Mathematics 2022, 10, 3640. https://doi.org/10.3390/math10193640
Alblowy AH, Maan N, Alharbi SA. Role of Glucose Risk Factors on Human Breast Cancer: A Nonlinear Dynamical Model Evaluation. Mathematics. 2022; 10(19):3640. https://doi.org/10.3390/math10193640
Chicago/Turabian StyleAlblowy, Abeer Hamdan, Normah Maan, and Sana Abdulkream Alharbi. 2022. "Role of Glucose Risk Factors on Human Breast Cancer: A Nonlinear Dynamical Model Evaluation" Mathematics 10, no. 19: 3640. https://doi.org/10.3390/math10193640