
Bistability and Robustness for Virus Infection Models with Nonmonotonic Immune Responses in Viral Infection Systems


Abstract
:1. Introduction
2. Bifurcations Analysis
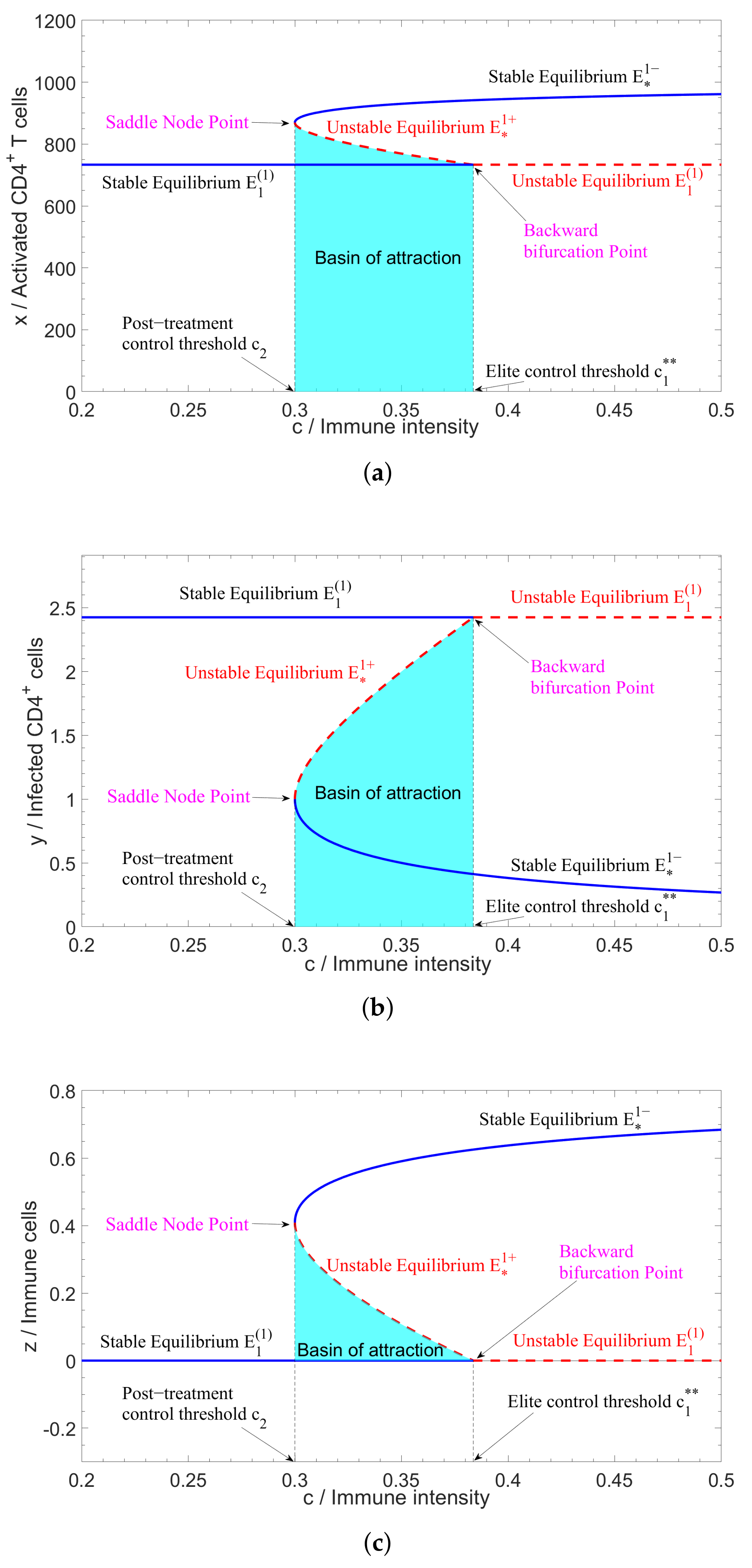
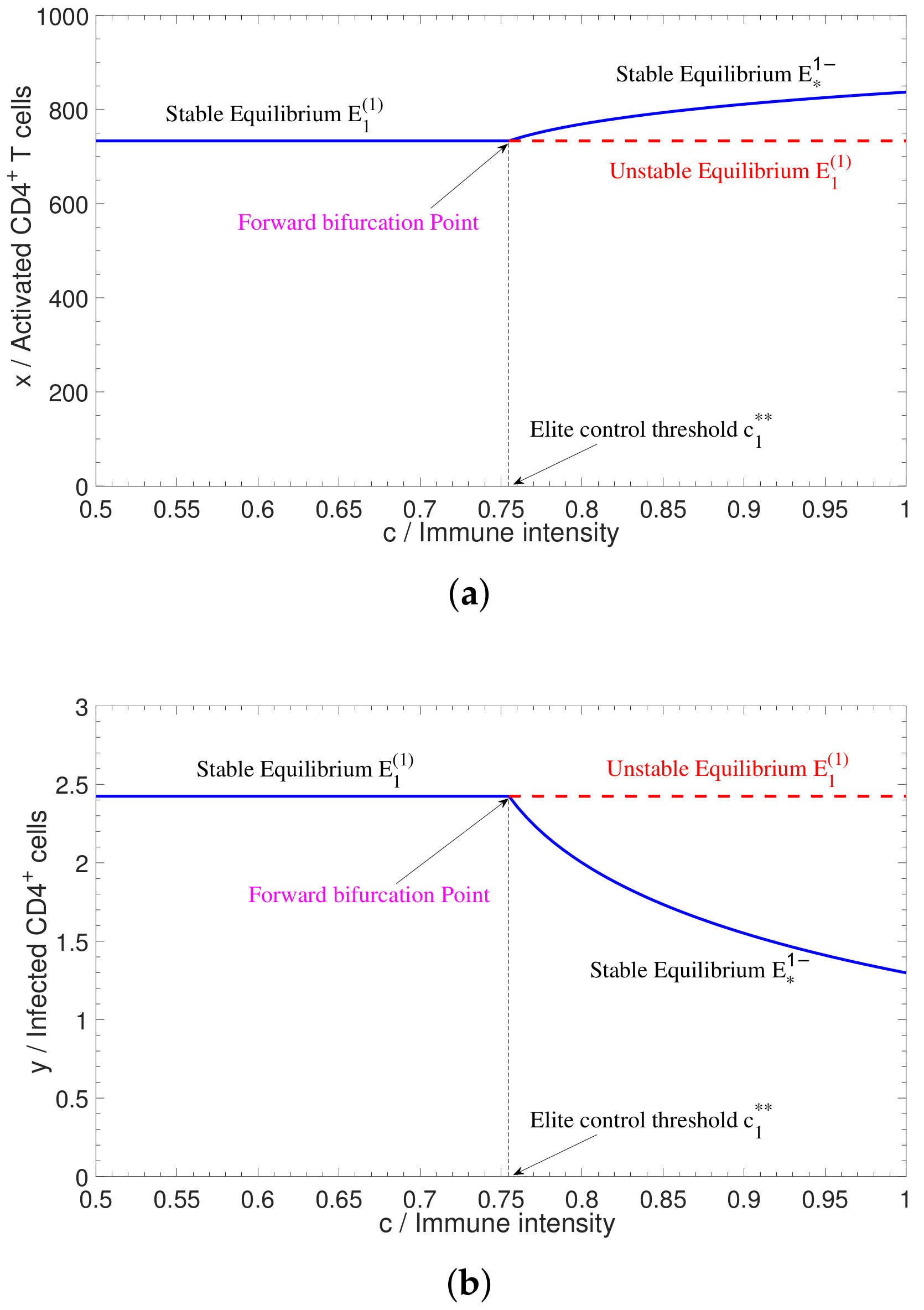
2.1. Bifurcations in Three-Dimensional Model
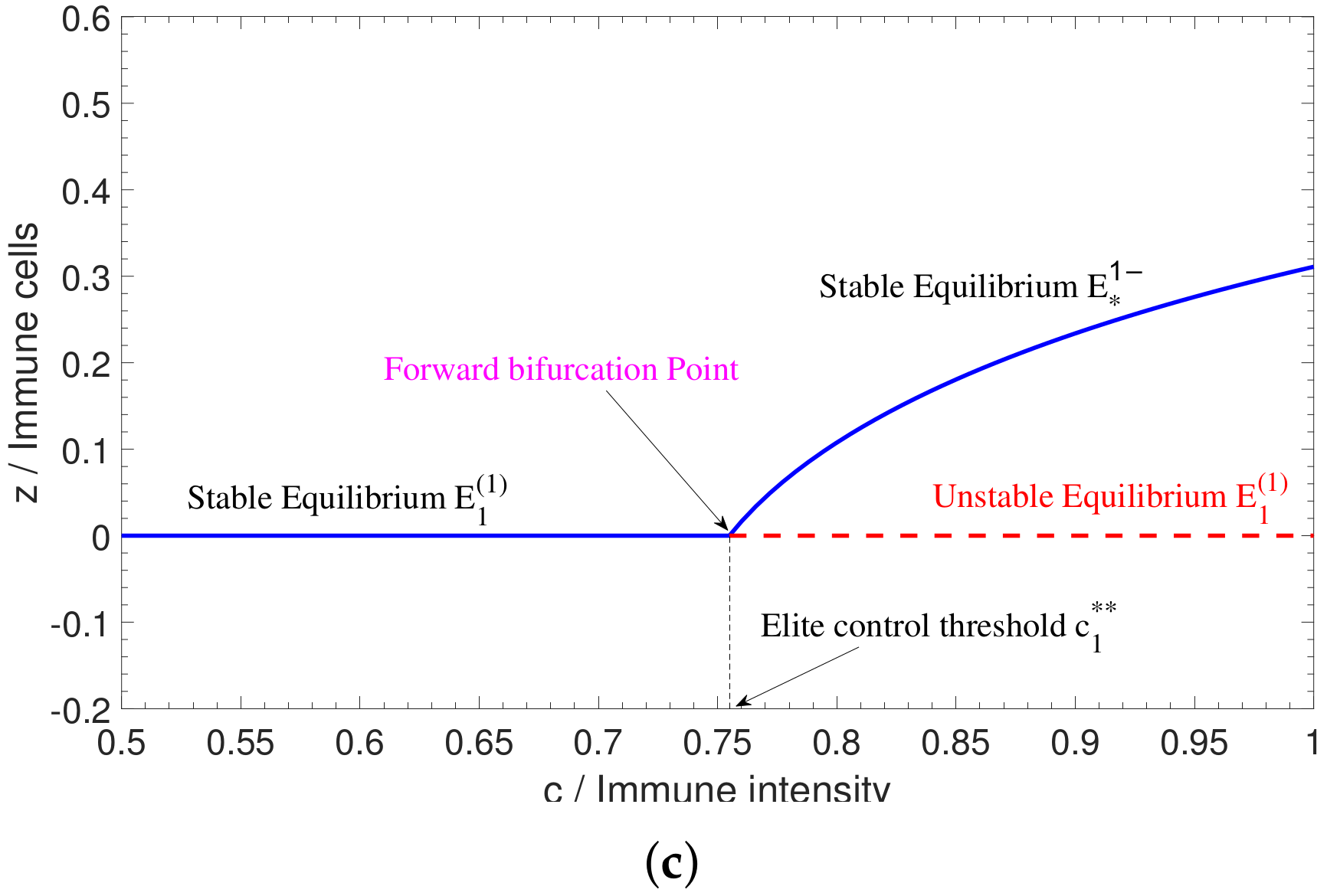
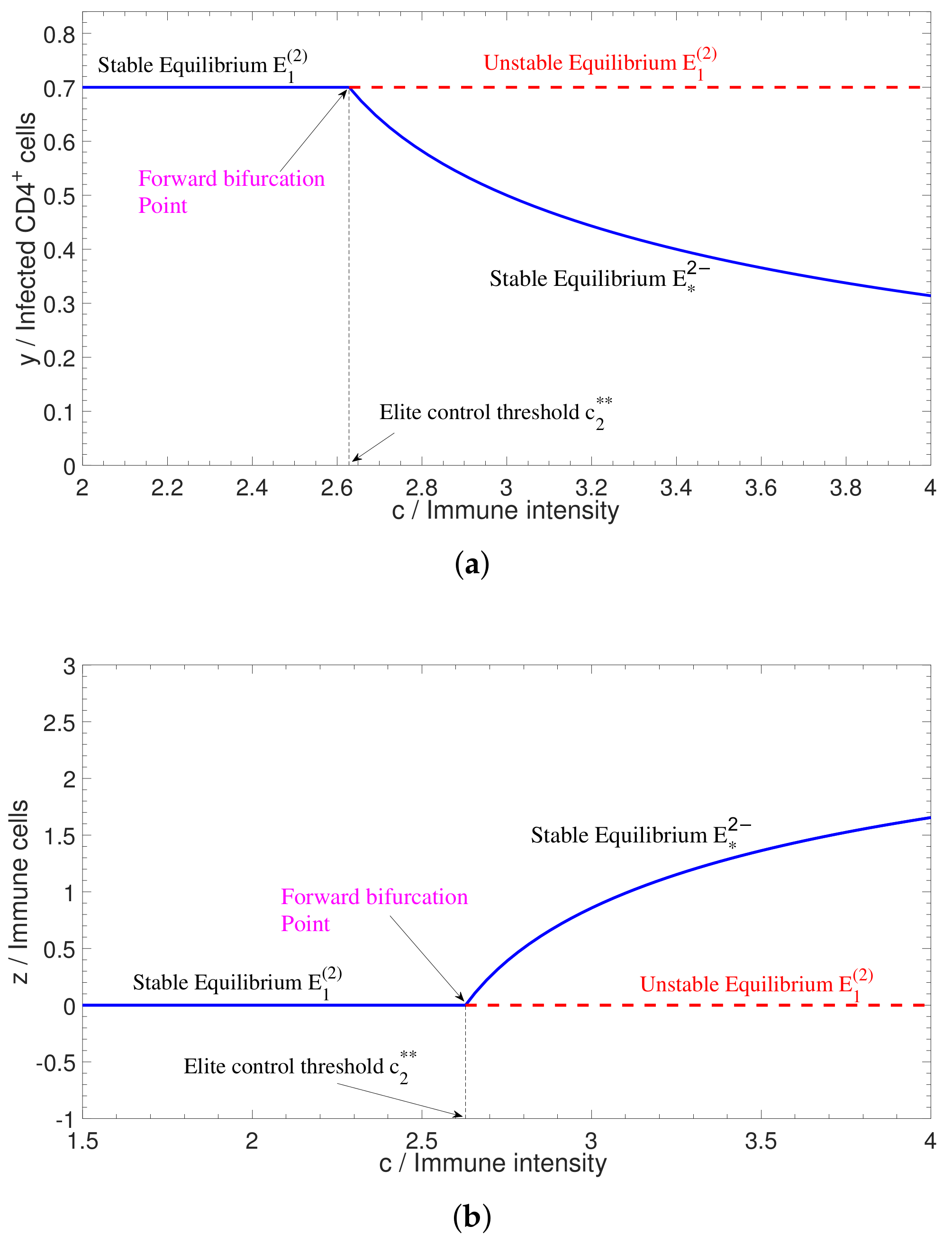
2.2. Bifurcations in Two-Dimensional Model
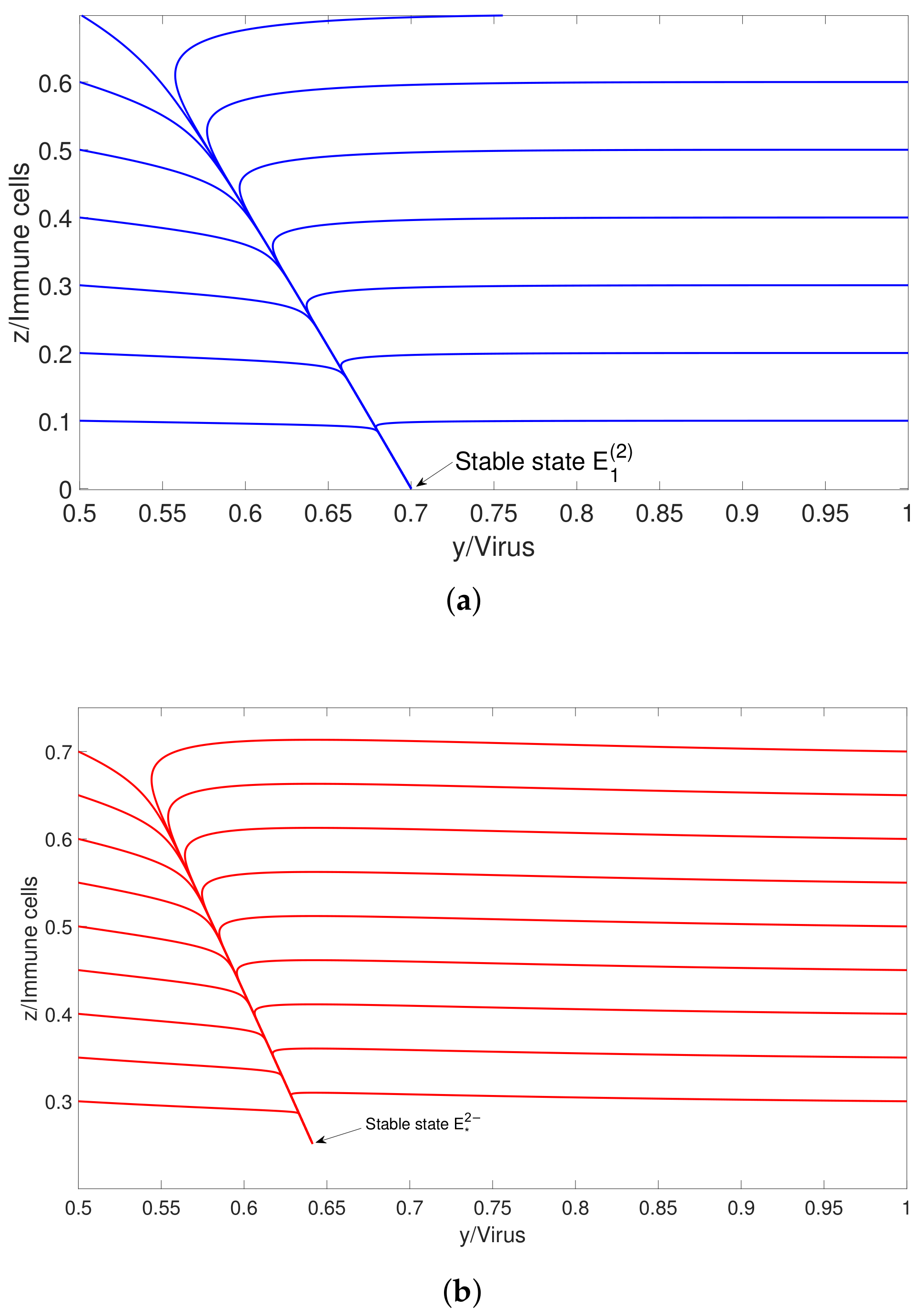
3. Robustness for Bistable System
4. Sensitive Analysis and Numerical Simulations
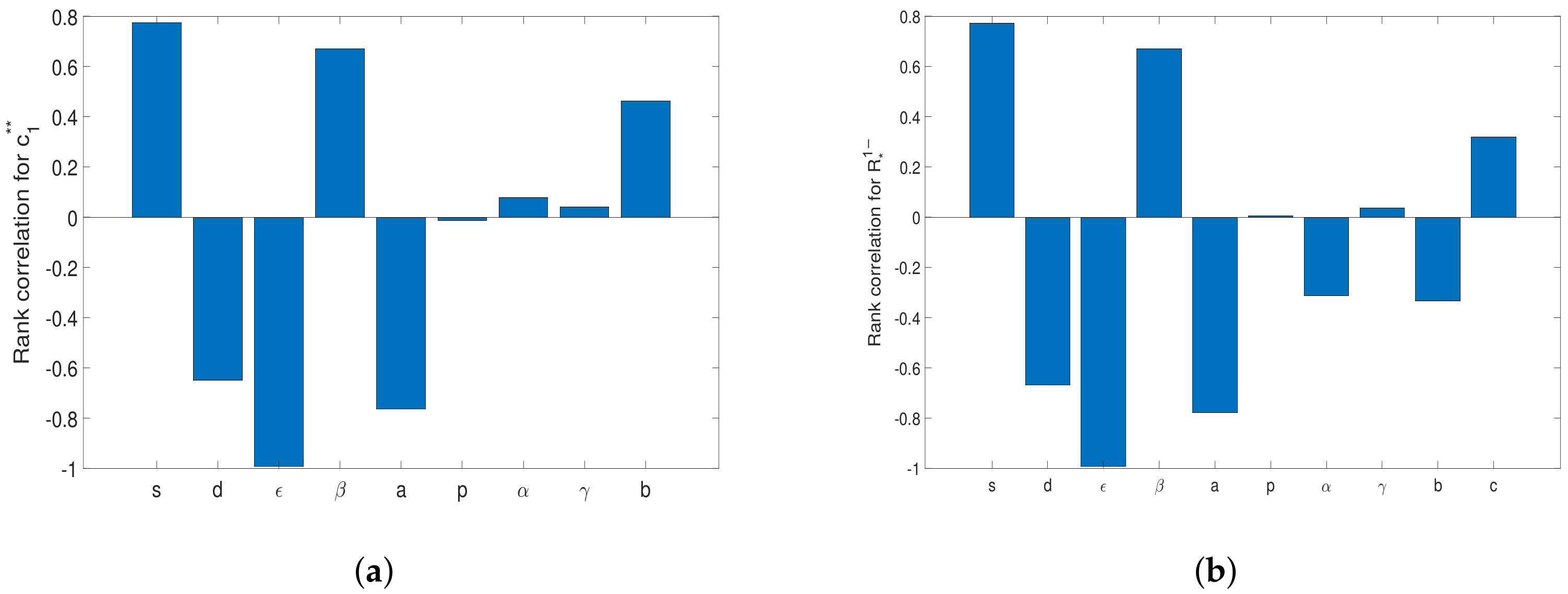
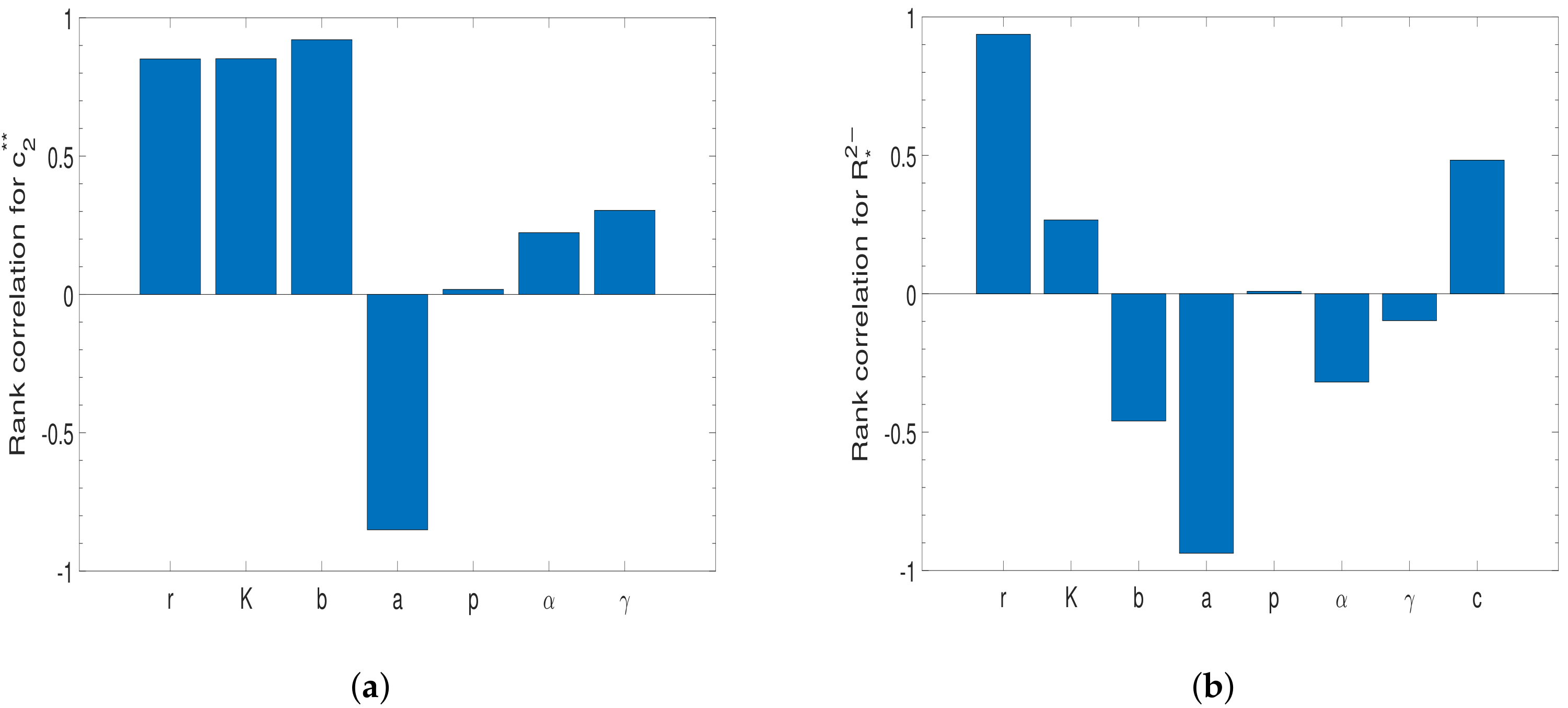
4.1. Sensitive Analysis
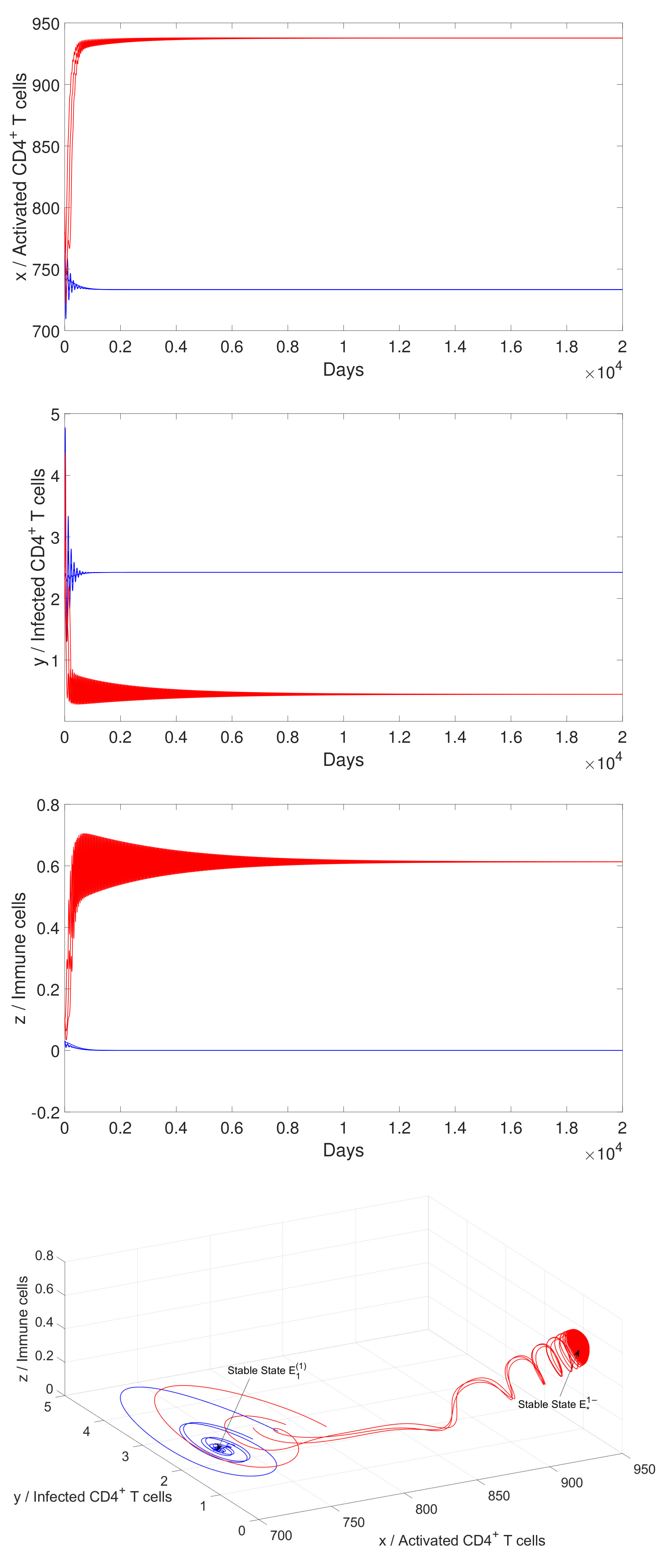
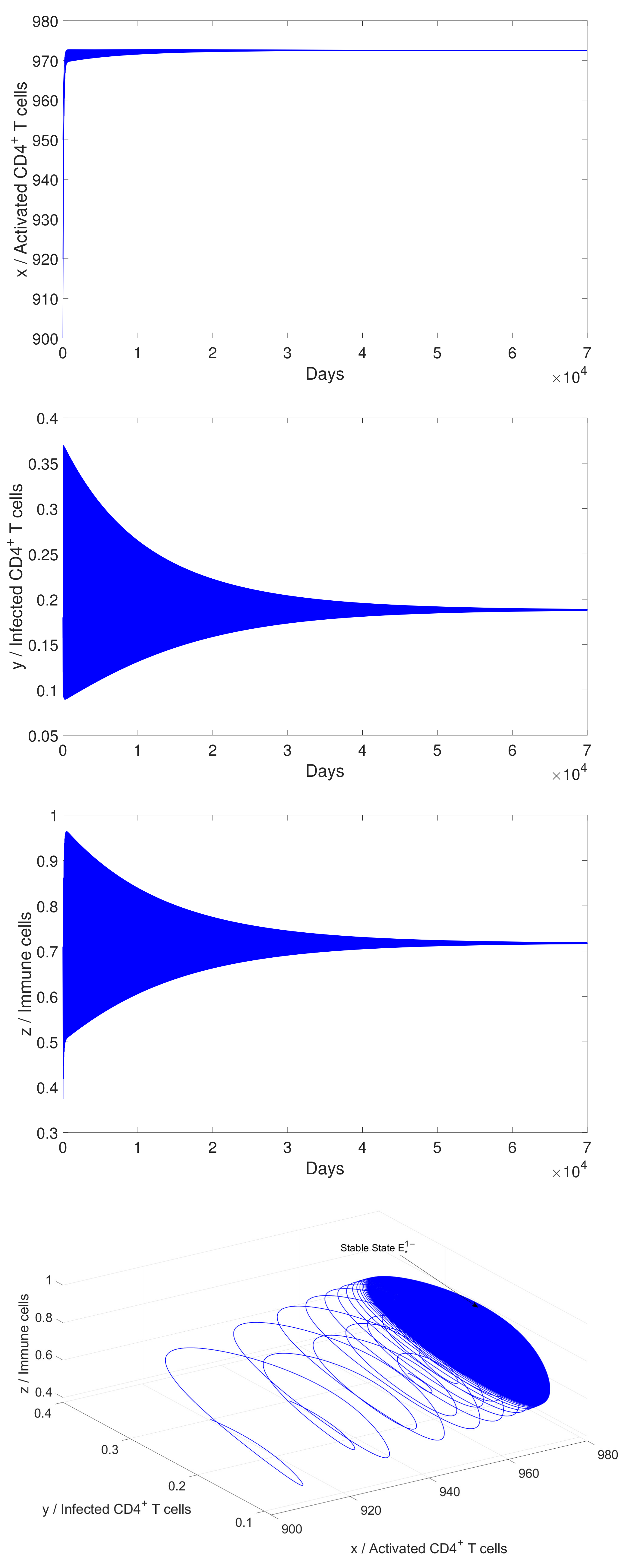
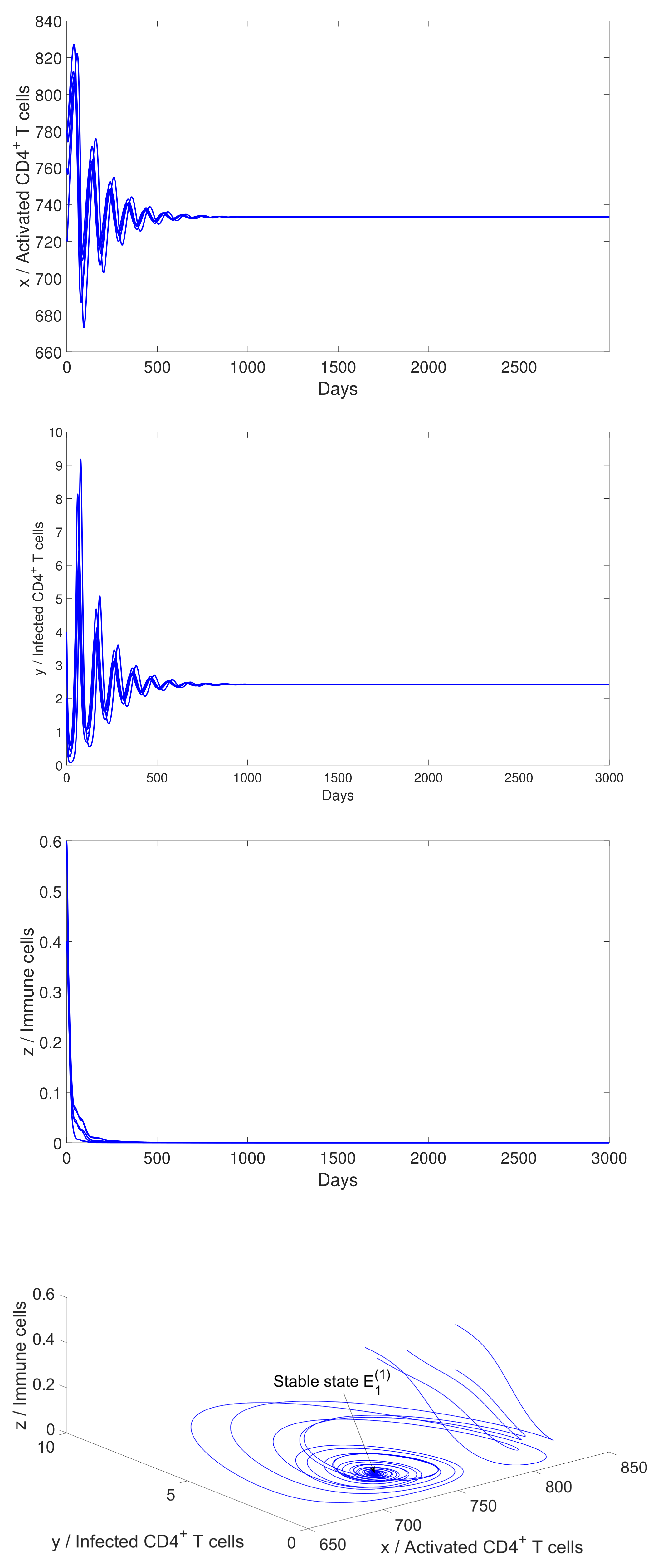
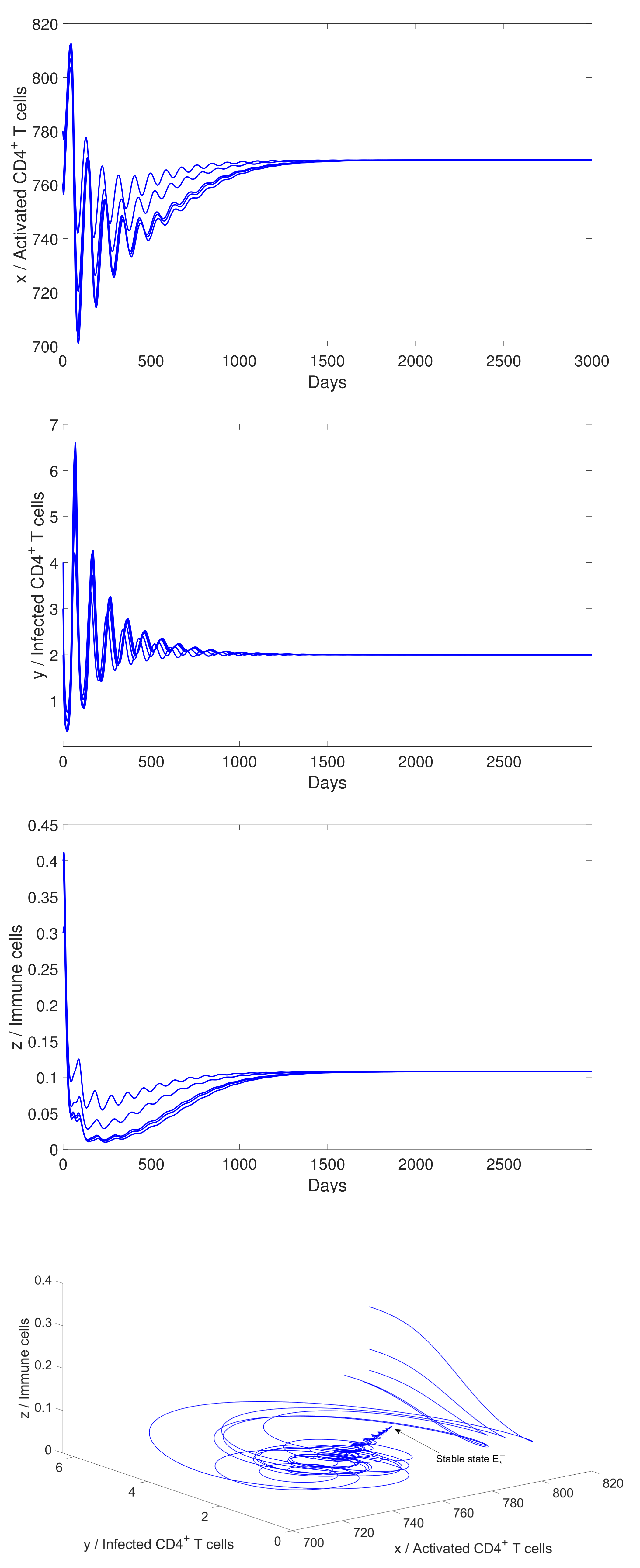
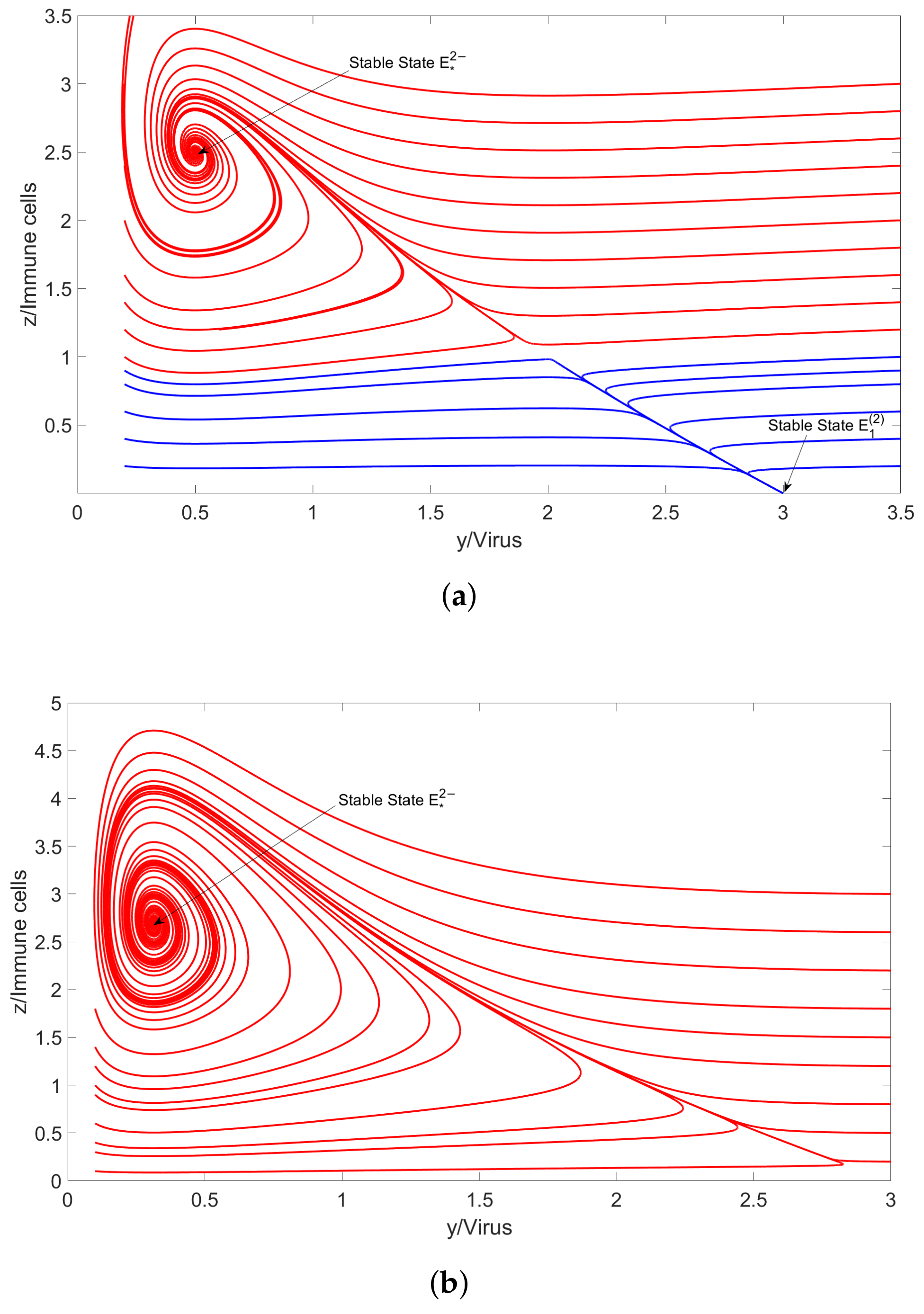
4.2. Numerical Simulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Description | Value | Reference |
|---|---|---|---|
| s | Production rate of CD T cells | 10 cells/μL/day | [29] |
| d | Death rate of CD T cells | [29] | |
| Drug efficacy | 0.9 | – | |
| Infection rate of CD T cells | 0.015 cells/μL/day | [6] | |
| a | Death rate of infected cells | – | |
| p | Killing rate of infected CD T cells by immune cells | – | |
| c | immune intensity | – | |
| Constant in nonmonotonic immune response | 1 cells/μL | – | |
| Constant in nonmonotonic immune response | 1 cells/μL | – | |
| b | Death rate of immune cells | – |
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, S.; Li, H.; Xu, F. Monotomic and nonmonotonic immune responses in viral infection systems. Discrete Cont. Dyn-B 2022, 27, 141–165. [Google Scholar] [CrossRef]
- Wang, S.; Xu, F. Thresholds and bistability in virus-immune dynamics. Appl. Math. Lett. 2018, 78, 105–111. [Google Scholar] [CrossRef]
- Wang, S.; Xu, F.; Rong, L. Bistability analysis of an hiv model with immune response. J. Biol. Syst. 2017, 25, 677–695. [Google Scholar] [CrossRef]
- Wang, S.; Xu, F. Analysis of an HIV model with post-treatment control. J. Appl. Anal. Comput. 2020, 10, 667–685. [Google Scholar] [CrossRef]
- Wang, X.; Tan, Y.; Cai, Y.; Wang, K.; Wang, W. Dynamics of a stochastic HBV infection model with cell-to-cell transmission and immune response. Math. Biosci. Eng. 2020, 18, 616–642. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.; Perelson, A. Post-treatment control of HIV infection. Proc. Natl. Acad. Sci. USA 2015, 112, 5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Tan, Y.; Cai, Y.; Wang, K.; Wang, W. Dynamics of a delayed integro-differential HIV infection model with multiple target cells and nonlocal dispersal. Eur. Phys. J. Plus 2021, 136, 117. [Google Scholar]
- Bi, K.; Chen, Y.; Zhao, S.; Ben-Arieh, D.; Wu, C.H.J. A new zoonotic visceral leishmaniasis dynamic transmission model with age-structure. Chaos Solitons Fractals 2020, 133, 109622. [Google Scholar] [CrossRef]
- AlAgha, A.; Elaiw, A. Stability of a general reaction-diffusion HIV-1 dynamics model with humoral immunity. Eur. Phys. J. Plus 2019, 134, 390. [Google Scholar] [CrossRef]
- Wang, W.; Ma, W.; Feng, Z. Dynamics of reaction–diffusion equations for modeling CD4+ T cells decline with general infection mechanism and distinct dispersal rates. Nonlinear Anal. RWA 2020, 51, 102976. [Google Scholar] [CrossRef]
- Ruan, S.; Xiao, D. Global analysis in a predator-prey system with nonmonotonic function response. SIAM J. Appl. Math. 2000, 61, 1445–1472. [Google Scholar]
- Wang, S.; Wang, X.; Wu, X. Bifurcation analysis for a food chain model with nonmonotonic nutrition conversion rate of predator to top predator. Int. J. Bifurcat. Chaos 2020, 30, 2050113. [Google Scholar] [CrossRef]
- Lu, M.; Huang, J.; Ruan, S.; Yu, P. Bifurcation analysis of an SIRS epidemic model with a generalized nonmonotone and saturated incidence rate. J. Differ. Equ. 2019, 267, 1859–1898. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Huang, J.; Ruan, S.; Yu, P. Global dynamics of a susceptible-infectious-recovered epidemic model with a generalized nonmonotone incidence rate. J. Dyn. Differ. Equ. 2020, 33, 1625. [Google Scholar] [CrossRef]
- Andrews, J.F. A mathematical model for the continuous culture of microorganisms utilizing inhibitory substrates. Biotechnol. Bioeng. 1968, 10, 707–723. [Google Scholar] [CrossRef]
- Xie, X.; Ma, J.; van den Driessche, P. Backward bifurcation in within-host HIV models. Math. Biosci. 2021, 335, 108569. [Google Scholar] [CrossRef]
- van den Driessche, P.; Watmough, J. A simple SIS epidemic model with a backward bifurcation. J. Math. Biol. 2000, 40, 525–540. [Google Scholar] [CrossRef]
- Bi, K.; Chen, Y.; Wu, C.H.J.; Ben-Arieh, D. A memetic algorithm for solving optimal control problems of Zika virus epidemic with equilibriums and backward bifurcation analysis. Commun. Nonlinear Sci. Numer. Simul. 2020, 84, 105176. [Google Scholar] [CrossRef]
- Chen, Y.; Bi, K.; Zhao, S.; Ben-Arieh, D.; Wu, C.H.J. Modeling individual fear factor with optimal control in a disease-dynamic system. Chaos Solitons Fractals 2017, 104, 531–545. [Google Scholar] [CrossRef]
- Zhao, S.; Kuang, Y.; Wu, C.H.; Ben-Arieh, D.; Ramalho-Ortigao, M.; Bi, K. Zoonotic visceral leishmaniasis transmission: Modeling, backward bifurcation, and optimal control. J. Math. Biol. 2016, 73, 1525–1560. [Google Scholar] [CrossRef]
- Bi, K.; Chen, Y.; Wu, C.H.J.; Ben-Arieh, D. Learning-based impulse control with event-triggered conditions for an epidemic dynamic system. Commun. Nonlinear Sci. Numer. Simul. 2022, 108, 106204. [Google Scholar] [CrossRef]
- Garba, S.M.; Gumel, A.B.; Bakar, M.A. Backward bifurcations in dengue transmission dynamics. Math. Biosci. 2008, 215, 11–25. [Google Scholar] [CrossRef] [PubMed]
- van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef]
- Castillo-Chavez, C.; Song, B. Dynamical models of tuberculosis and their applications. Math. Biosci. Eng. 2004, 1, 361–404. [Google Scholar] [CrossRef]
- Kitano, H. Towards a theory of biological robustness. Mol. Syst. Biol. 2007, 3, 137. [Google Scholar] [CrossRef]
- Xiao, Y.; Tang, S.; Zhou, Y.; Smith, R.; Wu, J.; Wang, N. Predicting the HIV/AIDS epidemic and measuring the effect of mobility in mainland China. J. Theor. Biol. 2013, 317, 271–285. [Google Scholar] [CrossRef]
- Blower, S.; Dowlatabadi, H. Sensitivity and uncertainty analysis of complex models of disease transmission: An HIV model, as an example. Int. Stat. Rev. 1994, 2, 229–243. [Google Scholar] [CrossRef]
- Marino, S.; Hogue, I.; Ray, C.; Kirschner, D. A methodology for performing global uncertainty and sensitivity analysis in systems biology. J. Theor. Biol. 2008, 254, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Bonhoeffer, S.; Rembiszewski, M.; Ortiz, G.; Nixon, D. Risks and benefits of structured antiretroviral drug therapy interruptions in HIV-1 infection. AIDS 2000, 14, 2313–2322. [Google Scholar] [CrossRef]













| System (1) | ||||
|---|---|---|---|---|
| LAS | — | — | Converges to | |
| US | LAS | — | Converges to | |
| , | LAS | — | — | Converges to |
| LAS | LAS | US | Bistable | |
| US | LAS | — | Converges to |
| System (2) | ||||
|---|---|---|---|---|
| GAS | — | — | Converges to | |
| US | GAS | — | Converges to | |
| , | GAS | — | — | Converges to |
| GAS | GAS | US | Bistable | |
| US | GAS | — | Converges to |
| Symbol | Description | Value | Reference |
|---|---|---|---|
| r | Growth rate of infected cells | – | |
| K | Environmental carrying capacity of infected cells | – | – |
| a | Death rate of cells | – | |
| p | Killing rate of infected CD T cells by immune cells | – | |
| c | immune intensity | – | |
| Constant in nonmonotonic immune response | 1 cells/μL | – | |
| Constant in nonmonotonic immune response | 1 cells/μL | – | |
| b | Death rate of immune cells | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Wang, S.; Xu, F. Bistability and Robustness for Virus Infection Models with Nonmonotonic Immune Responses in Viral Infection Systems. Mathematics 2022, 10, 2139. https://doi.org/10.3390/math10122139
Wang T, Wang S, Xu F. Bistability and Robustness for Virus Infection Models with Nonmonotonic Immune Responses in Viral Infection Systems. Mathematics. 2022; 10(12):2139. https://doi.org/10.3390/math10122139
Chicago/Turabian StyleWang, Tengfei, Shaoli Wang, and Fei Xu. 2022. "Bistability and Robustness for Virus Infection Models with Nonmonotonic Immune Responses in Viral Infection Systems" Mathematics 10, no. 12: 2139. https://doi.org/10.3390/math10122139