
A Novel Urinary Proteomics Classifier for Non-Invasive Evaluation of Interstitial Fibrosis and Tubular Atrophy in Chronic Kidney Disease

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Cohort
2.2. CE-MS Analysis
2.3. Sequencing of Peptides
2.4. Protease Prediction
2.5. Statistical Methods
3. Results
3.1. Patient Characteristics
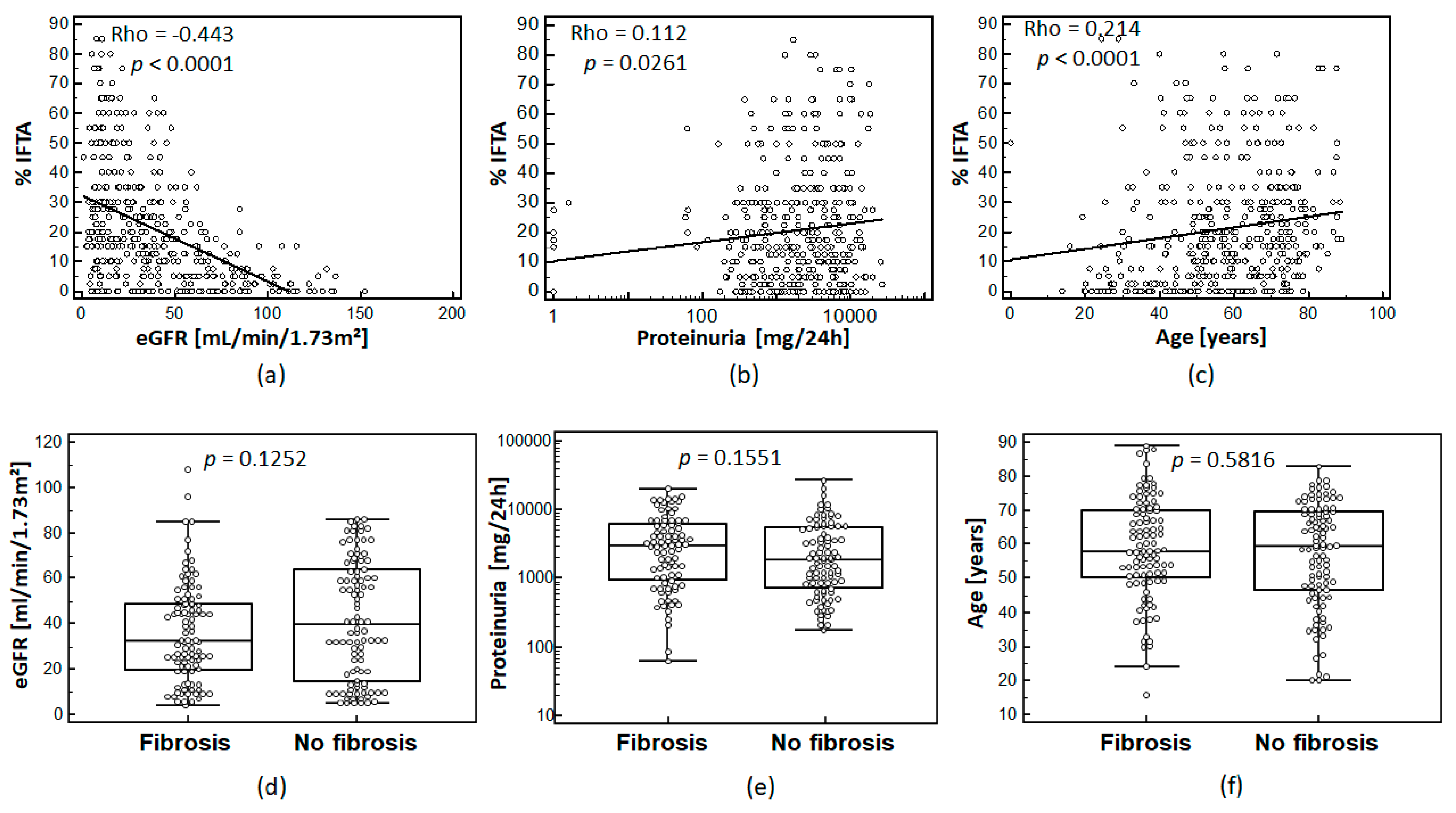
3.2. Relationship between IFTA and Clinical Parameters
3.3. Definition of Urinary Peptides Associated with IFTA
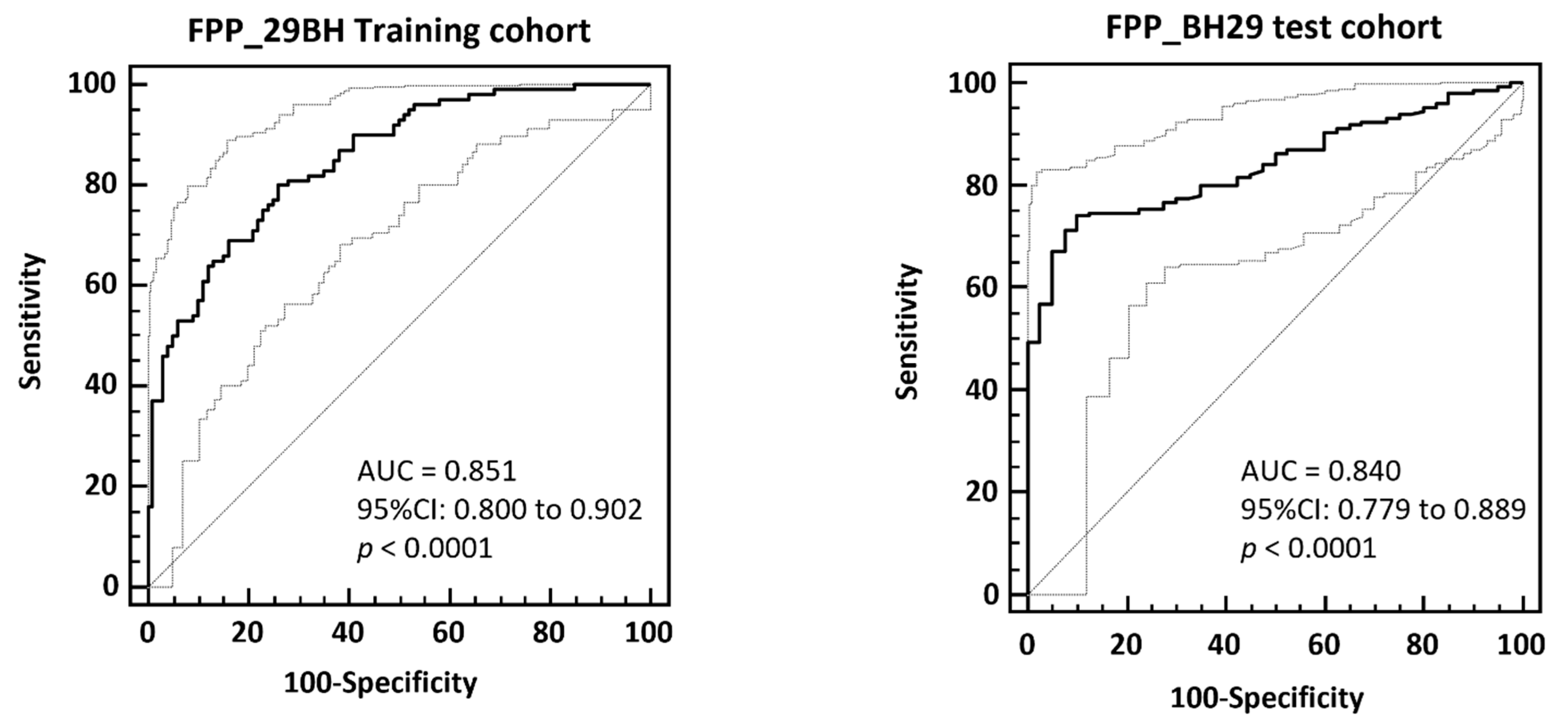
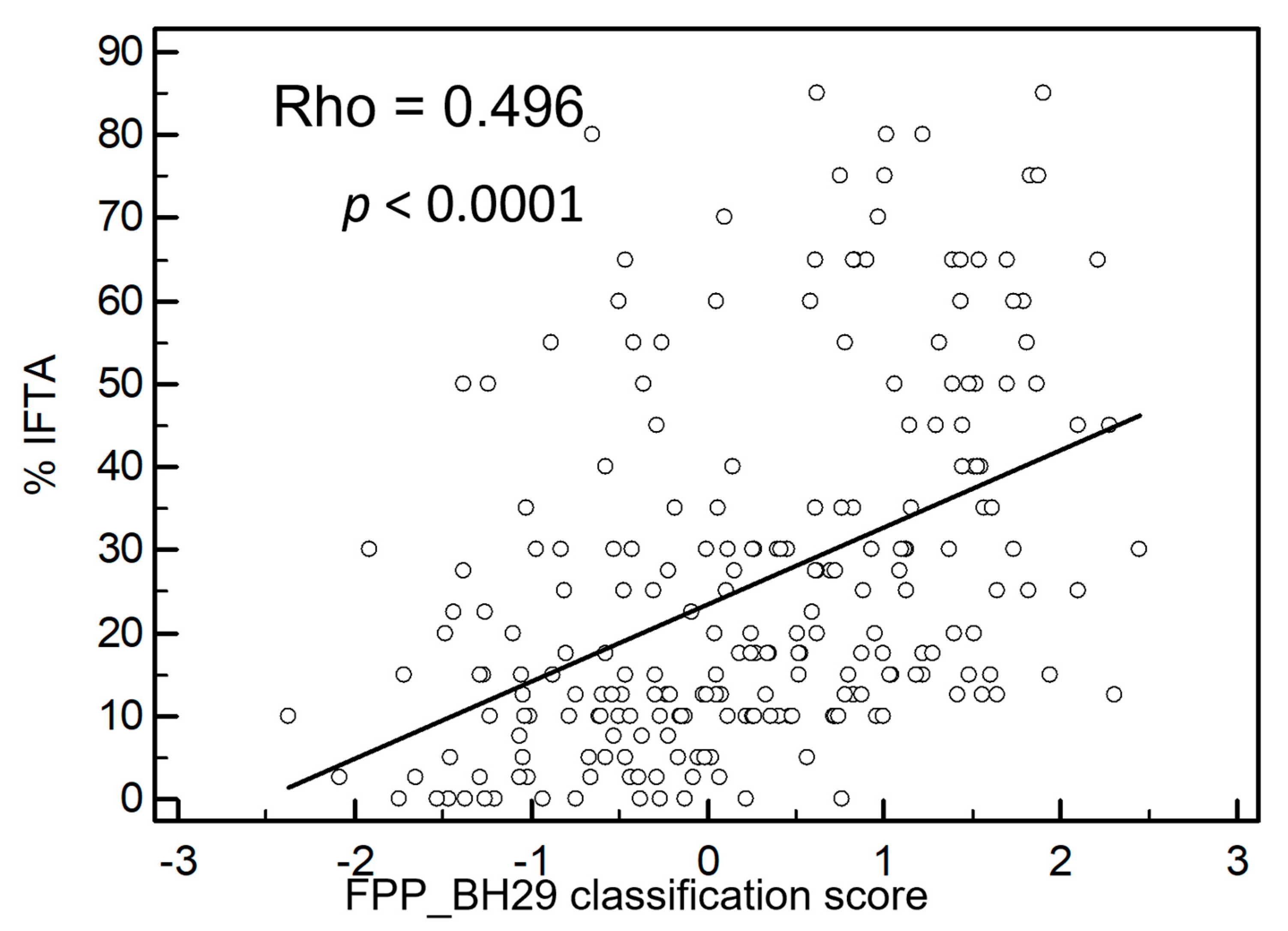
3.4. Validation of the FPP_29BH Classifier
3.5. Prediction of Proteases
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Latosinska, A.; Siwy, J.; Mischak, H.; Frantzi, M. Peptidomics and proteomics based on CE-MS as a robust tool in clinical application: The past, the present, and the future. Electrophoresis 2019, 40, 2294–2308. [Google Scholar] [CrossRef]
- Latosinska, A.; Siwy, J.; Faguer, S.; Beige, J.; Mischak, H.; Schanstra, J.P. Value of Urine Peptides in Assessing Kidney and Cardiovascular Disease. Proteom. Clin. Appl. 2021, 15, e2000027. [Google Scholar] [CrossRef]
- Magalhães, P.M.; Pejchinovski, K.; Markoska, M.; Banasik, M.; Klinger, D.; Svec-Billa, I.; Rychlik, M.; Rroji, A.; Restivo, G.; Capasso, F.; et al. Association of kidney fibrosis with urinary peptides: A path towards non-invasive liquid biopsies? Sci. Rep. 2017, 7, 16915. [Google Scholar] [CrossRef] [Green Version]
- Good, D.M.; Zurbig, P.; Argiles, A.; Bauer, H.W.; Behrens, G.; Coon, J.J.; Dakna, M.; Decramer, S.; Delles, C.; Dominiczak, A.F.; et al. Naturally occurring human urinary peptides for use in diagnosis of chronic kidney disease. Mol. Cell. Proteom. 2010, 9, 2424–2437. [Google Scholar] [CrossRef] [Green Version]
- Schanstra, J.P.; Zurbig, P.; Alkhalaf, A.; Argiles, A.; Bakker, S.J.; Beige, J.; Bilo, H.J.; Chatzikyrkou, C.; Dakna, M.; Dawson, J.; et al. Diagnosis and Prediction of CKD Progression by Assessment of Urinary Peptides. J. Am. Soc. Nephrol. 2015, 26, 1999–2010. [Google Scholar] [CrossRef]
- Siwy, J.; Zurbig, P.; Argiles, A.; Beige, J.; Haubitz, M.; Jankowski, J.; Julian, B.A.; Linde, P.G.; Marx, D.; Mischak, H.; et al. Noninvasive diagnosis of chronic kidney diseases using urinary proteome analysis. Nephrol. Dial. Transpl. 2017, 32, 2079–2089. [Google Scholar] [CrossRef]
- Farris, A.B.; Alpers, C.E. What is the best way to measure renal fibrosis: A pathologist’s perspective. Kidney Int. Suppl. 2014, 4, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikbov, B.; Purcell, C.A.; Levey, A.S.; Smith, M.; Abdoli, A.; Abebe, M.; Adebayo, O.M.; Afarideh, M.; Agarwal, S.K.; Agudelo-Botero, M.; et al. Global, regional, and national burden of chronic kidney disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D. Global Prevalence of Chronic Kidney Disease—A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef] [PubMed]
- Kidney Disease: Improving Global Outcomes (KDIGO) CKD Work Group. KDIGO 2012 Clinical Practice Guideline for the Evaluation and Management of Chronic Kidney Disease. Kidney Int. 2013, 3, 1–150. [Google Scholar]
- Stevens, P.E.; Levin, A.; Kidney, M.; Disease: Improving Global Outcomes Chronic Kidney Disease Guideline Development Work Group. Evaluation and management of chronic kidney disease: Synopsis of the kidney disease: Improving global outcomes 2012 clinical practice guideline. Ann. Intern. Med. 2013, 158, 825–830. [Google Scholar] [CrossRef] [Green Version]
- Farris, A.B.; Colvin, R.B. Renal interstitial fibrosis: Mechanisms and evaluation. Curr. Opin. Nephrol. Hypertens. 2012, 21, 289–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nangaku, M. Chronic hypoxia and tubulointerstitial injury: A final common pathway to end-stage renal failure. J. Am. Soc. Nephrol. 2006, 17, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, C.H.; Le, W.; Ni, Z.; Zhang, M.; Miao, L.; Luo, P.; Wang, R.; Lv, Z.; Chen, J.; Tian, J.; et al. A multicenter application and evaluation of the oxford classification of IgA nephropathy in adult chinese patients. Am. J. Kidney. Dis. 2012, 60, 812–820. [Google Scholar] [CrossRef]
- Solez, K.; Colvin, R.B.; Racusen, L.C.; Haas, M.; Sis, B.; Mengel, M.; Halloran, P.F.; Baldwin, W.; Banfi, G.; Collins, A.B.; et al. Banff 07 classification of renal allograft pathology: Updates and future directions. Am. J. Transpl. 2008, 8, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.S.; Stevens, L.A.; Schmid, C.H.; Zhang, Y.; Castro, A.F., III; Feldman, H.I.; Kusek, J.W.; Eggers, P.; Van Lente, F.; Greene, T.; et al. A new equation to estimate glomerular filtration rate. Ann. Intern. Med. 2009, 150, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Mischak, H.; Vlahou, A.; Ioannidis, J.P. Technical aspects and inter-laboratory variability in native peptide profiling: The CE-MS experience. Clin. Biochem. 2013, 46, 432–443. [Google Scholar] [CrossRef]
- Tofte, N.; Lindhardt, M.; Adamova, K.; Bakker, S.J.L.; Beige, J.; Beulens, J.W.J.; Birkenfeld, A.L.; Currie, G.; Delles, C.; Dimos, I.; et al. Early detection of diabetic kidney disease by urinary proteomics and subsequent intervention with spironolactone to delay progression (PRIORITY): A prospective observational study and embedded randomised placebo-controlled trial. Lancet Diabetes Endocrinol. 2020, 8, 301–312. [Google Scholar] [CrossRef]
- Jantos-Siwy, J.; Schiffer, E.; Brand, K.; Schumann, G.; Rossing, K.; Delles, C.; Mischak, H.; Metzger, J. Quantitative urinary proteome analysis for biomarker evaluation in chronic kidney disease. J. Proteome Res. 2009, 8, 268–281. [Google Scholar] [CrossRef]
- Klein, J.; Papadopoulos, T.; Mischak, H.; Mullen, W. Comparison of CE-MS/MS and LC-MS/MS sequencing demonstrates significant complementarity in natural peptide identification in human urine. Electrophoresis 2014, 35, 1060–1064. [Google Scholar] [CrossRef]
- Zurbig, P.; Renfrow, M.B.; Schiffer, E.; Novak, J.; Walden, M.; Wittke, S.; Just, I.; Pelzing, M.; Neususs, C.; Theodorescu, D.; et al. Biomarker discovery by CE-MS enables sequence analysis via MS/MS with platform-independent separation. Electrophoresis 2006, 27, 2111–2125. [Google Scholar] [CrossRef]
- Klein, J.; Eales, J.; Zürbig, P.; Vlahou, A.; Mischak, H.; Stevens, R. Proteasix: A tool for automated and large-scale prediction of proteases involved in naturally occurring peptide generation. Proteomics 2013, 13, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Dakna, M.; Harris, K.; Kalousis, A.; Carpentier, S.; Kolch, W.; Schanstra, J.P.; Haubitz, M.; Vlahou, A.; Mischak, H.; Girolami, M. Addressing the Challenge of Defining Valid Proteomic Biomarkers and Classifiers. BMC Bioinform. 2010, 11, 594. [Google Scholar] [CrossRef] [Green Version]
- Metz, C.E. Basic principles of ROC analysis. Semin. Nucl. Med. 1978, 8, 283–298. [Google Scholar] [CrossRef]
- Zweig, M.H.; Campbell, G. Receiver-operating characteristic (ROC) plots: A fundamental evaluation tool in clinical medicine. Clin. Chem. 1993, 39, 561–577. [Google Scholar] [CrossRef] [PubMed]
- Boor, P.; Ostendorf, T.; Floege, J. Renal fibrosis: Novel insights into mechanisms and therapeutic targets. Nat. Rev. Nephrol. 2010, 6, 643–656. [Google Scholar] [CrossRef]
- Zürbig, P.; Mischak, H.; Menne, J.; Haller, H. CKD273 Enables Efficient Prediction of Diabetic Nephropathy in Nonalbuminuric Patients. Diabetes Care 2019, 42, e4–e5. [Google Scholar] [CrossRef] [Green Version]
- Critselis, E.; Vlahou, A.; Stel, V.S.; Morton, R.L. Cost-effectiveness of screening type 2 diabetes patients for chronic kidney disease progression with the CKD273 urinary peptide classifier as compared to urinary albumin excretion. Nephrol. Dial. Transpl. 2018, 33, 441–449. [Google Scholar] [CrossRef]
- Pontillo, C.; Zhang, Z.-Y.; Schanstra, J.P.; Jacobs, L.; Zürbig, P.; Thijs, L.; Ramírez-Torres, A.; Heerspink, H.J.; Lindhardt, M.; Klein, R.; et al. Prediction of Chronic Kidney Disease Stage 3 by CKD273, a Urinary Proteomic Biomarker. Kidney Int. Rep. 2017, 2, 1066–1075. [Google Scholar] [CrossRef]
- Pontillo, C.; Mischak, H. Urinary peptide-based classifier CKD273: Towards clinical application in chronic kidney disease. Clin. Kidney J. 2017, 10, 192–201. [Google Scholar] [CrossRef]
- Magalhães, P.; Zürbig, P.; Mischak, H.; Schleicher, E. Urinary fetuin-A peptides as a new marker for impaired kidney function in patients with type 2 diabetes. Clin. Kidney J. 2021, 14, 269–276. [Google Scholar] [CrossRef]
- Westenfeld, R.; Jahnen-Dechent, W.; Ketteler, M. Vascular Calcification and Fetuin-A Deficiency in Chronic Kidney Disease. Trends Cardiovasc. Med. 2007, 17, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Ketteler, M.; Bongartz, P.; Westenfeld, R.; Wildberger, J.E.; Mahnken, A.H.; Böhm, R.; Metzger, T.; Wanner, C.; Jahnen-Dechent, W.; Floege, J. Association of low fetuin-A (AHSG) concentrations in serum with cardiovascular mortality in patients on dialysis: A cross-sectional study. Lancet 2003, 361, 827–833. [Google Scholar] [CrossRef]
- Zhou, Z.; Ji, Y.; Ju, H.; Chen, H.; Sun, M. Circulating Fetuin-A and Risk of All-Cause Mortality in Patients With Chronic Kidney Disease: A Systematic Review and Meta-Analysis. Front. Physiol. 2019, 10, 966. [Google Scholar] [CrossRef]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, D.G.K.; Boesby, L.; Nielsen, S.H.; Tepel, M.; Birot, S.; Karsdal, M.A.; Kamper, A.L.; Genovese, F. Collagen turnover profiles in chronic kidney disease. Sci. Rep. 2019, 9, 16062. [Google Scholar] [CrossRef]
- Papasotiriou, M.; Genovese, F.; Klinkhammer, B.M.; Kunter, U.; Nielsen, S.H.; Karsdal, M.A.; Floege, J.; Boor, P. Serum and urine markers of collagen degradation reflect renal fibrosis in experimental kidney diseases. Nephrol. Dial. Transpl. 2015, 30, 1112–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoul, B.E.; Squalli, T.; Servais, A.; Elie, C.; Meas-Yedid, V.; Trivint, C.; Vanmassenhove, J.; Grunfeld, J.P.; Olivo-Marin, J.C.; Thervet, E.; et al. Urinary procollagen III aminoterminal propeptide (PIIINP): A fibrotest for the nephrologist. Clin. J. Am. Soc. Nephrol. 2010, 5, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Soylemezoglu, O.; Wild, G.; Dalley, A.J.; MacNeil, S.; Milford-Ward, A.; Brown, C.B.; el Nahas, A.M. Urinary and serum type III collagen: Markers of renal fibrosis. Nephrol. Dial. Transpl. 1997, 12, 1883–1889. [Google Scholar] [CrossRef]
- Wang, H.; Zheng, C.; Lu, Y.; Jiang, Q.; Yin, R.; Zhu, P.; Zhou, M.; Liu, Z. Urinary Fibrinogen as a Predictor of Progression of CKD. Clin. J. Am. Soc. Nephrol. 2017, 12, 1922–1929. [Google Scholar] [CrossRef]
- Goicoechea, M.; De Vinuesa, S.G.; Gómez-Campderá, F.; Aragoncillo, I.; Verdalles, U.; Mosse, A.; Luño, J. Serum fibrinogen levels are an independent predictor of mortality in patients with chronic kidney disease (CKD) stages 3 and 4. Kidney Int. 2008, 74, S67–S70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craciun, F.L.; Ajay, A.K.; Hoffmann, D.; Saikumar, J.; Fabian, S.L.; Bijol, V.; Humphreys, B.D.; Vaidya, V.S. Pharmacological and genetic depletion of fibrinogen protects from kidney fibrosis. Am. J. Physiol. Physiol. 2014, 307, F471–F484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Wang, F.; Kong, Y.; Wu, R.; Zhang, G.; Wang, N.; Wang, L.; Lu, Z.; Liang, M. Antithrombin III prevents progression of chronic kidney disease following experimental ischaemic-reperfusion injury. J. Cell. Mol. Med. 2017, 21, 3506–3514. [Google Scholar] [CrossRef] [PubMed]
- Ozden, A.; Sarioglu, A.; Demirkan, N.C.; Bilgihan, A.; Düzcan, E. Antithrombin III reduces renal ischemia-reperfusion injury in rats. Res. Exp. Med. (Berl.) 2001, 200, 195–203. [Google Scholar]
- Madhusudhan, T.; Kerlin, B.; Isermann, B. The emerging role of coagulation proteases in kidney disease. Nat. Rev. Nephrol. 2016, 12, 94–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawczyk, K.M.; Nilsson, H.; Nystrom, J.; Lindgren, D.; Leandersson, K.; Sward, K.; Johansson, M.E. Localization and Regulation of Polymeric Ig Receptor in Healthy and Diseased Human Kidney. Am. J. Pathol. 2019, 189, 1933–1944. [Google Scholar] [CrossRef]
- He, T.; Siwy, J.; Metzger, J.; Mullen, W.; Mischak, H.; Schanstra, J.P.; Zürbig, P.; Jankowski, V. Associations of urinary polymeric immunoglobulin receptor peptides in the context of cardio-renal syndrome. Sci. Rep. 2020, 10, 1–7. [Google Scholar] [CrossRef]
- Rudnicki, M.; Siwy, J.; Wendt, R.; Lipphardt, M.; Koziolek, M.J.; Maixnerova, D.; Peters, B.; Kerschbaum, J.; Leierer, J.; Neprasova, M.; et al. Urine proteomics for prediction of disease progression in patients with IgA nephropathy. Nephrol. Dial. Transpl. 2020, 35, gfaa140-MO041. [Google Scholar] [CrossRef]
- Moreno, J.A.; Sevillano, A.; Gutierrez, E.; Guerrero-Hue, M.; Vazquez-Carballo, C.; Yuste, C.; Herencia, C.; Garcia-Caballero, C.; Praga, M.; Egido, J. Glomerular Hematuria: Cause or Consequence of Renal Inflammation? Int. J. Mol. Sci. 2019, 20, 2205. [Google Scholar] [CrossRef] [Green Version]
- Coppo, R.; Fervenza, F.C. Persistent Microscopic Hematuria as a Risk Factor for Progression of IgA Nephropathy: New Floodlight on a Nearly Forgotten Biomarker. J. Am. Soc. Nephrol. 2017, 28, 2831–2834. [Google Scholar] [CrossRef] [Green Version]
- Fox, C.; Cocchiaro, P.; Oakley, F.; Howarth, R.; Callaghan, K.; Leslie, J.; Luli, S.; Wood, K.M.; Genovese, F.; Sheerin, N.S.; et al. Inhibition of lysosomal protease cathepsin D reduces renal fibrosis in murine chronic kidney disease. Sci. Rep. 2016, 6, 20101. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Study Cohort (n = 435) | Training Cohort | Test Cohort | p Test VS. Training | |
|---|---|---|---|---|
| Characteristics | Fibrosis | No Fibrosis | ||
| Number of subjects | 100 | 100 | 235 | |
| Age (years) * | 58.8 ± 14.9 | 56.9 ± 15.4 | 58.3 ± 17.5 | 0.793 |
| Gender (men/women) | 64/36 | 55/45 | 139/96 | n/a |
| eGFR (mL/min/1.73 m2, CKD-EPI) * | 35.9 ± 21.8 | 41.6 ± 26 | 37.7 ± 35.3 | 0.714 |
| Proteinuria (mg/24 h) * | 4274 ± 4415 | 3809 ± 4395 | 3785 ± 4074 | 0.814 |
| IFTA% * | 29.1 ± 13.9 | 2.6 ± 2.8 | 25.7 ± 21.1 | 1.267 × 10−7 |
| Urinary Peptides | Training Cohort 100/100 | Etiology Matched Cohort 55/55 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Gene Symbol | Sequence | adj. p-Value (BH) | Mean Intensity Fibrosis | Mean Intensity No Fibrosis | Fold Change | Unaj.Wilcox-p-Value | Mean Intensity Fibrosis | Mean No Fibrosis | Fold Change |
| COL10A1 | GHPGPSGPPGKpGYGSpGLQGEpGLPGPPGPS | 2.42 × 10−4 | 782.77 | 380.77 | 2.056 | 1.34 × 10−2 | 755.71 | 443.15 | 1.705 |
| COL1A2 | GPQGVQGGKGEQGPPGPPGFQGLPGPSGpAGEVGKpGERG | 2.42 × 10−4 | 1151.22 | 272.13 | 4.230 | 3.21 × 10−3 | 1108.09 | 328.9 | 3.369 |
| COL1A2 | DQGPVGRTGEVGAVGpPGFAGEKGPSGEAGTAGPpGTpGP | 8.56 × 10−4 | 196.08 | 75.14 | 2.610 | 1.22 × 10−2 | 168.37 | 83.85 | 2.008 |
| AHSG | SLGSPSGEVSHPRKT | 8.72 × 10−4 | 2282.82 | 975.54 | 2.340 | 4.44 × 10−4 | 2515.71 | 1480.13 | 1.7 |
| AHSG | VVSLGSPSGEVSHPRKT | 9.37 × 10−3 | 11,404.37 | 7829.98 | 1.457 | 3.12 × 10−2 | 13,131.49 | 12,697.43 | 1.034 |
| PIGR | LFAEEKAVADTRDQADGSRASVDSGSSEEQGGSSRA | 1.22 × 10−2 | 774.23 | 654.63 | 1.183 | 2.49 × 10−3 | 624.32 | 360.05 | 1.734 |
| COL1A2 | VGRTGEVGAVGPpGFAGEKGPSGEAGTAGPpGTpGP | 1.82 × 10−2 | 109.41 | 46.41 | 2.357 | 2.91 × 10−1 | 92.33 | 59.3 | 1.557 |
| COL3A1 | ARGLpGppGSNGNPGPPGPSGSPGKDGPPGPAGNTGAPG | 2.34 × 10−2 | 902.6 | 594.85 | 1.517 | 1.01 × 10−1 | 889.74 | 771.39 | 1.153 |
| SERPINC1 | FSPEKSKLPGIVAEGRDDLYVSDAFHKAF | 2.34 × 10−2 | 9438.98 | 5838.02 | 1.617 | 1.80 × 10−3 | 11,768.37 | 8101.28 | 1.453 |
| COL2A1 | GETGAAGpPGpAGPAGERGEQGAPGP | 2.34 × 10−2 | 43.02 | 135 | 0.319 | 1.03 × 10−2 | 58.08 | 160.81 | 0.361 |
| COL4A1 | pGIPGFPGSKGEMGVMGTPGQPGSPGPVGAPGLPGEKGDH | 2.34 × 10−2 | 3045.2 | 1456.47 | 2.091 | 4.89 × 10−2 | 2563.4 | 1629 | 1.574 |
| COL1A1 | ANGApGNDGAKGDAGApGApGSQGApGLQGMpGERGAAGLPGp | 2.69 × 10−2 | 1210.38 | 814.9 | 1.485 | 1.94 × 10−1 | 1267.81 | 1012.08 | 1.253 |
| COL3A1 | ApGPAGSRGApGPQGpRGDKGETGERG | 2.69 × 10−2 | 1103.52 | 618.43 | 1.784 | 1.27 × 10−1 | 857.9 | 692.61 | 1.239 |
| COL1A1 | GADGQPGAKGEpGDAGAKGDAGPpGPAGP | 2.69 × 10−2 | 108.1 | 388.09 | 0.279 | 7.21 × 10−3 | 106.07 | 519.75 | 0.204 |
| COL1A1 | ANGApGNDGAKGDAGApGApGSQGApGLQGMpGERGAAGLpGp | 2.74 × 10−2 | 447.16 | 170.81 | 2.618 | 2.04 × 10−1 | 470.53 | 214.82 | 2.19 |
| HBA1 | AAHLPAEFTPAVHASLDKFL | 2.81 × 10−2 | 610.19 | 14,067.56 | 0.043 | 9.65 × 10−3 | 903.23 | 19,261.46 | 0.047 |
| COL1A1 | ADGQpGAKGEpGDAGAKGDAGPPGPAGP | 2.81 × 10−2 | 212.86 | 365.65 | 0.582 | 1.01 × 10−1 | 217.32 | 302.79 | 0.718 |
| COL3A1 | EGGKGAAGpPGPpGAAGTpGLQG | 2.81 × 10−2 | 689.84 | 500.7 | 1.378 | 7.94 × 10−2 | 705.78 | 551.83 | 1.279 |
| COL22A1 | GTEGKKGEAGPPGLPGPpGIAGpQGSQGERGADGEVGQKGDQGHPGVPGFMGPPGNPGP | 2.81 × 10−2 | 192.19 | 159.22 | 1.207 | 4.74 × 10−2 | 171.5 | 166.83 | 1.028 |
| AHSG | GVVSLGSPSGEVSHPRKT | 2.81 × 10−2 | 2476.64 | 1429.55 | 1.732 | 2.24 × 10−2 | 2876.18 | 2229.17 | 1.29 |
| PIGR | FAEEKAVADTRDQADGSRASVDSGSSEEQGGSSRALVSTLVPL | 3.06 × 10−2 | 891.67 | 377.71 | 2.361 | 3.43 × 10−2 | 861.08 | 320.08 | 2.69 |
| COL2A1 | ppGSNGNpGPPGPPGPSGKDGPKGARGDSGPPGRAGEPG | 3.50 × 10−2 | 412.14 | 184.53 | 2.233 | 2.54 × 10−1 | 396.08 | 243.4 | 1.627 |
| COL18A1 | DDILASPPRLPEPQPYPGAPHHSS | 3.77 × 10−2 | 611.67 | 433.29 | 1.412 | 5.12 × 10−1 | 560.24 | 524.1 | 1.069 |
| COL3A1 | EpGRDGVpGGPGm | 3.77 × 10−2 | 2254.17 | 1608.12 | 1.402 | 1.08 × 10−2 | 2160.1 | 1392.93 | 1.551 |
| HBA1 | AAHLPAEFTPAVHASLDKFLAS | 4.15 × 10−2 | 847.52 | 30,583.66 | 0.028 | 3.06 × 10−2 | 1046.13 | 30,595.91 | 0.034 |
| FGA | DEAGSEADHEGTHSTKRGHAKSRPV | 4.15 × 10−2 | 31,926.35 | 22,421.31 | 1.424 | 5.01 × 10−1 | 29,485.54 | 34,133.85 | 0.864 |
| AHSG | VSLGSPSGEVSHPRKT | 4.15 × 10−2 | 3680.2 | 2187.3 | 1.683 | 2.25 × 10−2 * | 2326.63 | 3525.8 | 0.66 * |
| COL3A1 | GpGSDGKPGPpG | 4.86 × 10−2 | 145.26 | 347.21 | 0.418 | 2.28 × 10−2 | 192.46 | 399.37 | 0.482 |
| COL1A1 | GSpGSpGPDGKTGPPGPAG | 4.86 × 10−2 | 74.29 | 178.76 | 0.416 | 4.19 × 10−2 | 68.72 | 157.74 | 0.436 |
| Cleaved Proteins | Protease (Gene) | n of Cleaving Sites | Fold Change | Average Fibrosis | Average No Fibrosis | p | |
|---|---|---|---|---|---|---|---|
| ↓ | ↑ | ||||||
| HBA1 (5) | Cathepsin D (CTSD) | 5 | 0 | 0.03 | 752.59 | 23977.22 | 0.0006 |
| COL2A1 (1) | Macrophage metalloelastase (MMP12), Neutrophil collagenase (MMP8) | 1 | 0 | 0.32 | 43.02 | 135.00 | 0.0002 |
| COL1A1 (1), COL1A2 (2) | 72 kDa type IV collagenase (MMP2) | 1 | 2 | 1.26 | 126.59 | 100.10 | 0.1580 |
| COL2A1 (1), COL18A1 (2), COL1A2 (2) | Collagenase 3 (MMP13) | 1 | 4 | 1.40 | 314.37 | 224.63 | 0.0002 |
| COL18A1 (1) | Cathepsin B (CTSB), Cathepsin K (CTSK), Procathepsin L (CTSL), Matrix metalloproteinase-20 (MMP20), Matrilysin (MMP7) | 0 | 1 | 1.41 | 611.67 | 433.29 | 0.0006 |
| COL18A1 (1), COL1A2 (2) | Matrix metalloproteinase-14 (MMP14) | 0 | 3 | 1.65 | 305.72 | 184.95 | 0.0001 |
| COL2A1 (1) | Interstitial collagenase (MMP1) | 0 | 1 | 2.23 | 412.14 | 184.53 | 0.0006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catanese, L.; Siwy, J.; Mavrogeorgis, E.; Amann, K.; Mischak, H.; Beige, J.; Rupprecht, H. A Novel Urinary Proteomics Classifier for Non-Invasive Evaluation of Interstitial Fibrosis and Tubular Atrophy in Chronic Kidney Disease. Proteomes 2021, 9, 32. https://doi.org/10.3390/proteomes9030032
Catanese L, Siwy J, Mavrogeorgis E, Amann K, Mischak H, Beige J, Rupprecht H. A Novel Urinary Proteomics Classifier for Non-Invasive Evaluation of Interstitial Fibrosis and Tubular Atrophy in Chronic Kidney Disease. Proteomes. 2021; 9(3):32. https://doi.org/10.3390/proteomes9030032
Chicago/Turabian StyleCatanese, Lorenzo, Justyna Siwy, Emmanouil Mavrogeorgis, Kerstin Amann, Harald Mischak, Joachim Beige, and Harald Rupprecht. 2021. "A Novel Urinary Proteomics Classifier for Non-Invasive Evaluation of Interstitial Fibrosis and Tubular Atrophy in Chronic Kidney Disease" Proteomes 9, no. 3: 32. https://doi.org/10.3390/proteomes9030032