
Special Nuclear Structures in the Germinal Vesicle of the Common Frog with Emphasis on the So-Called Karyosphere Capsule
 , and
, and 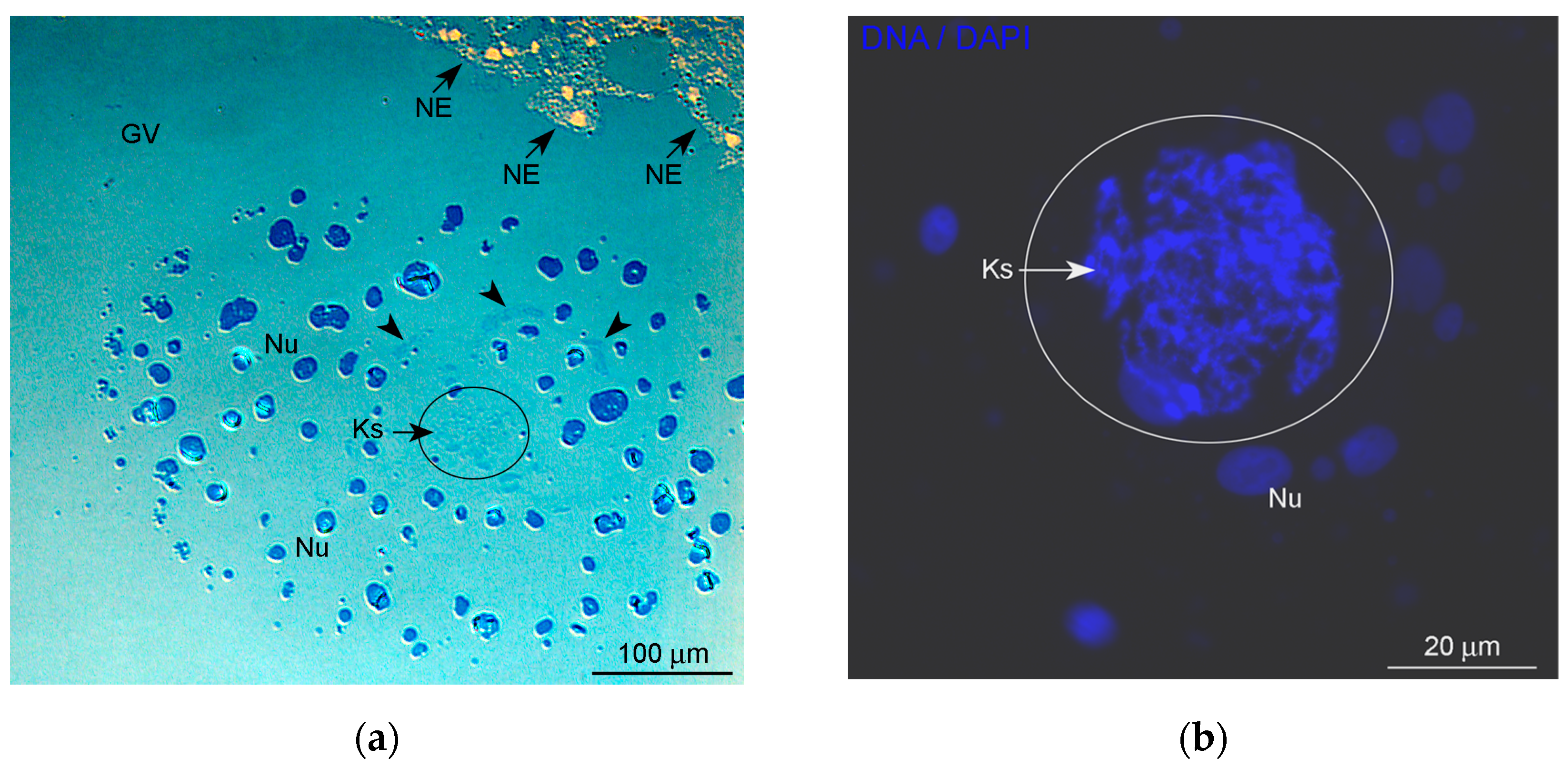{kind=link}
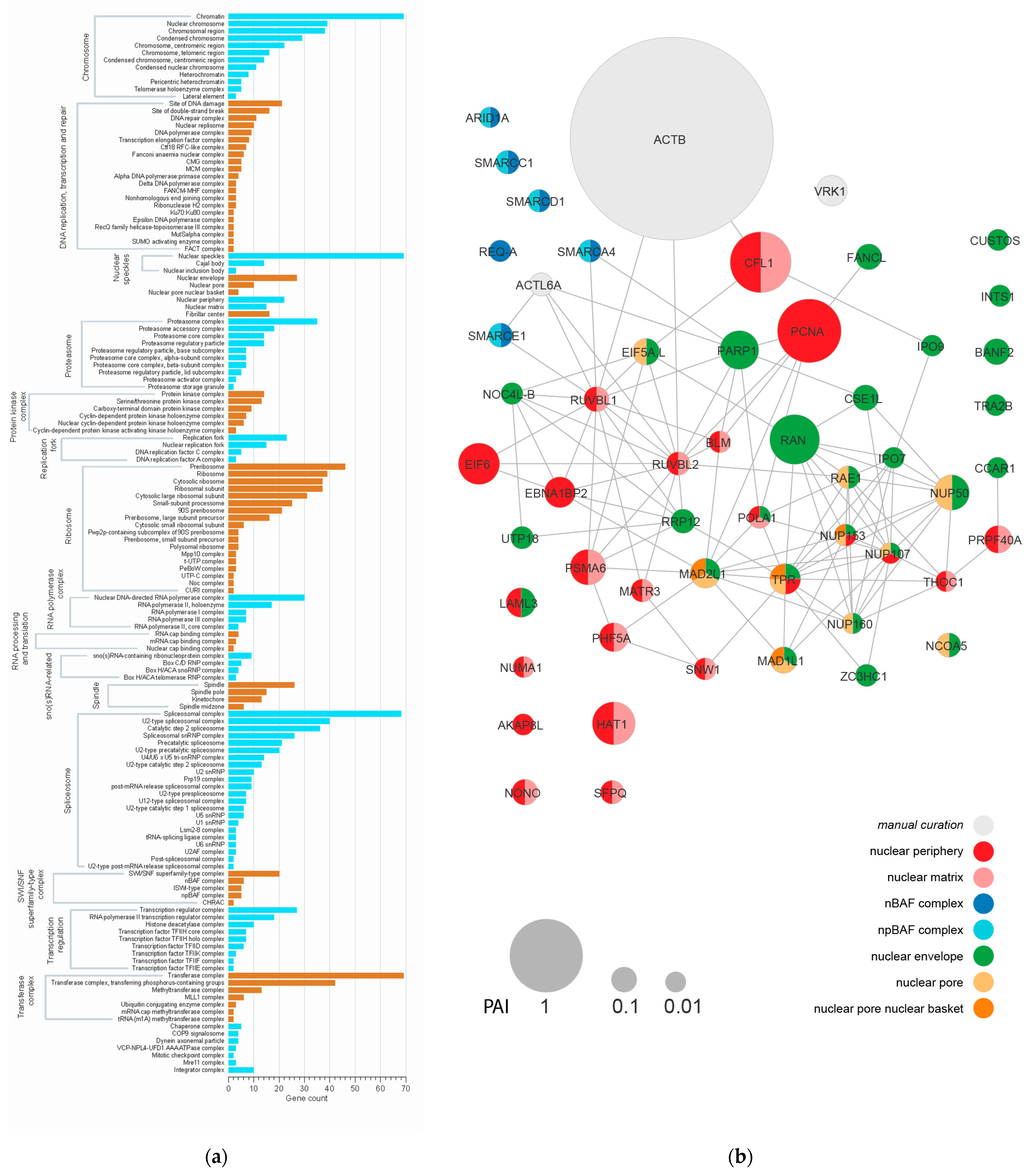{kind=link}
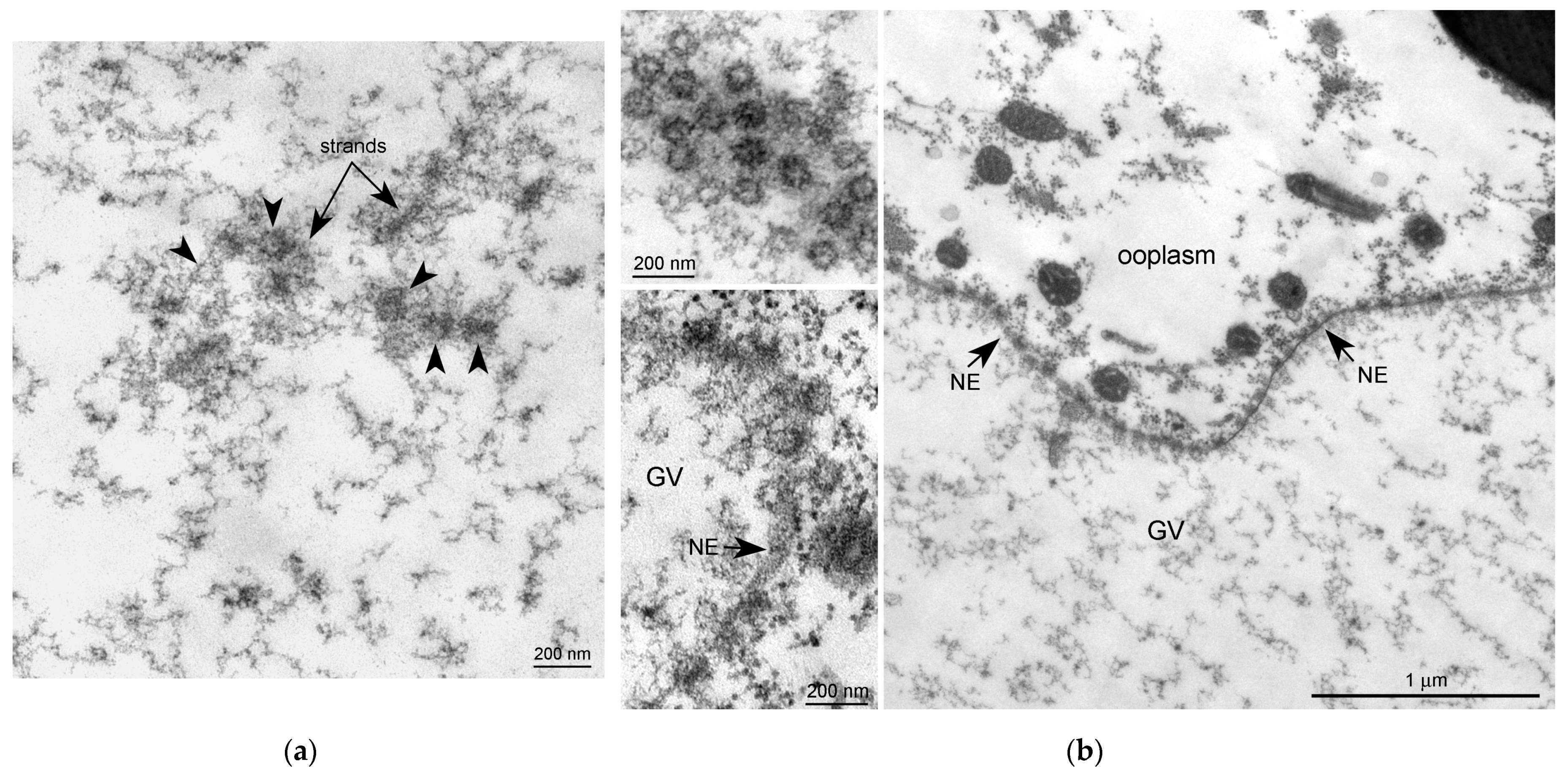{kind=link}
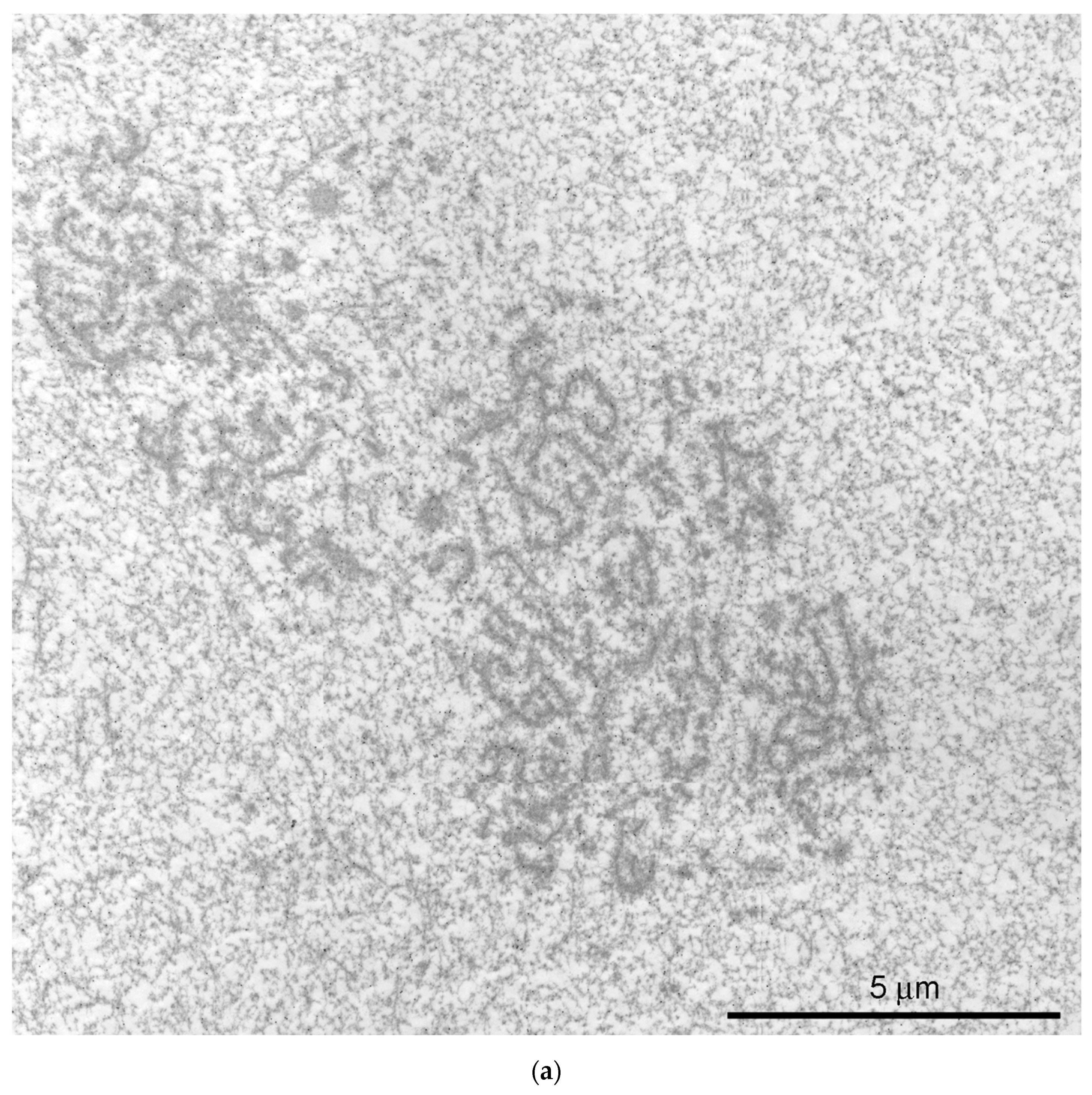{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
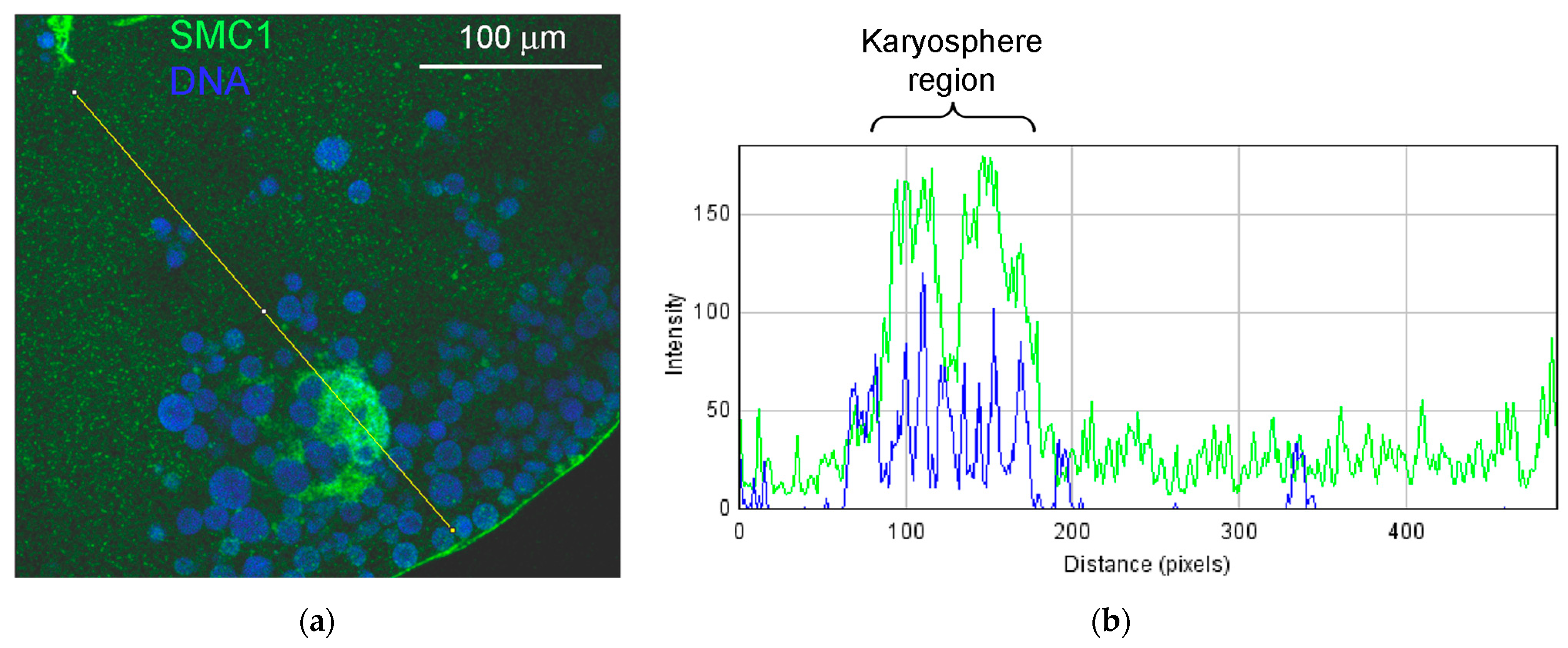{kind=link}
{kind=link}
{kind=link}
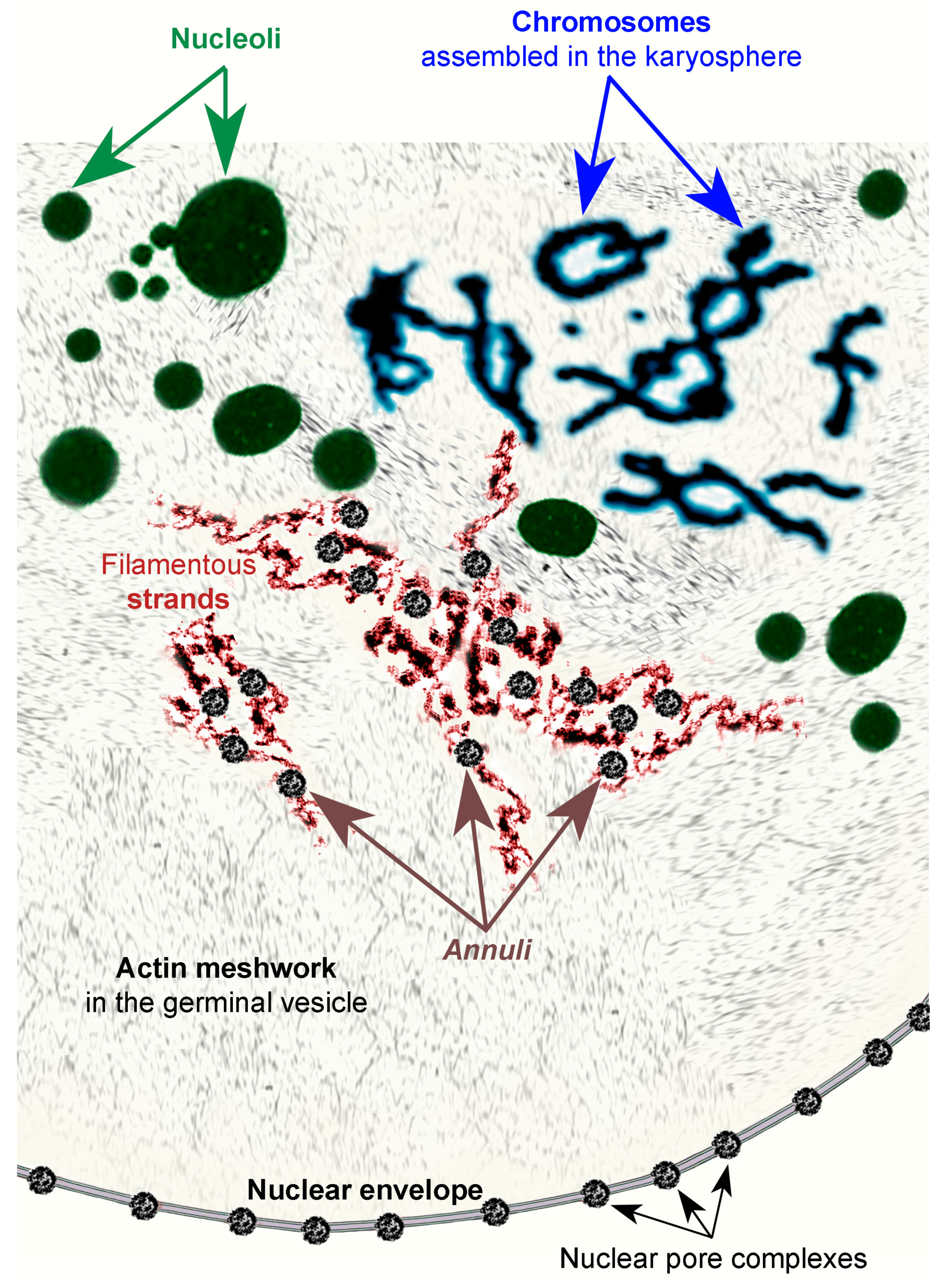{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Oocyte Retrieval
2.2. Primary Antibodies
2.3. Western Blot Analysis
2.4. Fluorescent Microscopy
2.5. Conventional Electron Microscopy
2.6. Immunoelectron Microscopy
Counting of Gold Labels and Statistical Analysis
2.7. Proteomic Analysis
3. Results
3.1. Proteomic Analysis of the Karyosphere
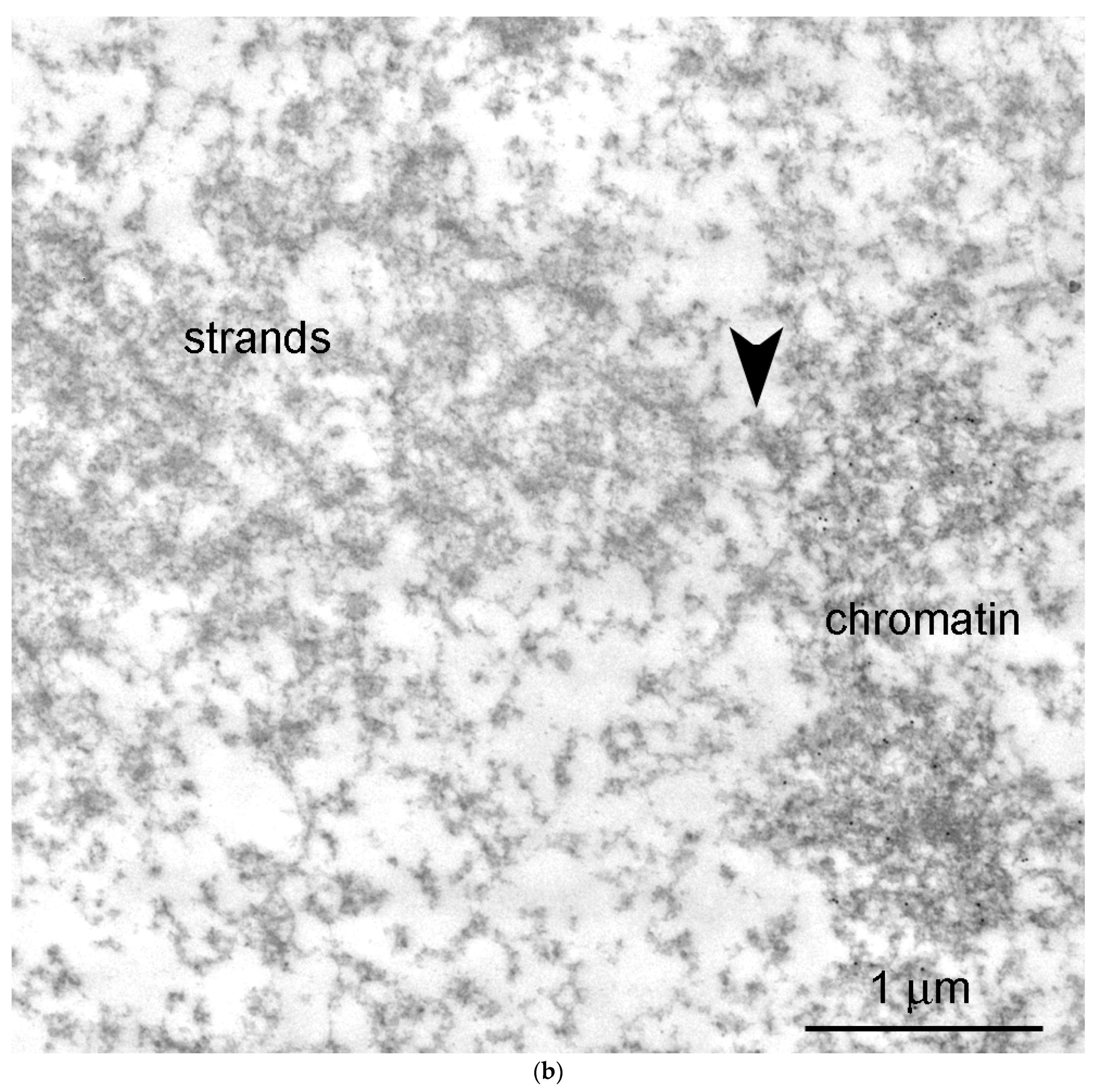
3.2. Morphology of Extrachromosomal Structures in Frog GV
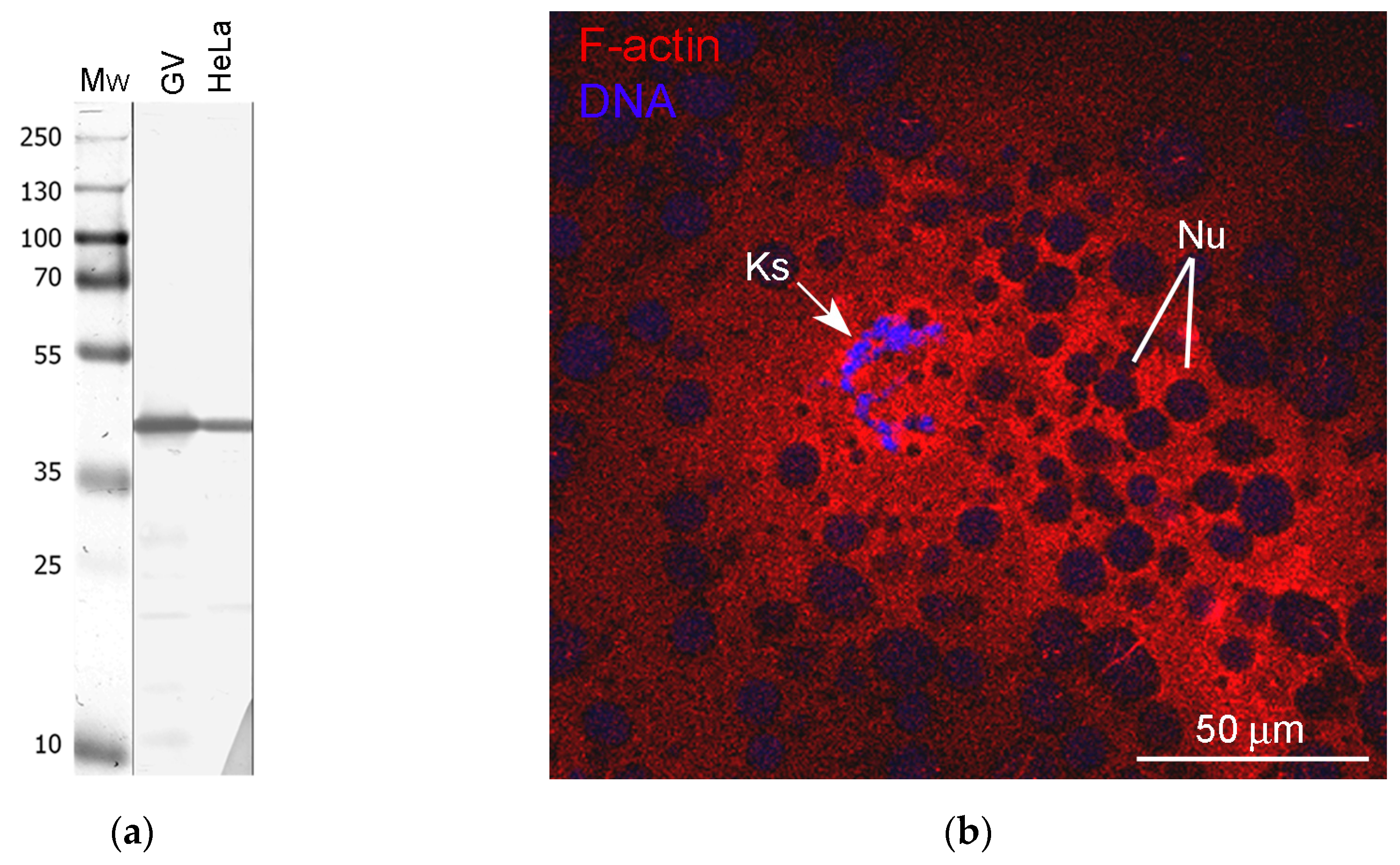
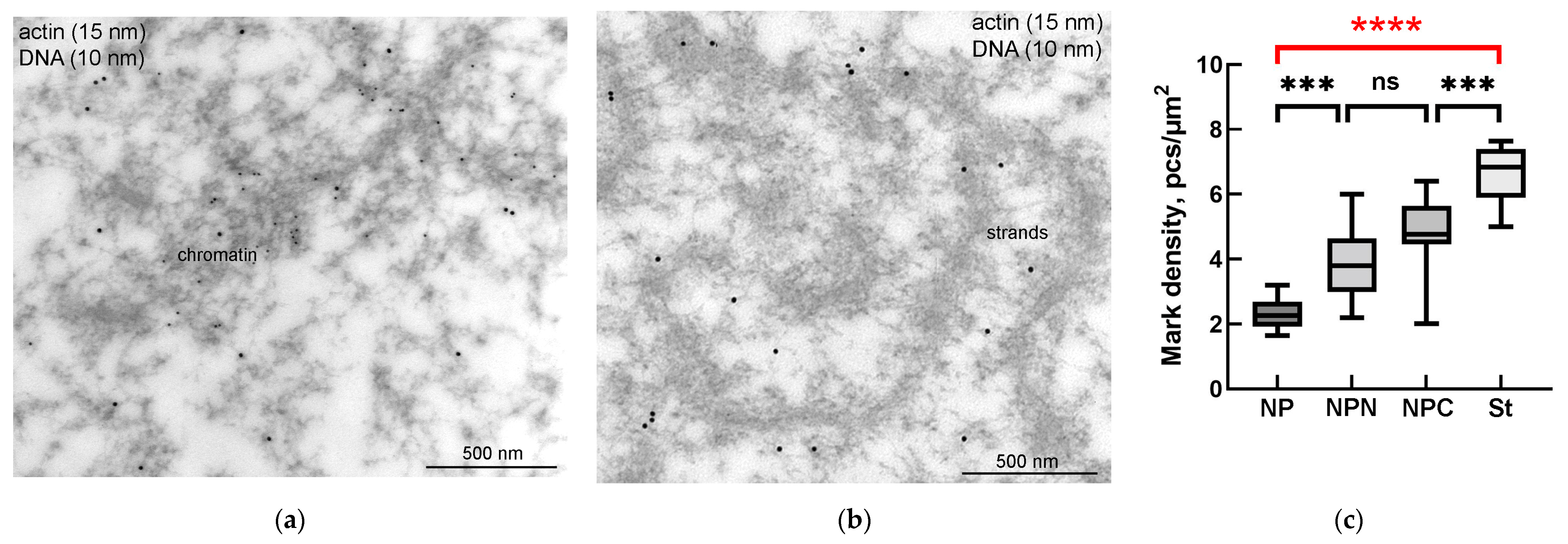
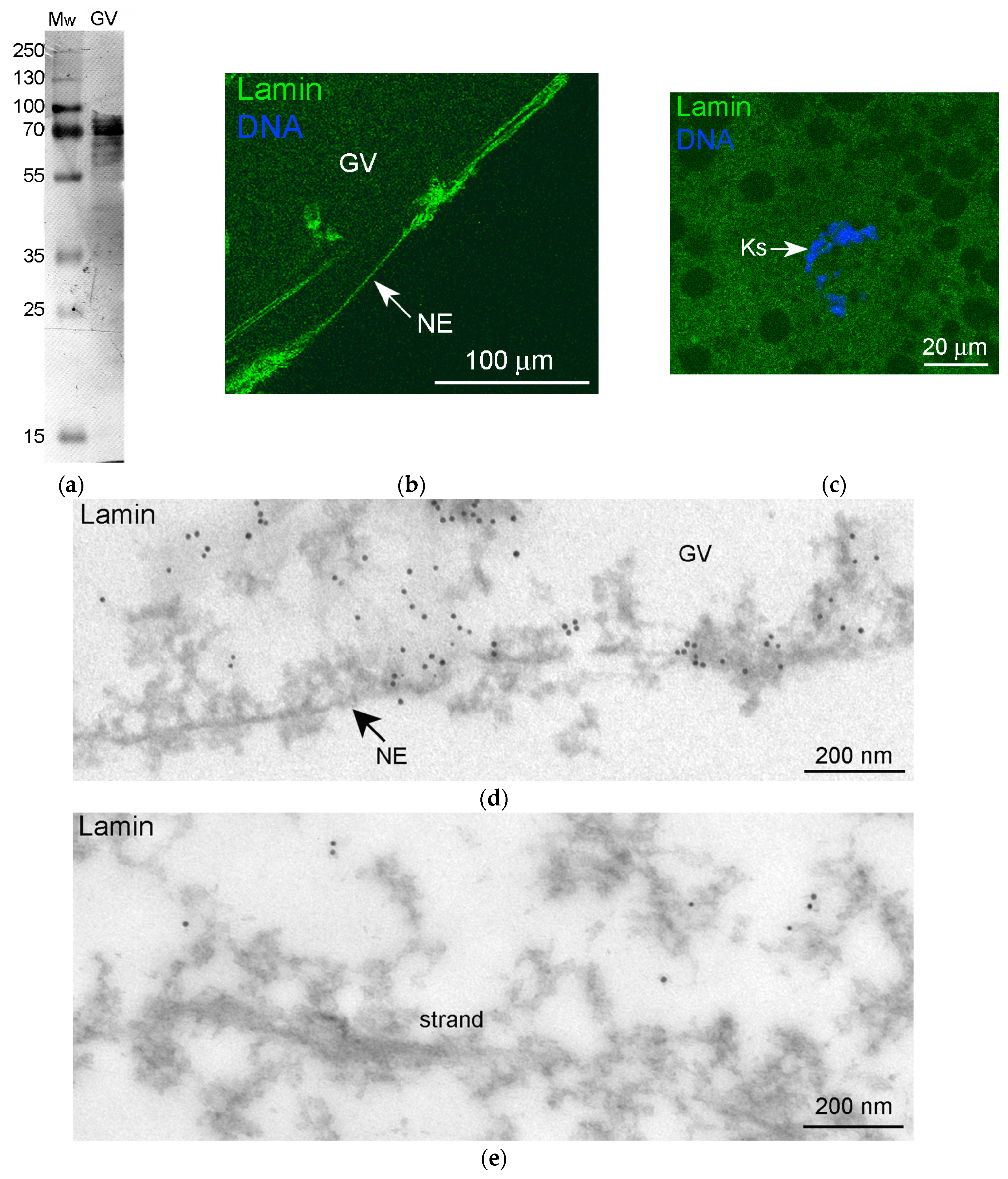
3.3. Nuclear Actin, Lamin, and the Extrachromosomal Strands
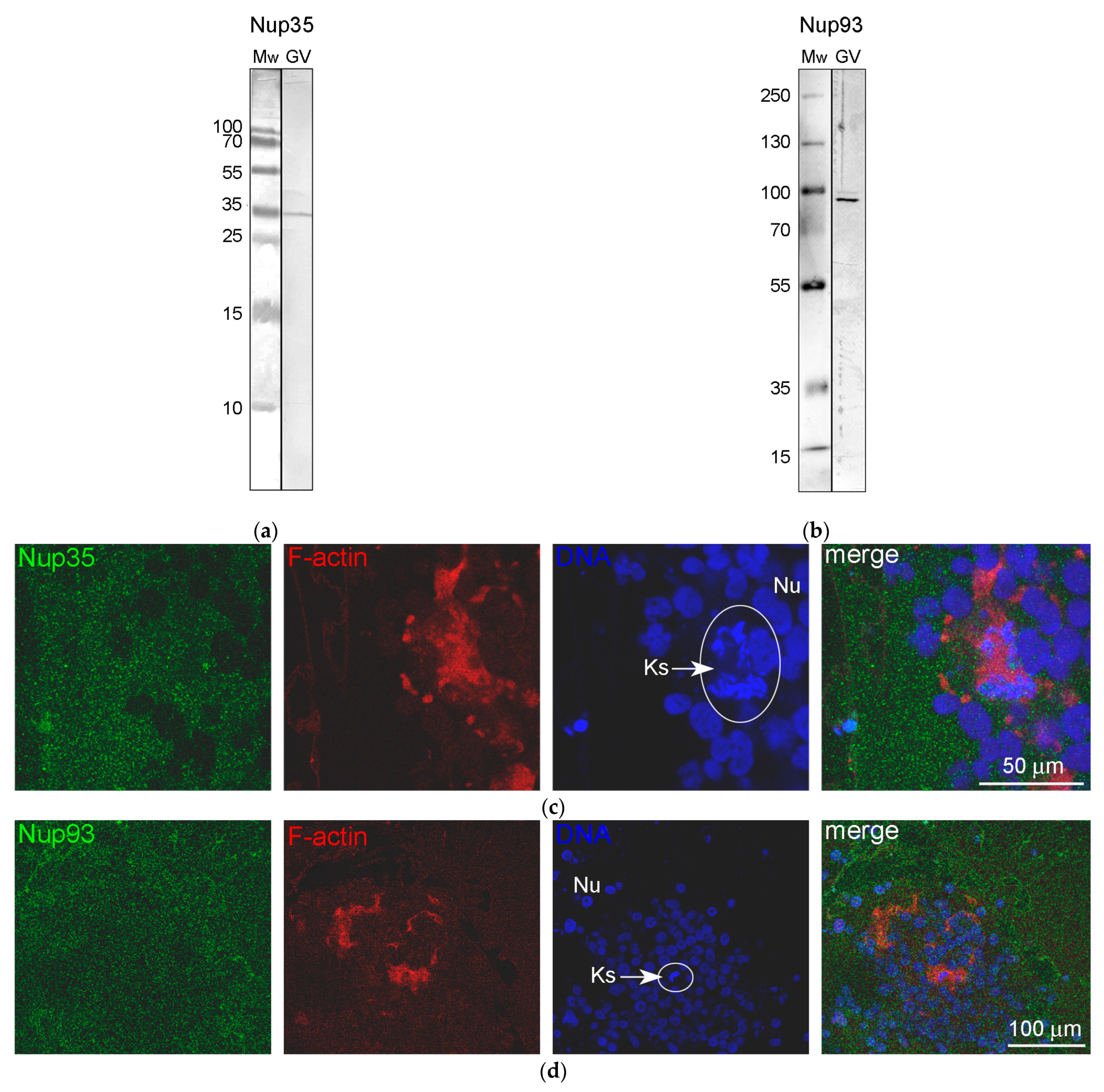
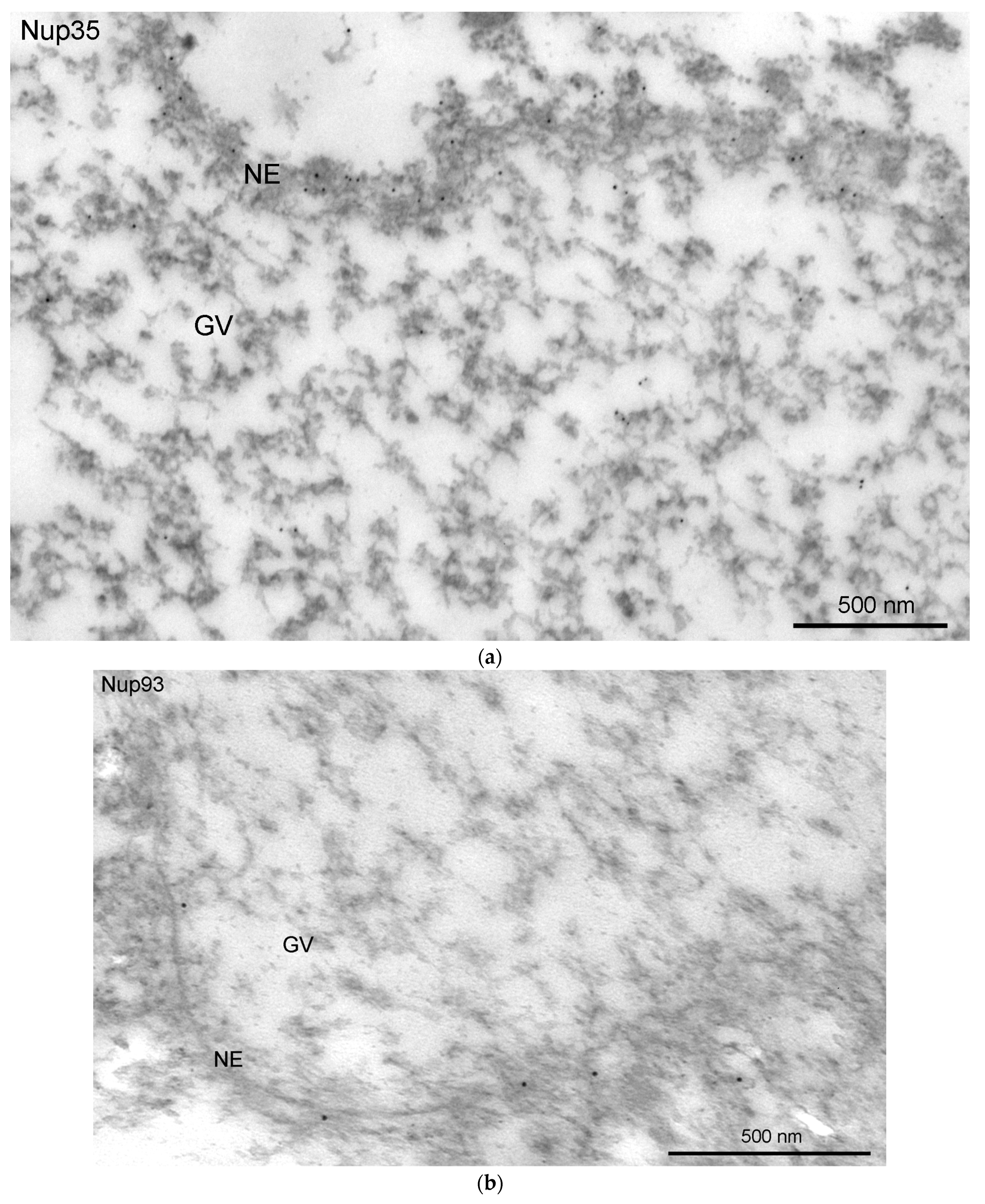
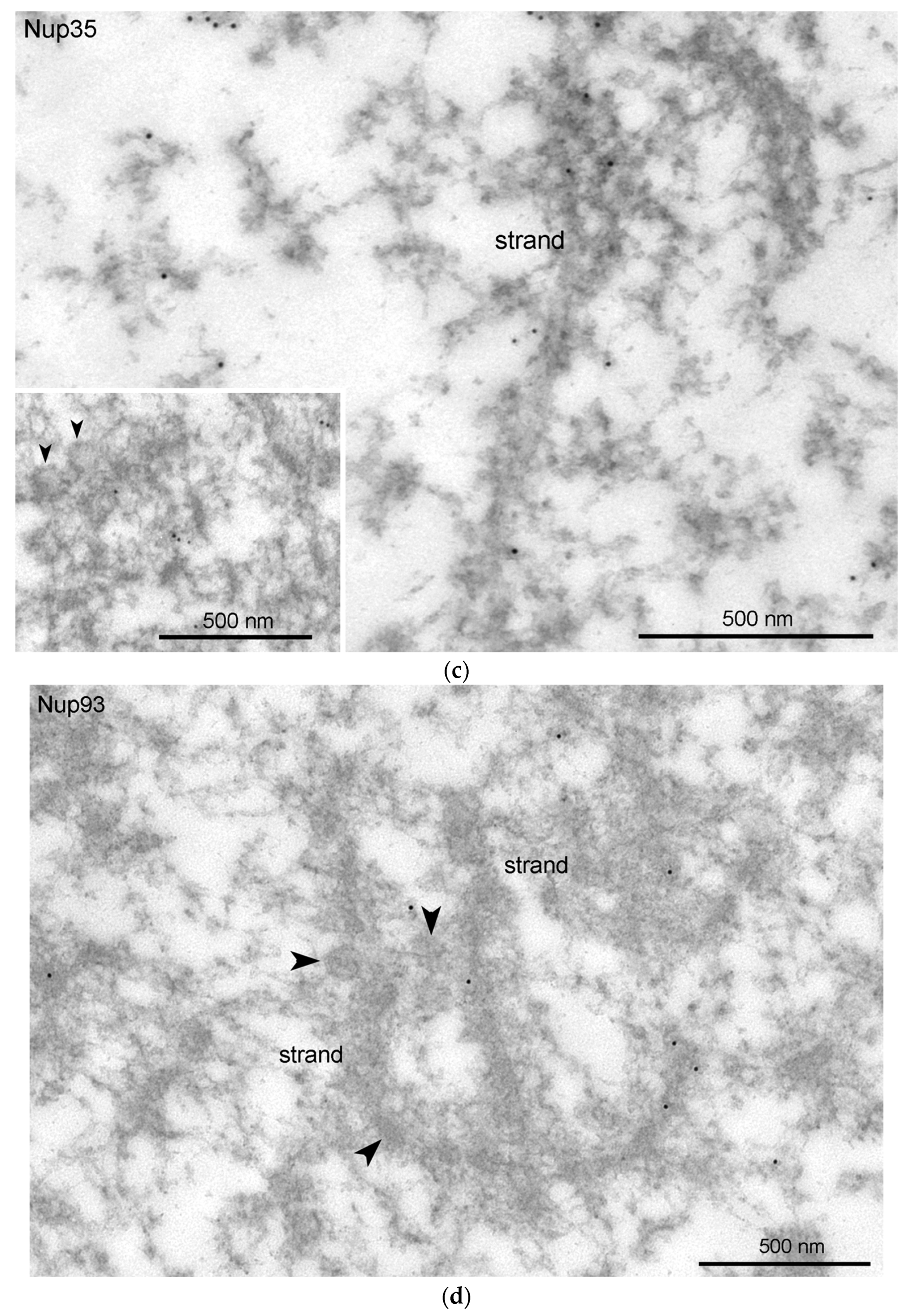
3.4. Nucleoporins and Annuli
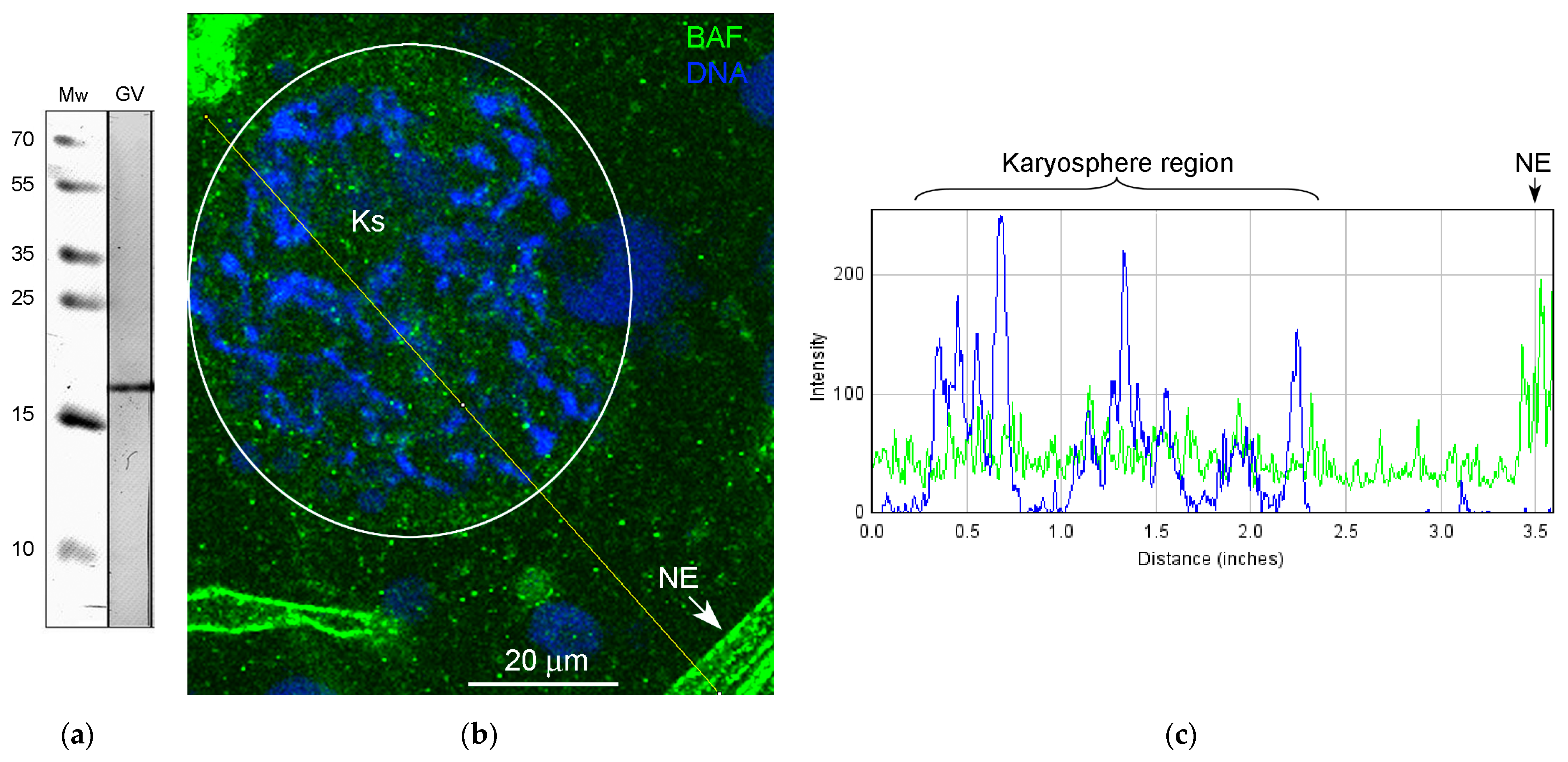
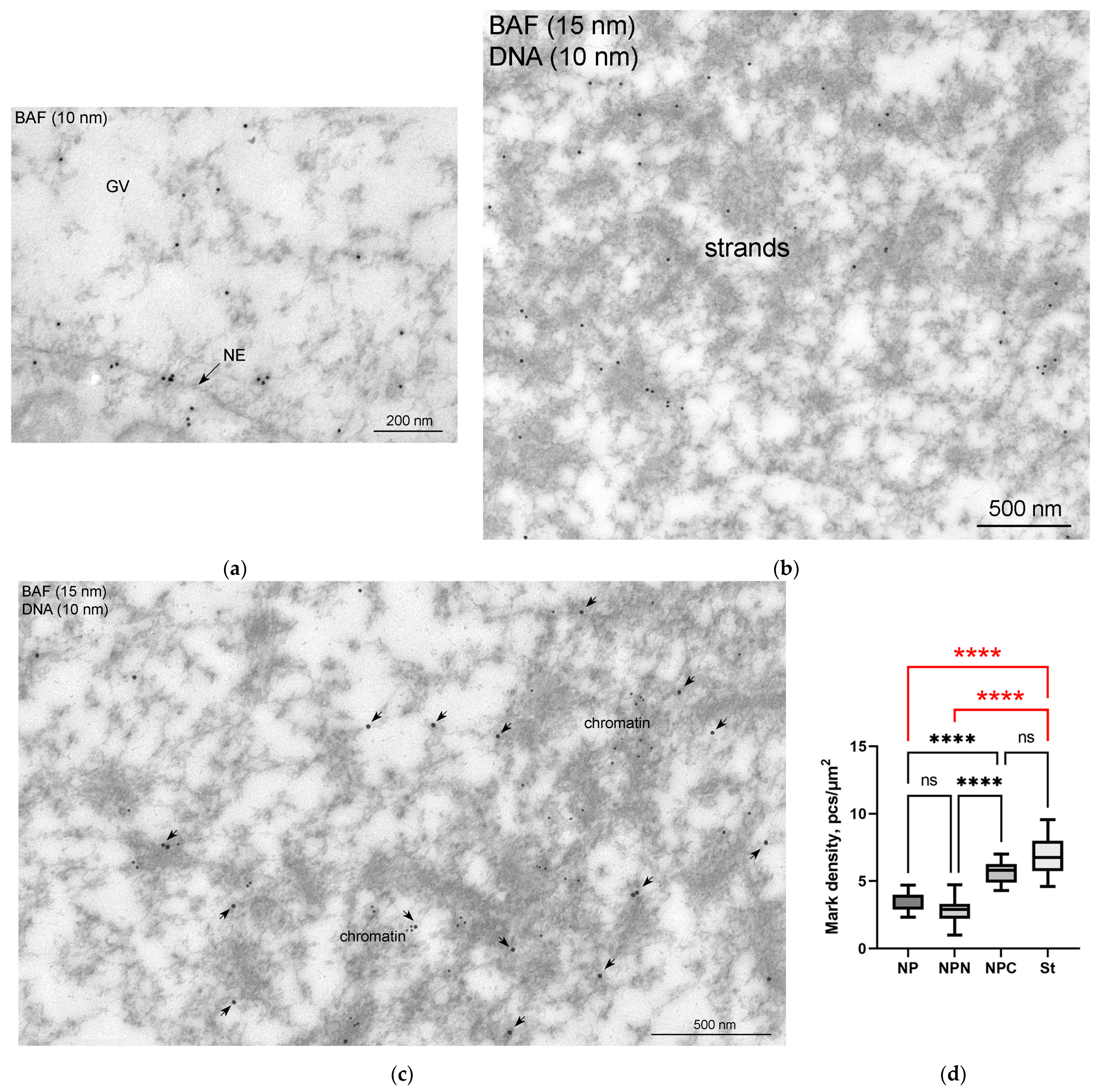
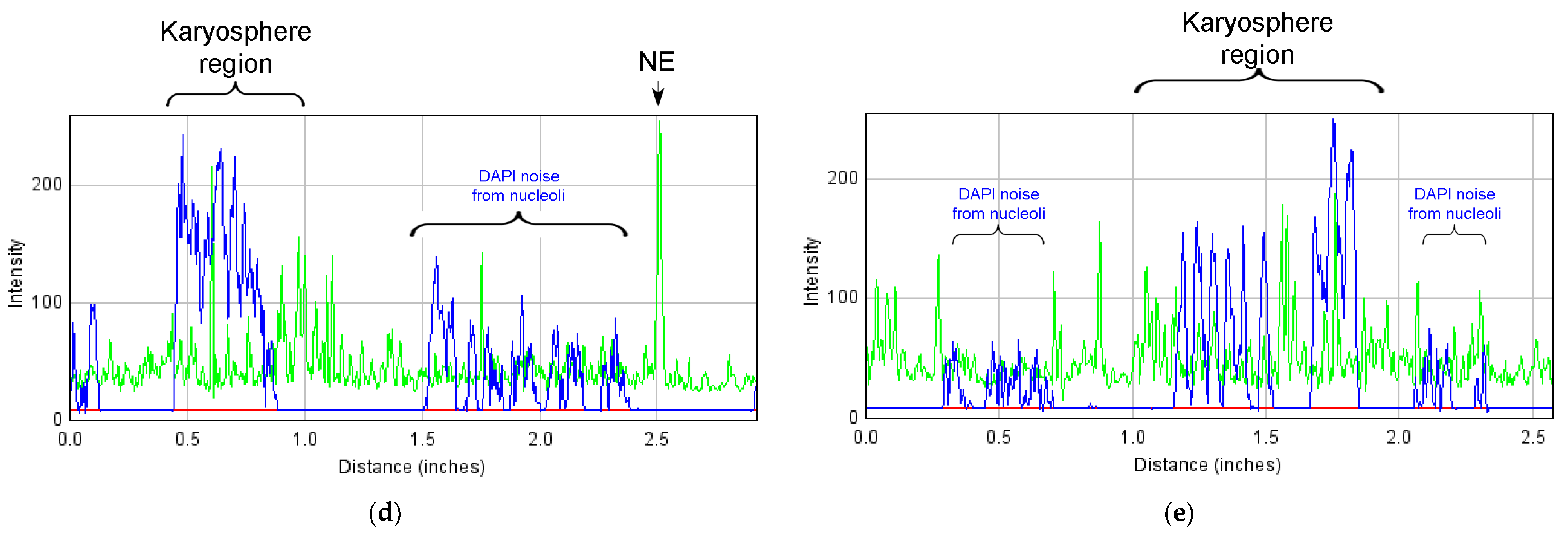
3.5. Nuclear Envelope-Related Proteins and the “Karyosphere Capsule”
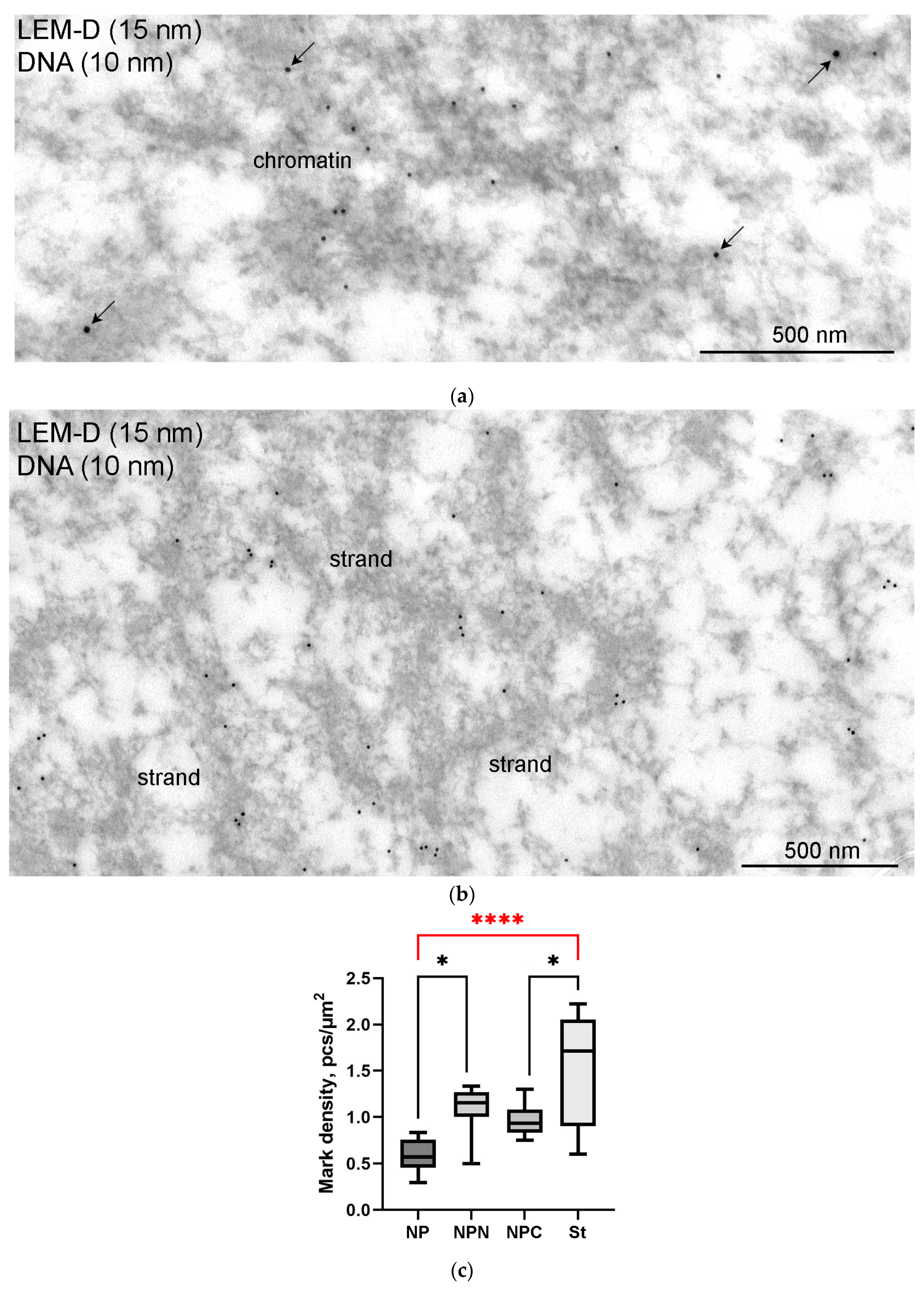
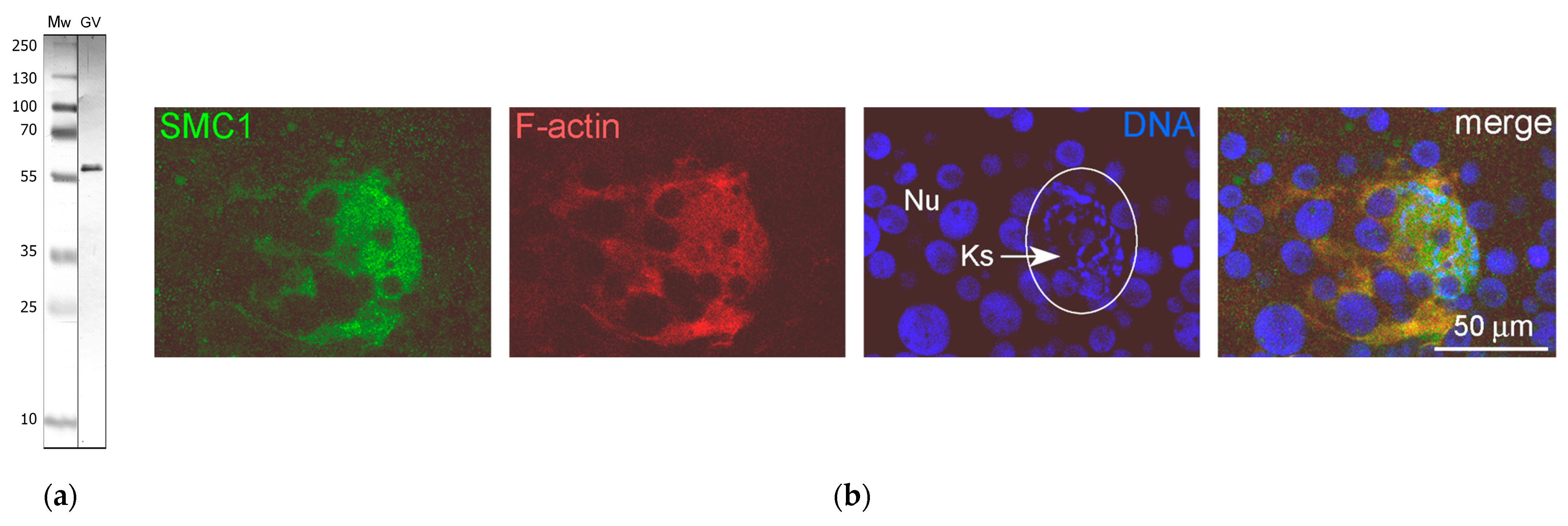
3.6. SMC1 Is a Component of the Strands
4. Discussion
4.1. What Does the Word “Capsule” Mean?
4.2. Extrachromosomal Strands Do Not Contain Structural Proteins Actin and Lamin B
4.3. Annuli Are Not the Autonomous Pore Complexes
4.4. BAF and LEMD-Proteins in the R. temporaria Germinal Vesicle
4.5. SMC1 Is a Component of the Strands
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gruenbaum, Y.; Goldman, R.D.; Meyuhas, R.; Mills, E.; Margalit, A.; Fridkin, A.; Dayani, Y.; Prokocimer, M.; Enosh, A. The nuclear lamina and its functions in the nucleus. Int. Rev. Cytol. 2023, 226, 1–62. [Google Scholar] [CrossRef]
- Barton, L.J.; Soshnev, A.A.; Geyer, P.K. Networking in the nucleus: A spotlight on LEM-domain proteins. Curr. Opin. Cell Biol. 2015, 34, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jamin, A.; Wiebe, M.S. Barrier to autointegration factor (BANF1): Interwoven roles in nuclear structure, genome integrity, innate immunity, stress responses and progeria. Curr. Opin. Cell Biol. 2015, 34, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Bogolyubova, I.O.; Bogolyubov, D.S. Functional interactions of BAF and LEM proteins in the formation of germ cells. Cell Tissue Biol. 2023, 17, 571–582. [Google Scholar]
- Zetka, M.; Paouneskou, D.; Jantsch, V. The nuclear envelope, a meiotic jack-of-all-trades. Curr. Opin. Cell Biol. 2020, 64, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, O.M.; Cullen, C.F.; Ohkura, H. NHK-1 phosphorylates BAF to allow karyosome formation in the Drosophila oocyte nucleus. J. Cell Biol. 2007, 179, 817–824. [Google Scholar] [CrossRef]
- Gruzova, M.N.; Parfenov, V.N. Karyosphere in oogenesis and intranuclear morphogenesis. Int. Rev. Cytol. 1993, 144, 1–52. [Google Scholar] [CrossRef]
- Bogolyubov, D.S. Karyosphere (karyosome): A peculiar structure of the oocyte nucleus. Int. Rev. Cell Mol. Biol. 2018, 337, 1–48. [Google Scholar] [CrossRef]
- Flora, P.; McCarthy, A.; Upadhyay, M.; Rangan, P. Role of chromatin modifications in Drosophila germline stem cell differentiation. Results Probl. Cell Differ. 2017, 59, 1–30. [Google Scholar] [CrossRef]
- Singh, A.; Dutta, D.; Paul, M.S.; Verma, D.; Mutsuddi, M.; Mukherjee, A. Pleiotropic functions of the chromodomain-containing protein Hat-trick during oogenesis in Drosophila melanogaster. G3 Bethesda 2018, 8, 1067–1077. [Google Scholar] [CrossRef]
- Fedorova, E.V.; Dorogova, N.V.; Bolobolova, E.U.; Fedorova, S.A.; Karagodin, D.A.; Ogienko, A.A.; Khruscheva, A.S.; Baricheva, E.M. GAGA protein is required for multiple aspects of Drosophila oogenesis and female fertility. Genesis 2019, 57, e23269. [Google Scholar] [CrossRef]
- Nieken, K.J.; O’Brien, K.; McDonnell, A.; Zhaunova, L.; Ohkura, H. A large-scale RNAi screen reveals that mitochondrial function is important for meiotic chromosome organization in oocytes. Chromosoma 2023, 132, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Blackman, M.W. The spermatogenesis of the myriapods. I. Notes on the spermatocytes and spermatids of Scolopendra. Kansas Univ. Quart. 1901, 10, 61–76. [Google Scholar]
- Loh, B.J.; Cullen, C.F.; Vogt, N.; Ohkura, H. The conserved kinase SRPK regulates karyosome formation and spindle microtubule assembly in Drosophila oocytes. J. Cell Sci. 2012, 125, 4457–4462. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, V.; Markova, M.; Zhivkova, R.; Chakarova, I.; Hadzhinesheva, V.; Delimitreva, S. Kariosphere, the enigmatic “Surrounded Nucleolus” of maturing oocytes: Review. Acta Morphol. Anthropol. 2017, 24, 78–84. [Google Scholar]
- Bogolyubova, I.; Bogolyubov, D. Heterochromatin morphodynamics in late oogenesis and early embryogenesis of mammals. Cells 2020, 9, 1497. [Google Scholar] [CrossRef] [PubMed]
- Bogolyubova, I.; Salimov, D.; Bogolyubov, D. Chromatin configuration in diplotene mouse and human oocytes during the period of transcriptional activity extinction. Int. J. Mol. Sci. 2023, 24, 11517. [Google Scholar] [CrossRef]
- Wagner, K. Über die Entwicklung des Froscheies. Arch. Zellforsch. 1923, 17, 1–44. [Google Scholar]
- Gruzova, M.N.; Parfenov, V.N. Ultrastructure of late oocyte nuclei in Rana temporaria. J. Cell Sci. 1977, 28, 1–13. [Google Scholar] [CrossRef]
- Gruzova, M.N.; Zaichikova, Z.P.; Sokolov, I.I. Functional organization of the nucleus in the oogenesis of Chrysopa perla L. (Insecta, Neuroptera). Chromosoma 1972, 37, 353–385. [Google Scholar] [CrossRef]
- Schultze, O. Untersuchungen über die Reifung und Befruchtung des Amphibieneies. Z. wiss. Zool. 1887, 45, 177–226. [Google Scholar]
- Gall, J.G.; Wu, Z.; Murphy, C.; Gao, H. Structure in the amphibian germinal vesicle. Exp. Cell Res. 2004, 296, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.G. Exporting actin. Nat. Cell Biol. 2006, 8, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Parfenov, V.N.; Davis, D.S.; Pochukalina, G.N.; Sample, C.E.; Bugaeva, E.A.; Murti, K.G. Nuclear actin filaments and their topological changes in frog oocytes. Exp. Cell Res. 1995, 217, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Świątek, P. Formation of the karyosome in developing oocytes of weevils (Coleoptera, Curculionidae). Tissue Cell 1999, 31, 587–593. [Google Scholar] [CrossRef]
- Rübsam, R.; Büning, J. F-actin is a component of the karyosome in neuropteran oocyte nuclei. Arthropod Struct. Dev. 2001, 30, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Bogolyubov, D.S.; Batalova, F.M.; Kiselyov, A.M.; Stepanova, I.S. Nuclear structures in Tribolium castaneum oocytes. Cell Biol. Int. 2013, 37, 1061–1079. [Google Scholar] [CrossRef]
- Ilicheva, N.; Podgornaya, O.; Bogolyubov, D.; Pochukalina, G. The karyosphere capsule in Rana temporaria oocytes contains structural and DNA-binding proteins. Nucleus 2018, 9, 516–529. [Google Scholar] [CrossRef]
- Duryee, W.R. Chromosomal physiology in relation to nuclear structure. Ann. N. Y. Acad. Sci. 1950, 50, 920–953. [Google Scholar] [CrossRef]
- Bogolyubova, I.O.; Bogolyubov, D.S. Effect of hormonal stimulation on the oocyte chromosomal apparatus in the common frog. J. Evol. Biochem. Physiol. 2023, 59, 1229–1236. [Google Scholar] [CrossRef]
- Wallace, R.A.; Jared, D.W.; Dumont, J.N.; Sega, M.W. Protein incorporation by isolated amphibian oocytes. III. Optimum incubation conditions. J. Exp. Zool. 1973, 184, 321–333. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.G.; Stephenson, E.C.; Erba, H.P.; Diaz, M.O.; Barsacchi-Pilone, G. Histone genes are located at the sphere loci of newt lampbrush chromosomes. Chromosoma 1981, 84, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape stringapp: Network analysis and visualization of proteomics data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Bogolyubov, D.S.; Travina, A.O.; Bogolyubova, I.O. Karyosphere capsule in oocytes of the grass frog: To be or not to be? A critical view. Cell Tissue Biol. 2022, 16, 521–539. [Google Scholar] [CrossRef]
- Ilicheva, N.V.; Pochukalina, G.N.; Podgornaya, O.I. Actin depolymerization disrupts karyosphere capsule integrity but not residual transcription in late oocytes of the grass frog Rana temporaria. J. Cell. Biochem. 2019, 120, 15057–15068. [Google Scholar] [CrossRef]
- Stick, R. cDNA cloning of the developmentally regulated lamin LIII of Xenopus laevis. EMBO J. 1988, 7, 3189–3197. [Google Scholar] [CrossRef]
- Dultz, E.; Wojtynek, M.; Medalia, O.; Onischenko, E. The nuclear pore complex: Birth, life, and death of a cellular behemoth. Cells 2022, 11, 1456. [Google Scholar] [CrossRef]
- Burla, R.; La Torre, M.; Maccaroni, K.; Verni, F.; Giunta, S.; Saggio, I. Interplay of the nuclear envelope with chromatin in physiology and pathology. Nucleus 2020, 11, 205–218. [Google Scholar] [CrossRef]
- Gu, M.; LaJoie, D.; Chen, O.S.; von Appen, A.; Ladinsky, M.S.; Redd, M.J.; Nikolova, L.; Bjorkman, P.J.; Sundquist, W.I.; Ullman, K.S.; et al. LEM2 recruits CHMP7 for ESCRT-mediated nuclear envelope closure in fission yeast and human cells. Proc. Natl. Acad. Sci. USA 2017, 114, E2166–E2175. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Shinohara, A. Chromosome architecture and homologous recombination in meiosis. Front. Cell Dev. Biol. 2023, 10, 1097446. [Google Scholar] [CrossRef] [PubMed]
- Matityahu, A.; Onn, I. It’s all in the numbers: Cohesin stoichiometry. Front. Mol. Biosci. 2022, 9, 1010894. [Google Scholar] [CrossRef]
- Hoencamp, C.; Rowland, B.D. Genome control by SMC complexes. Nat. Rev. Mol. Cell Biol. 2023, 24, 633–650. [Google Scholar] [CrossRef] [PubMed]
- Ivanovska, I.; Khandan, T.; Ito, T.; Orr-Weaver, T.L. A histone code in meiosis: The histone kinase, NHK-1, is required for proper chromosomal architecture in Drosophila oocytes. Gen. Dev. 2005, 19, 2571–2582. [Google Scholar] [CrossRef] [PubMed]
- Oldenkamp, R.; Rowland, B.D. A walk through the SMC cycle: From catching DNAs to shaping the genome. Mol. Cell 2022, 82, 1616–1630. [Google Scholar] [CrossRef] [PubMed]
- Nasmyth, K.; Haering, C.H. The structure and function of SMC and kleisin complexes. Annu. Rev. Biochem. 2005, 74, 595–648. [Google Scholar] [CrossRef]
- Blackman, M.W. The spermatogenesis of the myriapods. II. On the chromatin in the spermatocytes of Scolopendra heros. Biol. Bull. 1903, 5, 187–217. [Google Scholar] [CrossRef]
- Liu, J.-L.; Buszczak, M.; Gall, J.G. Nuclear bodies in the Drosophila germinal vesicle. Chromosome Res. 2006, 14, 465–475. [Google Scholar] [CrossRef]
- Razin, S.V.; Iarovaia, O.V.; Vassetzky, Y.S. A requiem to the nuclear matrix: From a controversial concept to 3D organization of the nucleus. Chromosoma 2014, 123, 217–224. [Google Scholar] [CrossRef]
- Pochukalina, G.N.; Parfenov, V.N. The organization of the karyosphere and capsule before oocyte maturation in the common frog. Tsitologiia 1994, 36, 1027–1034. [Google Scholar] [PubMed]
- Bogolyubov, D.; Alexandrova, O.; Tsvetkov, A.; Parfenov, V. An immunoelectron study of karyosphere and nuclear bodies in oocytes of mealworm beetle, Tenebrio molitor (Coleoptera: Polyphaga). Chromosoma 2000, 109, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Falahzadeh, K.; Banaei-Esfahani, A.; Shahhoseini, M. The potential roles of actin in the nucleus. Cell J. 2015, 17, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Clark, T.G.; Rosenbaum, J.L. An actin filament matrix in hand-isolated nuclei of X. laevis oocytes. Cell 1979, 18, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, M.T.; Stüven, T.; Kuhn, C.; Cordes, V.C.; Görlich, D. A selective block of nuclear actin export stabilizes the giant nuclei of Xenopus oocytes. Nat. Cell Biol. 2006, 8, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Clark, T.G.; Merriam, R.W. Diffusible and bound actin nuclei of Xenopus laevis oocytes. Cell 1977, 12, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Morozova, K.N.; Kiseleva, E.V. Nuclear and cytoplasmic organization in Xenopus oocytes after disruption of actin filaments by latrunculin. Cell Tissue Biol. 2008, 2, 300–310. [Google Scholar] [CrossRef]
- Plessner, M.; Grosse, R. Dynamizing nuclear actin filaments. Curr. Opin. Cell Biol. 2019, 56, 1–6. [Google Scholar] [CrossRef]
- Kiseleva, E.; Drummond, S.P.; Goldberg, M.W.; Rutherford, S.A.; Allen, T.D.; Wilson, K.L. Actin- and protein-4.1-containing filaments link nuclear pore complexes to subnuclear organelles in Xenopus oocyte nuclei. J. Cell Sci. 2004, 117, 2481–2490. [Google Scholar] [CrossRef]
- Maslova, A.; Krasikova, A. Nuclear actin depolymerization in transcriptionally active avian and amphibian oocytes leads to collapse of intranuclear structures. Nucleus 2012, 3, 300–311. [Google Scholar] [CrossRef]
- Feric, M.; Brangwynne, C.P. A nuclear F-actin scaffold stabilizes ribonucleoprotein droplets against gravity in large cells. Nat. Cell Biol. 2013, 15, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Brangwynne, C.P.; Mitchison, T.J.; Hyman, A.A. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 4334–4339. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, V.; Delimitreva, S.; Chakarova, I.; Zhivkova, R.; Hadzhinesheva, V.; Markova, M. Dynamics of lamins B and A/C and nucleoporin Nup160 during meiotic maturation in mouse oocytes. Folia Biol. Praha 2017, 63, 6–12. [Google Scholar] [PubMed]
- Pochukalina, G.N.; Ilicheva, N.V.; Podgornaya, O.I.; Voronin, A.P. Nucleolus-like body of mouse oocytes contains lamin A and B and TRF2 but not actin and topo II. Mol. Cytogenet. 2016, 9, 50. [Google Scholar] [CrossRef]
- Bogolyubov, D.S.; Bogolyubova, I.O. Unusual biomolecular condensates containing nucleoporins or lamin in the germinal vesicle of the common frog. Cell Tissue Biol. 2023, 17, 492–502. [Google Scholar] [CrossRef]
- Parfenov, V.N. The karyosphere during late oogenesis in Rana ridibunda. Eur. J. Cell Biol. 1979, 19, 102–108. [Google Scholar]
- Segura-Totten, M.; Wilson, K.L. BAF: Roles in chromatin, nuclear structure and retrovirus integration. Trends Cell Biol. 2004, 14, 261–266. [Google Scholar] [CrossRef]
- Fiil, A.; Moens, P. The development, structure and function of modified synaptonemal complexes in mosqito oocytes. Chromosoma 1973, 41, 37–62. [Google Scholar] [CrossRef]
- Kim, E.; Barth, R.; Dekker, C. Looping the genome with SMC complexes. Annu. Rev. Biochem. 2023, 92, 15–41. [Google Scholar] [CrossRef]





















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogolyubov, D.S.; Shabelnikov, S.V.; Travina, A.O.; Sulatsky, M.I.; Bogolyubova, I.O. Special Nuclear Structures in the Germinal Vesicle of the Common Frog with Emphasis on the So-Called Karyosphere Capsule. J. Dev. Biol. 2023, 11, 44. https://doi.org/10.3390/jdb11040044
Bogolyubov DS, Shabelnikov SV, Travina AO, Sulatsky MI, Bogolyubova IO. Special Nuclear Structures in the Germinal Vesicle of the Common Frog with Emphasis on the So-Called Karyosphere Capsule. Journal of Developmental Biology. 2023; 11(4):44. https://doi.org/10.3390/jdb11040044
Chicago/Turabian StyleBogolyubov, Dmitry S., Sergey V. Shabelnikov, Alexandra O. Travina, Maksim I. Sulatsky, and Irina O. Bogolyubova. 2023. "Special Nuclear Structures in the Germinal Vesicle of the Common Frog with Emphasis on the So-Called Karyosphere Capsule" Journal of Developmental Biology 11, no. 4: 44. https://doi.org/10.3390/jdb11040044