
Regulation and Function of FOXC1 in Osteoblasts
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Mice
2.2. Human Cells
2.3. Cell Lines
2.4. shGATA4
2.5. RNA and qPCR
2.6. Immunoprecipitation and Immunoblotting
2.7. Chromatin Immunoprecipitation (ChIP)
2.8. ChIP-reChIP
2.9. Immunohistochemistry (IHC)
2.10. Luciferase Assay
2.11. Cistrome Data Browser
2.12. FOXC1 Expression in Osteoporotic Patients
2.13. Statistical Analysis
3. Results
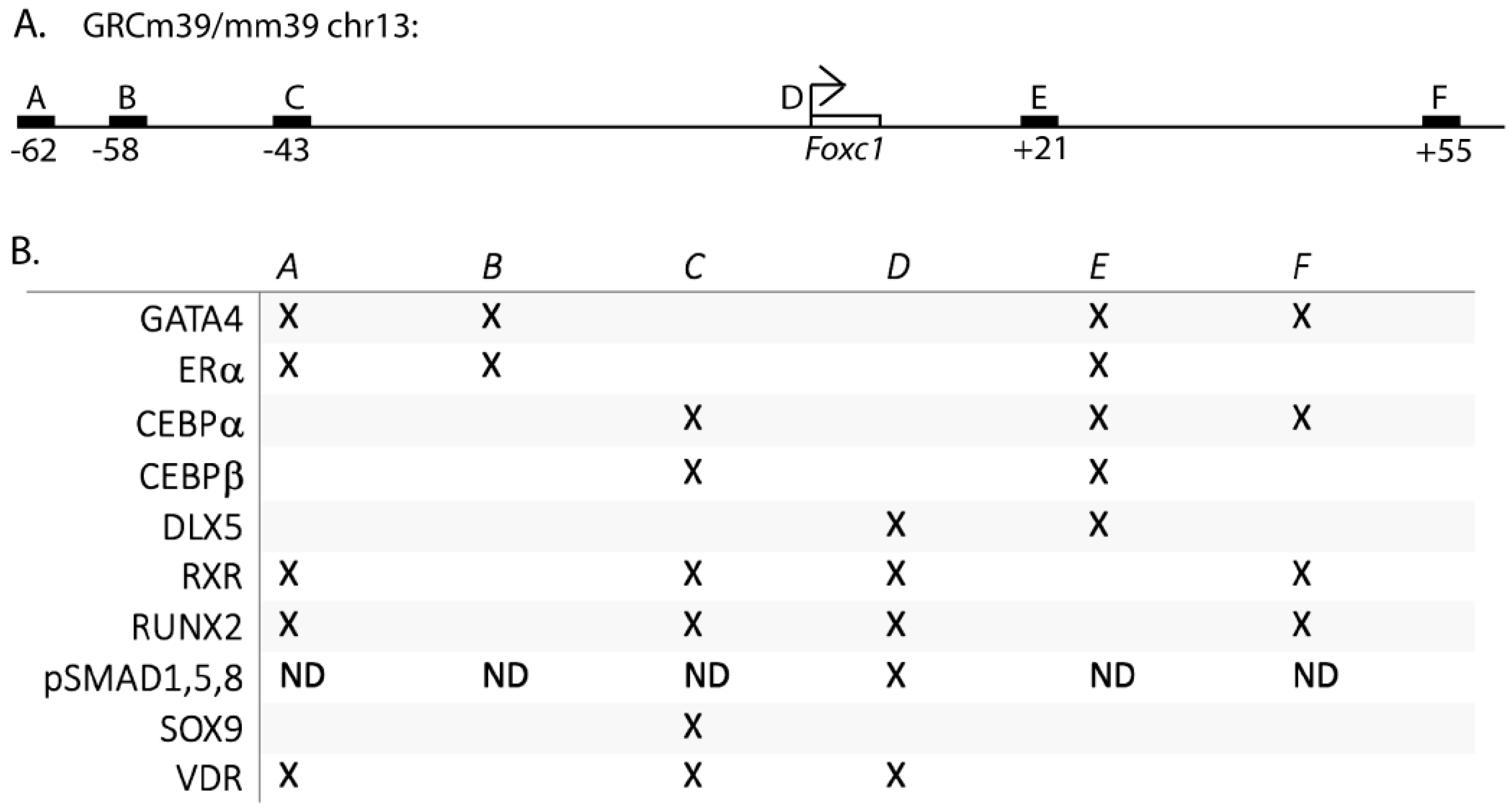
3.1. The Transcriptional Regulation of Foxc1
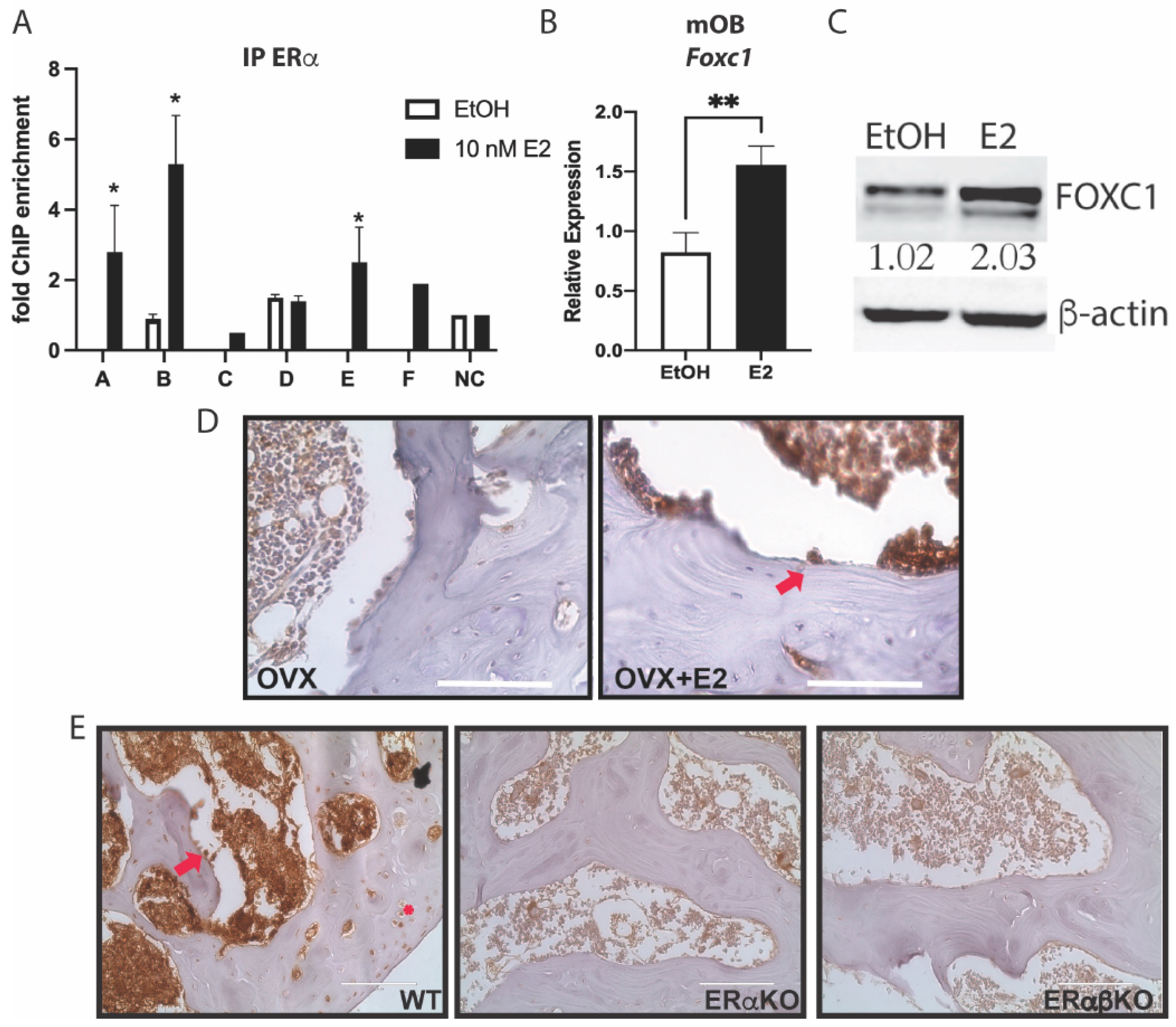
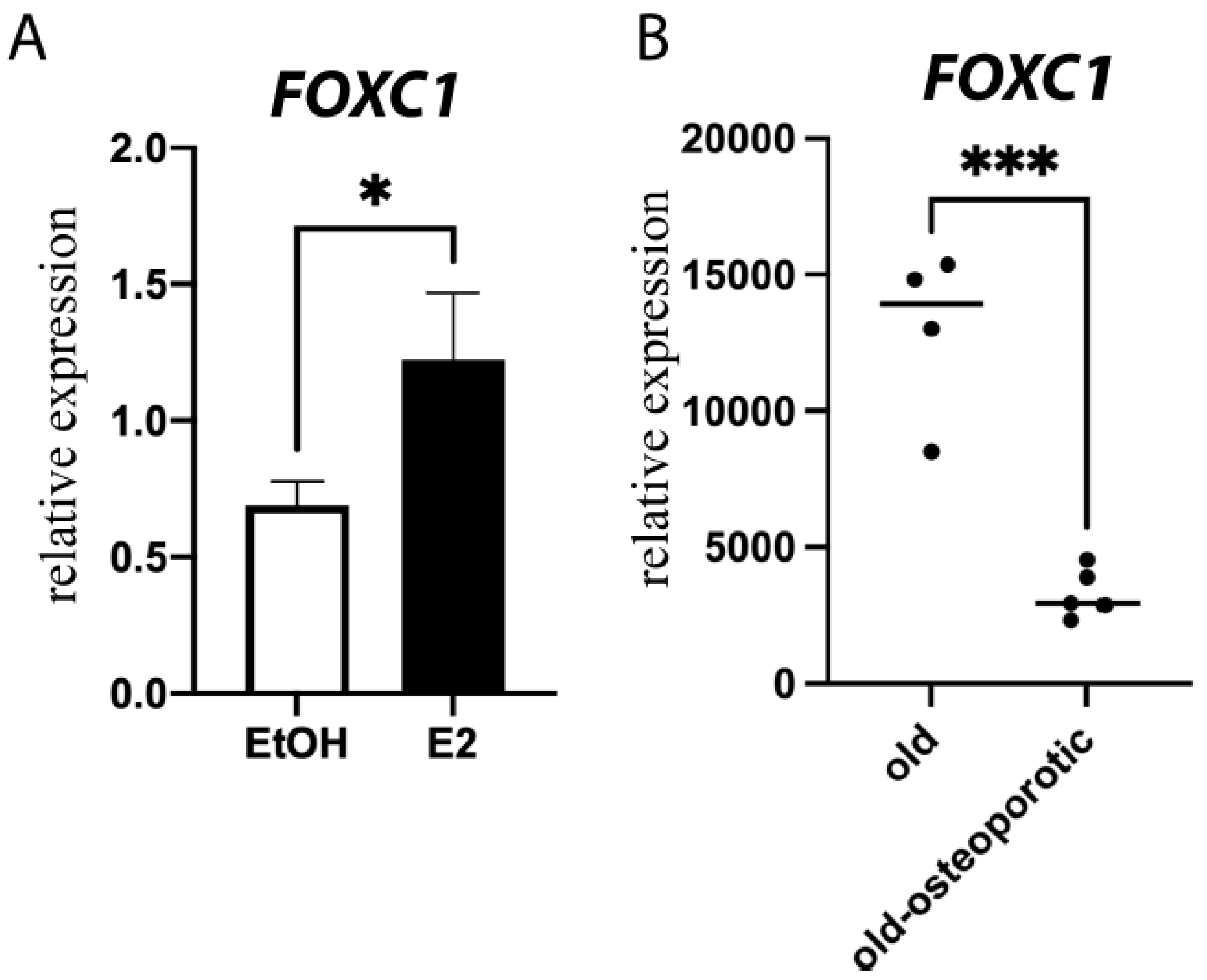
3.2. Estrogen Regulates Foxc1 in Osteoblasts
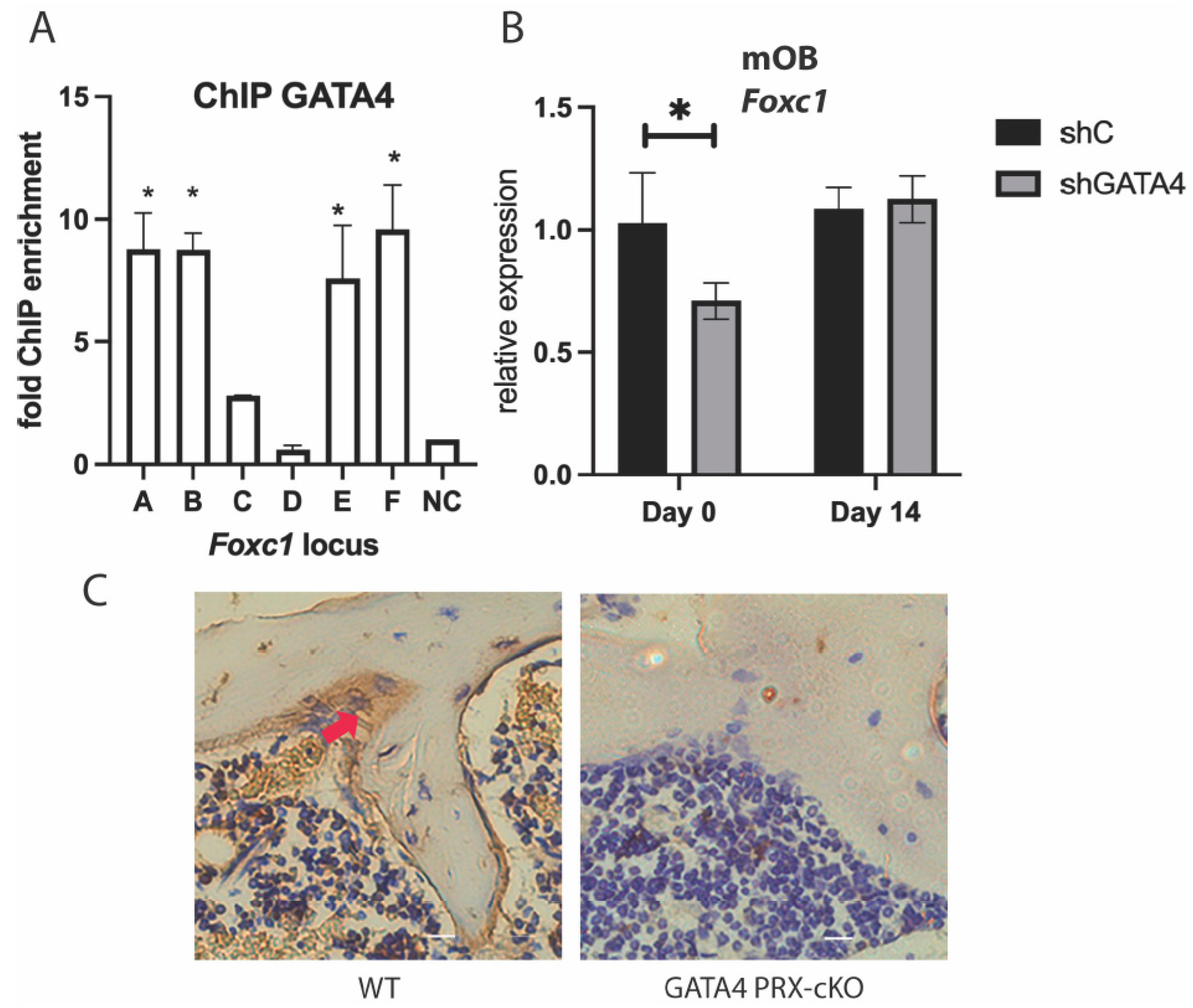
3.3. GATA4 Regulates Foxc1 Expression
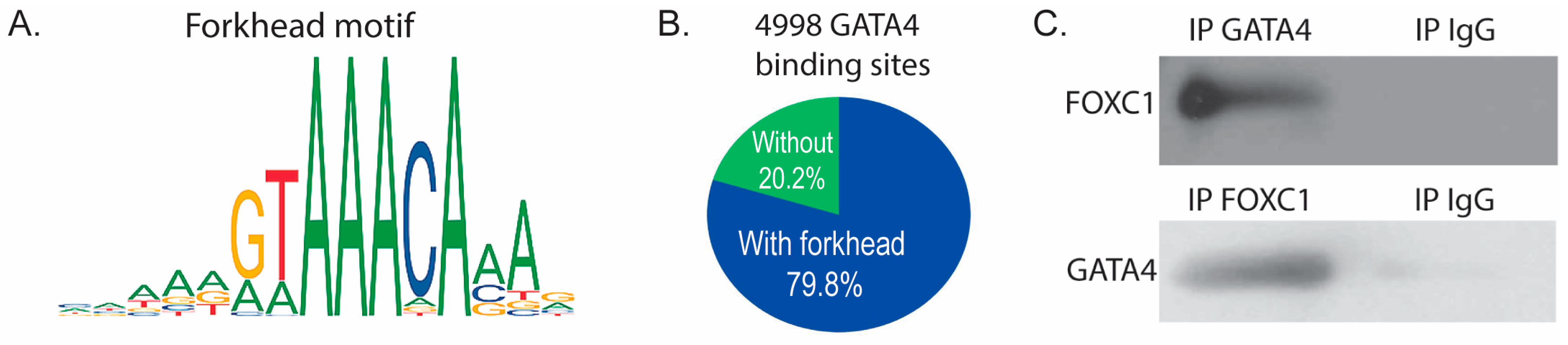
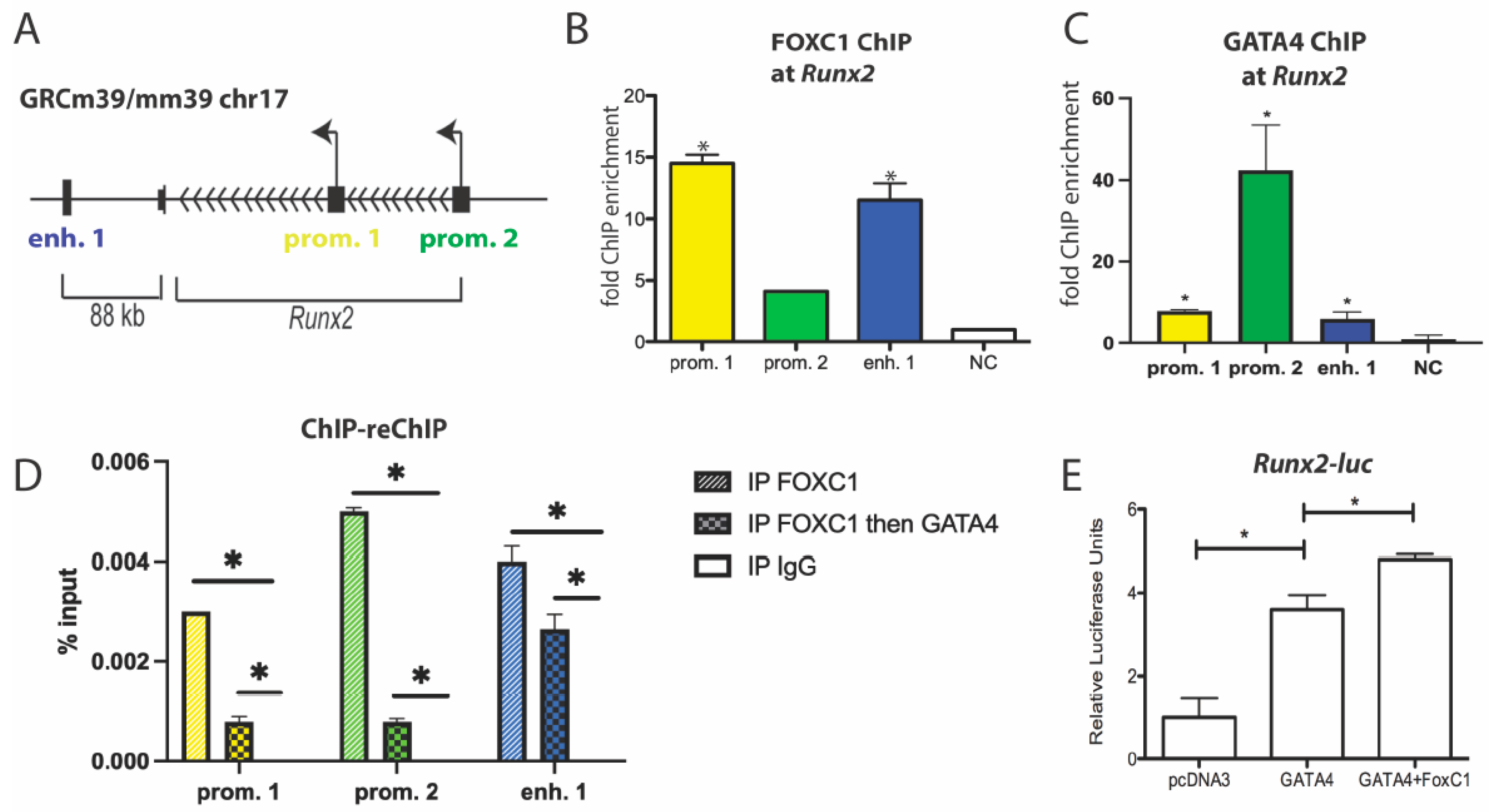
3.4. GATA4 and FOXC1 Interact in Osteoblasts to Regulate Osteoblast Differentiation
3.5. Clinical Relevance of FOXC1 and Osteoporosis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
| ChIP-seq | |
| ERα | GSE28918 |
| CEBPα | GSM2104127 |
| CEBPβ | GSM2104228 |
| DLX5 | GSM1976254 |
| RXR | GSM2104118 |
| RUNX2 | GSM2104159 |
| SOX9 | GSM826703 |
| VDR | GSM2104116 |
Conflicts of Interest
References
- Khalid, A.B.; Krum, S.A. Estrogen receptors alpha and beta in bone. Bone 2016, 87, 130–135. [Google Scholar] [CrossRef]
- Krum, S.A. Direct transcriptional targets of sex steroid hormones in bone. J. Cell. Biochem. 2011, 112, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Guemes, M.; Garcia, A.J.; Rigueur, D.; Runke, S.; Wang, W.; Zhao, G.; Mayorga, V.H.; Atti, E.; Tetradis, S.; Peault, B.; et al. GATA4 is essential for bone mineralization via ERalpha and TGFbeta/BMP pathways. J. Bone Miner. Res. 2014, 29, 2676–2687. [Google Scholar] [CrossRef]
- Patient, R.K.; McGhee, J.D. The GATA family (vertebrates and invertebrates). Curr. Opin. Genet. Dev. 2002, 12, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.B.; Slayden, A.V.; Kumpati, J.; Perry, C.D.; Lillo, M.A.; Arroyo, S.R.; Miranda-Carboni, G.; Krum, S.A. GATA4 directly regulates Runx2 expression and osteoblast differentiation. JBMR Plus 2018, 2, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Carboni, G.A.; Guemes, M.; Bailey, S.; Anaya, E.; Corselli, M.; Peault, B.; Krum, S.A. GATA4 regulates estrogen receptor-alpha-mediated osteoblast transcription. Mol. Endocrinol. 2011, 25, 1126–1136. [Google Scholar] [CrossRef]
- Khalid, A.B.; Pence, J.; Suthon, S.; Lin, J.; Miranda-Carboni, G.A.; Krum, S.A. GATA4 regulates mesenchymal stem cells via direct transcriptional regulation of the WNT signalosome. Bone 2021, 144, 115819. [Google Scholar] [CrossRef]
- Khalid, A.B.; Slayden, A.V.; Kumpati, J.; Perry, C.D.; Berryhill, S.B.; Crawford, J.A.; Fatima, I.; Morselli, M.; Pellegrini, M.; Miranda-Carboni, G.A.; et al. GATA4 represses RANKL in osteoblasts via multiple long-range enhancers to regulate osteoclast differentiation. Bone 2018, 116, 78–86. [Google Scholar] [CrossRef]
- Zaret, K.S.; Carroll, J.S. Pioneer transcription factors: Establishing competence for gene expression. Genes Dev. 2011, 25, 2227–2241. [Google Scholar] [CrossRef]
- Zhou, P.; Zhang, Y.; Sethi, I.; Ye, L.; Trembley, M.A.; Cao, Y.; Akerberg, B.N.; Xiao, F.; Zhang, X.; Li, K.; et al. GATA4 Regulates Developing Endocardium Through Interaction With ETS1. Circ. Res. 2022, 131, e152–e168. [Google Scholar] [CrossRef]
- Cirillo, L.A.; Zaret, K.S. An early developmental transcription factor complex that is more stable on nucleosome core particles than on free DNA. Mol. Cell 1999, 4, 961–969. [Google Scholar] [CrossRef]
- Krum, S.A.; Miranda-Carboni, G.A.; Lupien, M.; Eeckhoute, J.; Carroll, J.S.; Brown, M. Unique ERα cistromes control cell type-specific gene regulation. Mol. Endocrinol. 2008, 22, 2393–2406. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.C.; Carpenter, C.; Nebert, D.W.; Vasiliou, V. Update of human and mouse forkhead box (FOX) gene families. Hum. Genom. 2010, 4, 345–352. [Google Scholar] [CrossRef]
- Gokce, G.; Oren, N.C.; Ozgonul, C. Axenfeld-Rieger syndrome associated with severe maxillofacial and skeletal anomalies. J. Oral Maxillofac. Pathol. JOMFP 2015, 19, 109. [Google Scholar] [CrossRef]
- Machida, A.; Okuhara, S.; Harada, K.; Iseki, S. Difference in apical and basal growth of the frontal bone primordium in Foxc1ch/ch mice. Congenit. Anom. 2014, 54, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.K.; Lass, J.H.; Chakravarti, A. Pleiotropic skeletal and ocular phenotypes of the mouse mutation congenital hydrocephalus (ch/Mf1) arise from a winged helix/forkhead transcriptionfactor gene. Hum. Mol. Genet. 1999, 8, 625–637. [Google Scholar] [CrossRef]
- Almubarak, A.; Lavy, R.; Srnic, N.; Hu, Y.; Maripuri, D.P.; Kume, T.; Berry, F.B. Loss of Foxc1 and Foxc2 function in chondroprogenitor cells disrupts endochondral ossification. J. Biol. Chem. 2021, 297, 101020. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, N.; Li, H.; Wang, M.; Shen, H.; Si, J.; Shen, G. The Transcription Factor Foxc1 Promotes Osteogenesis by Directly Regulating Runx2 in Response of Intermittent Parathyroid Hormone (1-34) Treatment. Front. Pharmacol. 2020, 11, 592. [Google Scholar] [CrossRef]
- Mirzayans, F.; Lavy, R.; Penner-Chea, J.; Berry, F.B. Initiation of early osteoblast differentiation events through the direct transcriptional regulation of Msx2 by FOXC1. PLoS ONE 2012, 7, e49095. [Google Scholar] [CrossRef]
- He, A.; Shen, X.; Ma, Q.; Cao, J.; von Gise, A.; Zhou, P.; Wang, G.; Marquez, V.E.; Orkin, S.H.; Pu, W.T. PRC2 directly methylates GATA4 and represses its transcriptional activity. Genes Dev. 2012, 26, 37–42. [Google Scholar] [CrossRef]
- Dupont, S.; Krust, A.; Gansmuller, A.; Dierich, A.; Chambon, P.; Mark, M. Effect of single and compound knockouts of estrogen receptors alpha (ERalpha) and beta (ERbeta) on mouse reproductive phenotypes. Development 2000, 127, 4277–4291. [Google Scholar] [CrossRef]
- Krum, S.A.; Miranda-Carboni, G.A.; Hauschka, P.V.; Carroll, J.S.; Lane, T.F.; Freedman, L.P.; Brown, M. Estrogen protects bone by inducing Fas ligand in osteoblasts to regulate osteoclast survival. EMBO J. 2008, 27, 535–545. [Google Scholar] [CrossRef]
- Suthon, S.; Lin, J.; Perkins, R.S.; Crockarell, J.R., Jr.; Miranda-Carboni, G.A.; Krum, S.A. Estrogen receptor alpha and NFATc1 bind to a bone mineral density-associated SNP to repress WNT5B in osteoblasts. Am. J. Hum. Genet. 2022, 109, 97–115. [Google Scholar] [CrossRef]
- Monroe, D.G.; Getz, B.J.; Johnsen, S.A.; Riggs, B.L.; Khosla, S.; Spelsberg, T.C. Estrogen receptor isoform-specific regulation of endogenous gene expression in human osteoblastic cell lines expressing either ERalpha or ERbeta. J. Cell. Biochem. 2003, 90, 315–326. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Fedchenko, N.; Reifenrath, J. Different approaches for interpretation and reporting of immunohistochemistry analysis results in the bone tissue—A review. Diagn. Pathol. 2014, 9, 221. [Google Scholar] [CrossRef]
- Zhang, Y.; Hassan, M.Q.; Xie, R.L.; Hawse, J.R.; Spelsberg, T.C.; Montecino, M.; Stein, J.L.; Lian, J.B.; van Wijnen, A.J.; Stein, G.S. Co-stimulation of the bone-related Runx2 P1 promoter in mesenchymal cells by SP1 and ETS transcription factors at polymorphic purine-rich DNA sequences (Y-repeats). J. Biol. Chem. 2009, 284, 3125–3135. [Google Scholar] [CrossRef] [PubMed]
- Morrisey, E.E.; Ip, H.S.; Lu, M.M.; Parmacek, M.S. GATA-6: A zinc finger transcription factor that is expressed in multiple cell lineages derived from lateral mesoderm. Dev. Biol. 1996, 177, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Qin, Q.; Wu, Q.; Sun, H.; Zheng, R.; Zang, C.; Zhu, M.; Wu, J.; Shi, X.; Taing, L.; et al. Cistrome Data Browser: A data portal for ChIP-Seq and chromatin accessibility data in human and mouse. Nucleic Acids Res. 2017, 45, D658–D662. [Google Scholar] [CrossRef]
- Zheng, R.; Wan, C.; Mei, S.; Qin, Q.; Wu, Q.; Sun, H.; Chen, C.H.; Brown, M.; Zhang, X.; Meyer, C.A.; et al. Cistrome Data Browser: Expanded datasets and new tools for gene regulatory analysis. Nucleic Acids Res. 2019, 47, D729–D735. [Google Scholar] [CrossRef]
- Benisch, P.; Schilling, T.; Klein-Hitpass, L.; Frey, S.P.; Seefried, L.; Raaijmakers, N.; Krug, M.; Regensburger, M.; Zeck, S.; Schinke, T.; et al. The transcriptional profile of mesenchymal stem cell populations in primary osteoporosis is distinct and shows overexpression of osteogenic inhibitors. PLoS ONE 2012, 7, e45142. [Google Scholar] [CrossRef]
- Suthon, S.; Perkins, R.S.; Lin, J.; Crockarell, J.R., Jr.; Miranda-Carboni, G.A.; Krum, S.A. GATA4 and estrogen receptor alpha bind at SNPs rs9921222 and rs10794639 to regulate AXIN1 expression in osteoblasts. Hum. Genet. 2022, 141, 1849–1861. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.; Mirzayans, F.; Berry, F. Foxc1 Expression in Early Osteogenic Differentiation Is Regulated by BMP4-SMAD Activity. J. Cell. Biochem. 2016, 117, 1707–1717. [Google Scholar] [CrossRef] [PubMed]
- McLeay, R.C.; Bailey, T.L. Motif Enrichment Analysis: A unified framework and an evaluation on ChIP data. BMC Bioinform. 2010, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Barutcu, A.R.; Tai, P.W.; Wu, H.; Gordon, J.A.; Whitfield, T.W.; Dobson, J.R.; Imbalzano, A.N.; Lian, J.B.; van Wijnen, A.J.; Stein, J.L.; et al. The bone-specific Runx2-P1 promoter displays conserved three-dimensional chromatin structure with the syntenic Supt3h promoter. Nucleic Acids Res. 2014, 42, 10360–10372. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Li, J.; Yao, W.; Wang, W.; Shi, B.; Yuan, F.; Dong, J.; Zhang, H. FOXC1 Negatively Regulates DKK1 Expression to Promote Gastric Cancer Cell Proliferation Through Activation of Wnt Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 662624. [Google Scholar] [CrossRef]
- Liu, K.; Ni, J.D.; Li, W.Z.; Pan, B.Q.; Yang, Y.T.; Xia, Q.; Huang, J. The Sp1/FOXC1/HOTTIP/LATS2/YAP/beta-catenin cascade promotes malignant and metastatic progression of osteosarcoma. Mol. Oncol. 2020, 14, 2678–2695. [Google Scholar] [CrossRef]
- Lam, E.W.; Brosens, J.J.; Gomes, A.R.; Koo, C.Y. Forkhead box proteins: Tuning forks for transcriptional harmony. Nat. Rev. Cancer 2013, 13, 482–495. [Google Scholar] [CrossRef]
- Lin, Y.H.; Jewell, B.E.; Gingold, J.; Lu, L.; Zhao, R.; Wang, L.L.; Lee, D.F. Osteosarcoma: Molecular Pathogenesis and iPSC Modeling. Trends Mol. Med. 2017, 23, 737–755. [Google Scholar] [CrossRef]
- Qiu, Z.; Dan, Z.; Che, X. FoxC1 promotes osteosarcoma cell proliferation and metastasis through the activation of EZH2. Int. J. Clin. Exp. Med. 2017, 10, 376–384. [Google Scholar]
- Won, K.Y.; Park, H.R.; Park, Y.K. Prognostic implication of immunohistochemical Runx2 expression in osteosarcoma. Tumori 2009, 95, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Sadikovic, B.; Thorner, P.; Chilton-Macneill, S.; Martin, J.W.; Cervigne, N.K.; Squire, J.; Zielenska, M. Expression analysis of genes associated with human osteosarcoma tumors shows correlation of RUNX2 overexpression with poor response to chemotherapy. BMC Cancer 2010, 10, 202. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suthon, S.; Lin, J.; Perkins, R.S.; Miranda-Carboni, G.A.; Krum, S.A. Regulation and Function of FOXC1 in Osteoblasts. J. Dev. Biol. 2023, 11, 38. https://doi.org/10.3390/jdb11030038
Suthon S, Lin J, Perkins RS, Miranda-Carboni GA, Krum SA. Regulation and Function of FOXC1 in Osteoblasts. Journal of Developmental Biology. 2023; 11(3):38. https://doi.org/10.3390/jdb11030038
Chicago/Turabian StyleSuthon, Sarocha, Jianjian Lin, Rachel S. Perkins, Gustavo A. Miranda-Carboni, and Susan A. Krum. 2023. "Regulation and Function of FOXC1 in Osteoblasts" Journal of Developmental Biology 11, no. 3: 38. https://doi.org/10.3390/jdb11030038