
Heme Oxygenase-1 Is Upregulated during Differentiation of Keratinocytes but Its Expression Is Dispensable for Cornification of Murine Epidermis
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Mice
2.3. Cell Culture
2.4. Histology, Immunohistochemistry and Immunofluorescence Analysis
2.5. Transglutaminase Activity Labeling on Tissue Sections
2.6. Western Blot Analysis
2.7. Quantitative RT-PCR Analysis
2.8. Single-Cell RNA-Sequencing (scRNA-seq) Data Analysis
3. Results
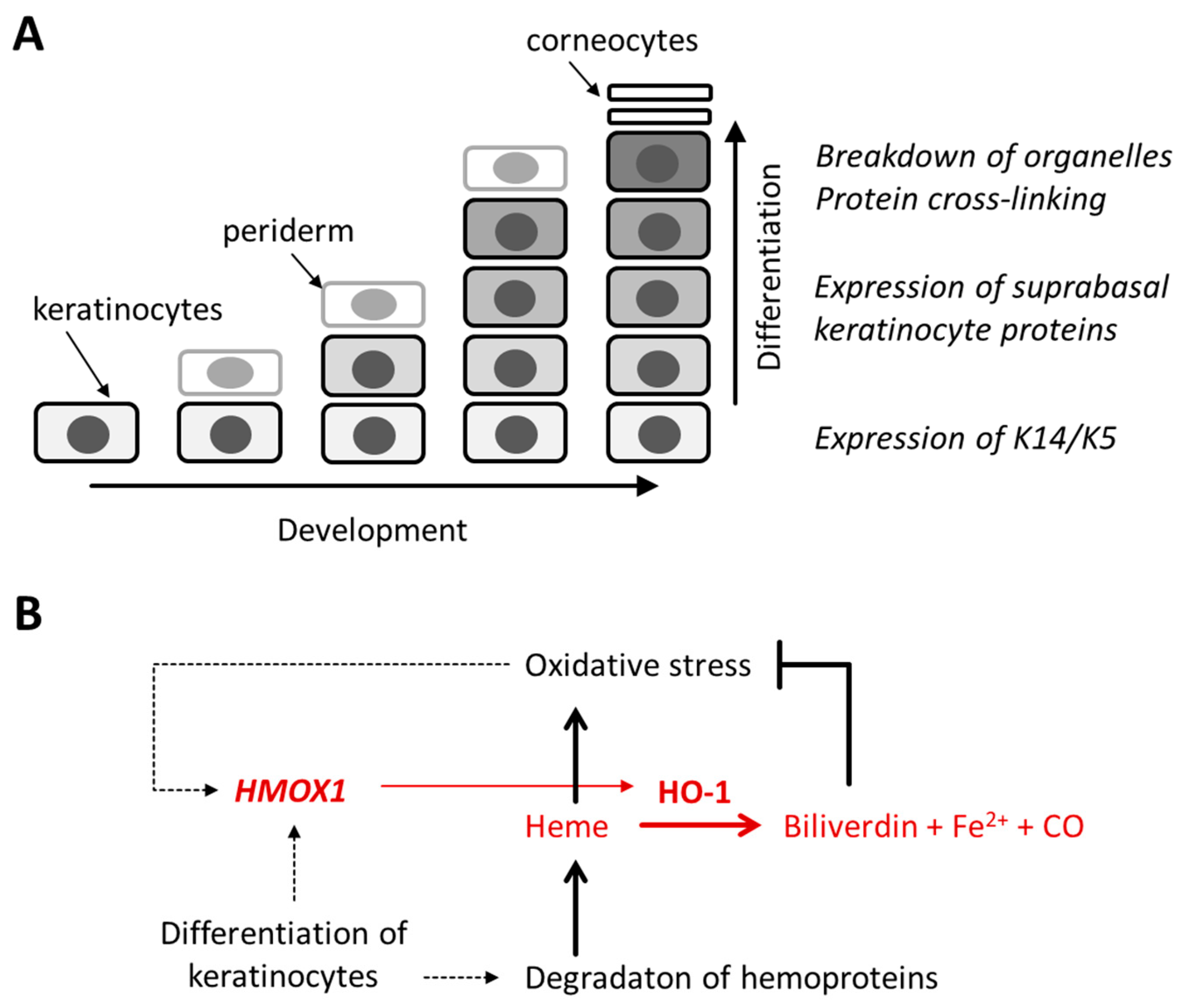
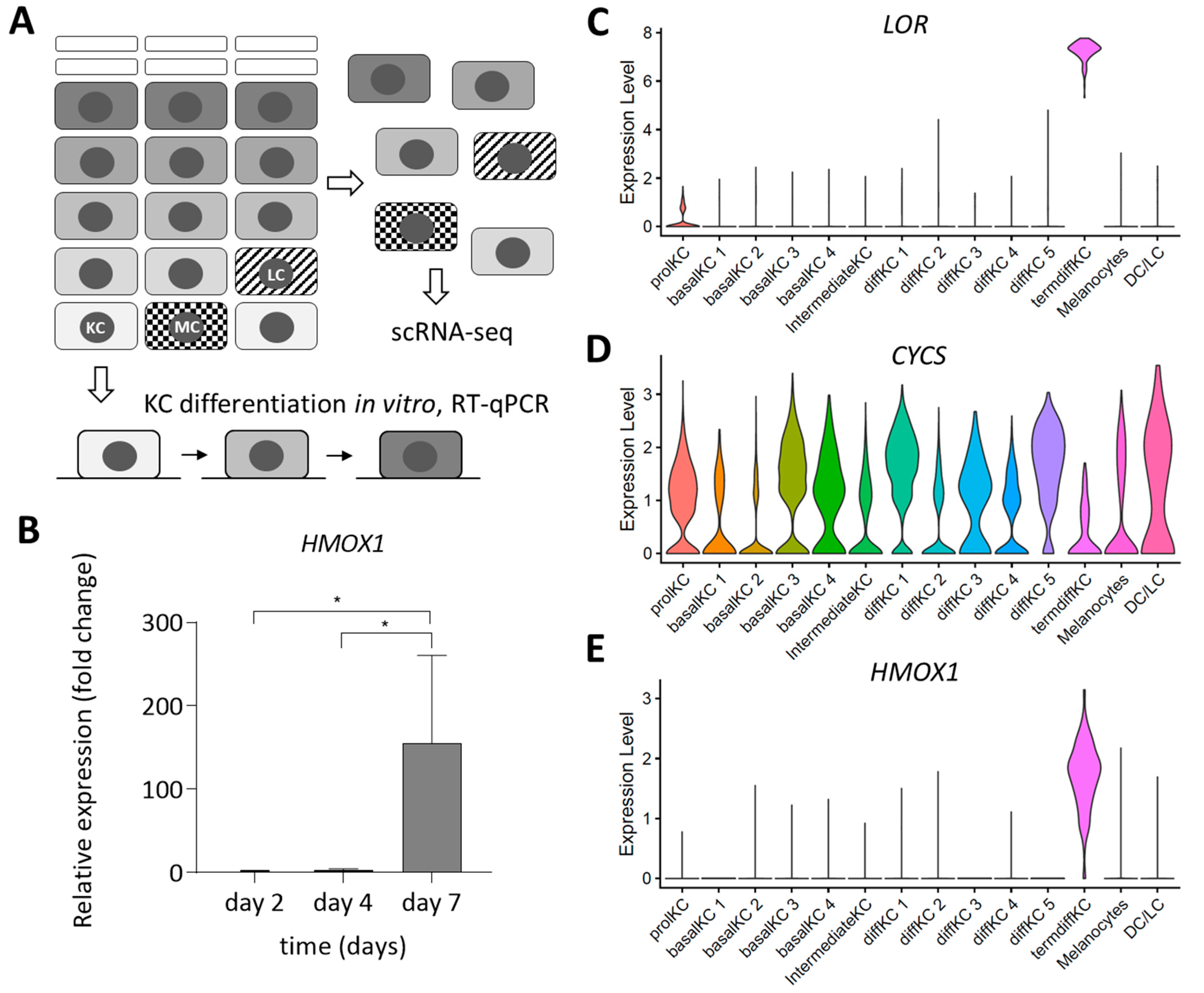
3.1. Expression of HMOX1 Is Upregulated during Terminal Differentiation of Epidermal Keratinocytes
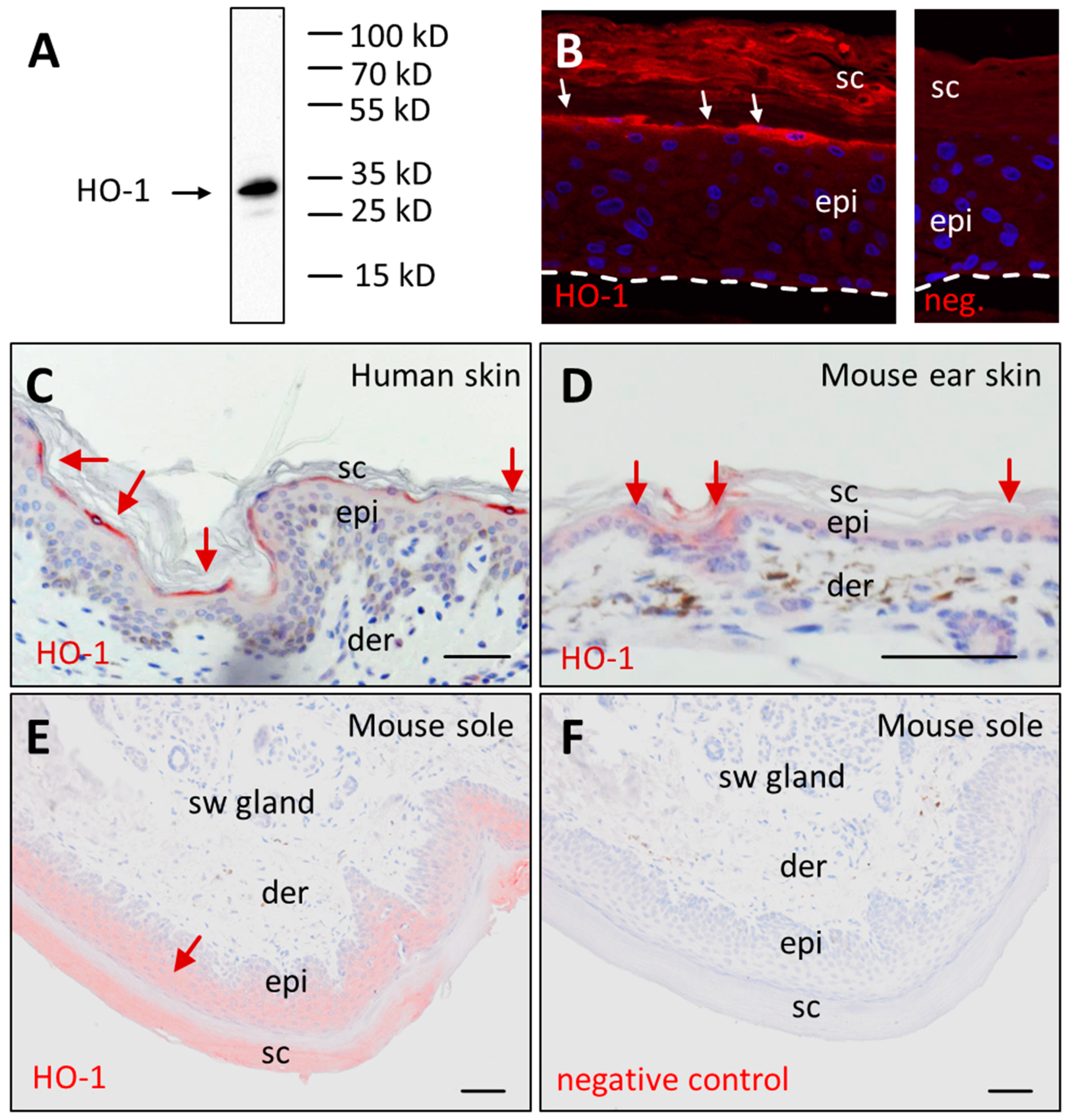
3.2. Immunolabeling Confirms Presence of HO-1 Protein in Terminally Differentiated Epidermal Keratinocytes
3.3. Development of Mouse Skin, Hair and Teeth Is Not Altered by Epithelial Deletion of HO-1
3.4. HO-1 Deletion Does Not Impair Terminal Differentiation under Standard Housing Conditions of Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Proksch, E.; Brandner, J.M.; Jensen, J.M. The skin: An indispensable barrier. Exp. Dermatol. 2008, 17, 1063–1072. [Google Scholar] [CrossRef]
- Matsui, T.; Amagai, M. Dissecting the formation, structure and barrier function of the stratum corneum. Int. Immunol. 2015, 27, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckhart, L.; Zeeuwen, P.L.J.M. The skin barrier: Epidermis vs environment. Exp. Dermatol. 2018, 27, 805–806. [Google Scholar] [CrossRef] [PubMed]
- Christophers, E.; Schröder, J.M. Evolution of innate defense in human skin. Exp. Dermatol. 2022, 31, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.C.; Fuchs, E. Building and maintaining the skin. Cold Spring Harb. Perspect. Biol. 2022, 14, a040840. [Google Scholar] [CrossRef]
- Eckhart, L.; Lippens, S.; Tschachler, E.; Declercq, W. Cell death by cornification. Biochim. Biophys. Acta 2013, 1833, 3471–3480. [Google Scholar] [CrossRef]
- Byrne, C.; Hardman, M.; Nield, K. Covering the limb-formation of the integument. J. Anat. 2003, 202, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Flora, P.; Ezhkova, E. Regulatory mechanisms governing epidermal stem cell function during development and homeostasis. Development 2020, 147, dev194100. [Google Scholar] [CrossRef] [PubMed]
- Hammond, N.L.; Dixon, J.; Dixon, M.J. Periderm: Life-cycle and function during orofacial and epidermal development. Semin Cell Dev. Biol. 2019, 91, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Morita, K.; Furuse, M.; Yoshida, Y.; Itoh, M.; Sasaki, H.; Tsukita, S.; Miyachi, Y. Molecular architecture of tight junctions of periderm differs from that of the maculae occludentes of epidermis. J. Investig. Dermatol. 2002, 118, 1073–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, H.; Langbein, L.; Reichelt, J.; Praetzel-Wunder, S.; Buchberger, M.; Ghannadan, M.; Tschachler, E.; Eckhart, L. Loss of keratin K2 expression causes aberrant aggregation of K10, hyperkeratosis, and inflammation. J. Investig. Dermatol. 2014, 134, 2579–2588. [Google Scholar] [CrossRef] [Green Version]
- Bickenbach, J.R.; Greer, J.M.; Bundman, D.S.; Rothnagel, J.A.; Roop, D.R. Loricrin expression is coordinated with other epidermal proteins and the appearance of lipid lamellar granules in development. J. Investig. Dermatol. 1995, 104, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Toulza, E.; Mattiuzzo, N.R.; Galliano, M.F.; Jonca, N.; Dossat, C.; Jacob, D.; de Daruvar, A.; Wincker, P.; Serre, G.; Guerrin, M. Large-scale identification of human genes implicated in epidermal barrier function. Genome Biol. 2007, 8, R107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardman, M.J.; Sisi, P.; Banbury, D.N.; Byrne, C. Patterned acquisition of skin barrier function during development. Development 1998, 125, 1541–1552. [Google Scholar] [CrossRef]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef]
- Ryter, S.W. Heme oxygenase-1: An anti-inflammatory effector in cardiovascular, lung, and related metabolic disorders. Antioxidants 2022, 11, 555. [Google Scholar] [CrossRef] [PubMed]
- Numata, I.; Okuyama, R.; Memezawa, A.; Ito, Y.; Takeda, K.; Furuyama, K.; Shibahara, S.; Aiba, S. Functional expression of heme oxygenase-1 in human differentiated epidermis and its regulation by cytokines. J. Investig. Dermatol. 2009, 129, 2594–2603. [Google Scholar] [CrossRef] [Green Version]
- Asano, M.; Yamasaki, K.; Yamauchi, T.; Terui, T.; Aiba, S. Epidermal iron metabolism for iron salvage. J. Dermatol. Sci. 2017, 87, 101–109. [Google Scholar] [CrossRef] [Green Version]
- McMahon, M.; Ding, S.; Acosta-Jimenez, L.P.; Frangova, T.G.; Henderson, C.J.; Wolf, C.R. Measuring in vivo responses to endogenous and exogenous oxidative stress using a novel haem oxygenase 1 reporter mouse. J. Physiol. 2018, 596, 105–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gozzelino, R.; Jeney, V.; Soares, M.P. Mechanisms of cell protection by heme oxygenase-1. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 323–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udayanga, K.G.; Miyata, H.; Yokoo, Y.; Qi, W.M.; Takahara, E.; Mantani, Y.; Yokoyama, T.; Hoshi, N.; Kitagawa, H. Immunohistochemical study of the apoptosis process in epidermal epithelial cells of rats under a physiological condition. Histol. Histopathol. 2011, 26, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Jais, A.; Einwallner, E.; Sharif, O.; Gossens, K.; Lu, T.T.; Soyal, S.M.; Medgyesi, D.; Neureiter, D.; Paier-Pourani, J.; Dalgaard, K.; et al. Heme oxygenase-1 drives metaflammation and insulin resistance in mouse and man. Cell 2014, 158, 25–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukseree, S.; Mildner, M.; Rossiter, H.; Pammer, J.; Zhang, C.F.; Watanapokasin, R.; Tschachler, E.; Eckhart, L. Autophagy in the thymic epithelium is dispensable for the development of self-tolerance in a novel mouse model. PLoS ONE 2012, 7, e38933. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, C.F.; Rossiter, H.; Eckhart, L.; König, U.; Karner, S.; Mildner, M.; Bochkov, V.N.; Tschachler, E.; Gruber, F. Autophagy is induced by UVA and promotes removal of oxidized phospholipids and protein aggregates in epidermal keratinocytes. J. Investig. Dermatol. 2013, 133, 1629–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, H.; Eckhart, L.; Mildner, M.; Jaeger, K.; Buchberger, M.; Ghannadan, M.; Tschachler, E. DNase1L2 degrades nuclear DNA during corneocyte formation. J. Investig. Dermatol. 2007, 127, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Lachner, J.; Mlitz, V.; Tschachler, E.; Eckhart, L. Epidermal cornification is preceded by the expression of a keratinocyte-specific set of pyroptosis-related genes. Sci. Rep. 2017, 7, 17446. [Google Scholar] [CrossRef] [Green Version]
- Sukseree, S.; Rossiter, H.; Mildner, M.; Pammer, J.; Buchberger, M.; Gruber, F.; Watanapokasin, R.; Tschachler, E.; Eckhart, L. Targeted deletion of Atg5 reveals differential roles of autophagy in keratin K5-expressing epithelia. Biochem. Biophys. Res. Commun. 2013, 430, 689–694. [Google Scholar] [CrossRef]
- Sachslehner, A.P.; Surbek, M.; Lachner, J.; Paudel, S.; Eckhart, L. Identification of chicken transglutaminase 1 and in situ localization of transglutaminase activity in avian skin and esophagus. Genes 2021, 12, 1565. [Google Scholar] [CrossRef]
- Kalinina, P.; Vorstandlechner, V.; Buchberger, M.; Eckhart, L.; Lengauer, B.; Golabi, B.; Laggner, M.; Hiess, M.; Sterniczky, B.; Födinger, D.; et al. The whey acidic protein WFDC12 is specifically expressed in terminally differentiated keratinocytes and regulates epidermal serine protease activity. J. Investig. Dermatol. 2021, 141, 1198–1206. [Google Scholar] [CrossRef]
- Vorstandlechner, V.; Laggner, M.; Kalinina, P.; Haslik, W.; Radtke, C.; Shaw, L.; Lichtenberger, B.M.; Tschachler, E.; Ankersmit, H.J.; Mildner, M. Deciphering the functional heterogeneity of skin fibroblasts using single-cell RNA sequencing. FASEB J. 2020, 34, 3677–3692. [Google Scholar] [CrossRef]
- Sukseree, S.; Schwarze, U.Y.; Gruber, R.; Gruber, F.; Quiles Del Rey, M.; Mancias, J.D.; Bartlett, J.D.; Tschachler, E.; Eckhart, L. ATG7 is essential for secretion of iron from ameloblasts and normal growth of murine incisors during aging. Autophagy 2020, 16, 1851–1857. [Google Scholar] [CrossRef]
- Chavanas, S.; Garner, C.; Bodemer, C.; Ali, M.; Teillac, D.H.; Wilkinson, J.; Bonafé, J.L.; Paradisi, M.; Kelsell, D.P.; Ansai, S.; et al. Localization of the Netherton syndrome gene to chromosome 5q32, by linkage analysis and homozygosity mapping. Am. J. Hum. Genet. 2000, 66, 914–921. [Google Scholar] [CrossRef] [Green Version]
- Kelsell, D.P.; Norgett, E.E.; Unsworth, H.; The, M.T.; Cullup, T.; Mein, C.A.; Dopping-Hepenstal, P.J.; Dale, B.A.; Tadini, G.; Fleckman, P.; et al. Mutations in ABCA12 underlie the severe congenital skin disease harlequin ichthyosis. Am. J. Hum. Genet. 2005, 76, 794–803. [Google Scholar] [CrossRef] [Green Version]
- Koch, P.J.; de Viragh, P.A.; Scharer, E.; Bundman, D.; Longley, M.A.; Bickenbach, J.; Kawachi, Y.; Suga, Y.; Zhou, Z.; Huber, M.; et al. Lessons from loricrin-deficient mice: Compensatory mechanisms maintaining skin barrier function in the absence of a major cornified envelope protein. J. Cell Biol. 2000, 151, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Spazierer, D.; Fuchs, P.; Reipert, S.; Fischer, I.; Schmuth, M.; Lassmann, H.; Wiche, G. Epiplakin is dispensable for skin barrier function and for integrity of keratin network cytoarchitecture in simple and stratified epithelia. Mol. Cell Biol. 2006, 26, 559–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaRock, D.L.; Johnson, A.F.; Wilde, S.; Sands, J.S.; Monteiro, M.P.; LaRock, C.N. Group A streptococcus induces GSDMA-dependent pyroptosis in keratinocytes. Nature 2022, 605, 527–531. [Google Scholar] [CrossRef]
- Tanaka, S.; Mizushina, Y.; Kato, Y.; Tamura, M.; Shiroishi, T. Functional conservation of gsdma cluster genes specifically duplicated in the mouse genome. G3 2013, 3, 1843–1850. [Google Scholar] [CrossRef] [Green Version]
- de Guzman Strong, C.; Conlan, S.; Deming, C.B.; Cheng, J.; Sears, K.E.; Segre, J.A. A milieu of regulatory elements in the epidermal differentiation complex syntenic block: Implications for atopic dermatitis and psoriasis. Hum. Mol. Genet. 2010, 19, 1453–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, B.; Mlitz, V.; Fischer, H.; Tschachler, E.; Eckhart, L. Comparative genomics reveals conservation of filaggrin and loss of caspase-14 in dolphins. Exp. Dermatol. 2015, 24, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Holthaus, K.B.; Lachner, J.; Ebner, B.; Tschachler, E.; Eckhart, L. Gene duplications and gene loss in the epidermal differentiation complex during the evolutionary land-to-water transition of cetaceans. Sci. Rep. 2021, 11, 12334. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Luo, X.; Tang, J.; Xu, S.; Huang, K.; Wang, X.; Feng, T.; Que, T.; Jia, M.; Guo, X.; et al. High-quality genomes of pangolins: Insights into the molecular basis of scale formation and adaption to myrmecophagous diet. Mol. Biol. Evol. 2023, 40, msac262. [Google Scholar] [CrossRef]
- Fischer, H.; Rossiter, H.; Ghannadan, M.; Jaeger, K.; Barresi, C.; Declercq, W.; Tschachler, E.; Eckhart, L. Caspase-14 but not caspase-3 is processed during the development of fetal mouse epidermis. Differentiation 2005, 73, 406–413. [Google Scholar] [CrossRef]
- Ingraham, C.R.; Kinoshita, A.; Kondo, S.; Yang, B.; Sajan, S.; Trout, K.J.; Malik, M.I.; Dunnwald, M.; Goudy, S.L.; Lovett, M.; et al. Abnormal skin, limb and craniofacial morphogenesis in mice deficient for interferon regulatory factor 6 (Irf6). Nat. Genet. 2006, 38, 1335–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckhart, L.; Tschachler, E. Control of cell death-associated danger signals during cornification prevents autoinflammation of the skin. Exp. Dermatol. 2018, 27, 884–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, C.L.; Tokito, M.K.; Uppala, R.; Sarkar, M.K.; Gudjonsson, J.E.; Holzbaur, E.L.F. NIX initiates mitochondrial fragmentation via DRP1 to drive epidermal differentiation. Cell Rep. 2021, 34, 108689. [Google Scholar] [CrossRef]
- Ishitsuka, Y.; Roop, D.R. The epidermis: Redox governor of health and diseases. Antioxidants 2021, 11, 47. [Google Scholar] [CrossRef]
- Applegate, L.A.; Noël, A.; Vile, G.; Frenk, E.; Tyrrell, R.M. Two genes contribute to different extents to the heme oxygenase enzyme activity measured in cultured human skin fibroblasts and keratinocytes: Implications for protection against oxidant stress. Photochem. Photobiol. 1995, 61, 285–291. [Google Scholar] [CrossRef]
- Tyrrell, R.M. Solar ultraviolet A radiation: An oxidizing skin carcinogen that activates heme oxygenase-1. Antioxid. Redox. Signal. 2004, 6, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Grochot-Przeczek, A.; Lach, R.; Mis, J.; Skrzypek, K.; Gozdecka, M.; Sroczynska, P.; Dubiel, M.; Rutkowski, A.; Kozakowska, M.; Zagorska, A.; et al. Heme oxygenase-1 accelerates cutaneous wound healing in mice. PLoS ONE 2009, 4, e5803. [Google Scholar] [CrossRef] [Green Version]
- Leman, G.; Pavel, P.; Hermann, M.; Crumrine, D.; Elias, P.M.; Minzaghi, D.; Goudounèche, D.; Roshardt Prieto, N.M.; Cavinato, M.; Wanner, A.; et al. Mitochondrial activity is upregulated in nonlesional atopic dermatitis and amenable to therapeutic intervention. J. Investig. Dermatol. 2022, 142, 2623–2634.e12. [Google Scholar] [CrossRef]
- Kirino, M.; Kirino, Y.; Takeno, M.; Nagashima, Y.; Takahashi, K.; Kobayashi, M.; Murakami, S.; Hirasawa, T.; Ueda, A.; Aihara, M.; et al. Heme oxygenase 1 attenuates the development of atopic dermatitis-like lesions in mice: Implications for human disease. J. Allergy Clin. Immunol. 2008, 122, 290–297, 297.e1-8. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.Y.R.; Tan, C.L.; Chin, T.; Morenc, M.; Ho, C.Y.; Rovito, H.A.; Quek, L.S.; Soon, A.L.; Lim, J.S.Y.; Dreesen, O.; et al. Nicotinamide prevents UVB- and oxidative stress‒induced photoaging in human primary keratinocytes. J. Investig. Dermatol. 2022, 142, 1670–1681.e12. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surbek, M.; Sukseree, S.; Sachslehner, A.P.; Copic, D.; Golabi, B.; Nagelreiter, I.M.; Tschachler, E.; Eckhart, L. Heme Oxygenase-1 Is Upregulated during Differentiation of Keratinocytes but Its Expression Is Dispensable for Cornification of Murine Epidermis. J. Dev. Biol. 2023, 11, 12. https://doi.org/10.3390/jdb11010012
Surbek M, Sukseree S, Sachslehner AP, Copic D, Golabi B, Nagelreiter IM, Tschachler E, Eckhart L. Heme Oxygenase-1 Is Upregulated during Differentiation of Keratinocytes but Its Expression Is Dispensable for Cornification of Murine Epidermis. Journal of Developmental Biology. 2023; 11(1):12. https://doi.org/10.3390/jdb11010012
Chicago/Turabian StyleSurbek, Marta, Supawadee Sukseree, Attila Placido Sachslehner, Dragan Copic, Bahar Golabi, Ionela Mariana Nagelreiter, Erwin Tschachler, and Leopold Eckhart. 2023. "Heme Oxygenase-1 Is Upregulated during Differentiation of Keratinocytes but Its Expression Is Dispensable for Cornification of Murine Epidermis" Journal of Developmental Biology 11, no. 1: 12. https://doi.org/10.3390/jdb11010012