
In Vitro Comparison of Sex-Specific Splicing Efficiencies of fem Pre-mRNA under Monoallelic and Heteroallelic Conditions of csd, a Master Sex-Determining Gene in the Honeybee
Abstract
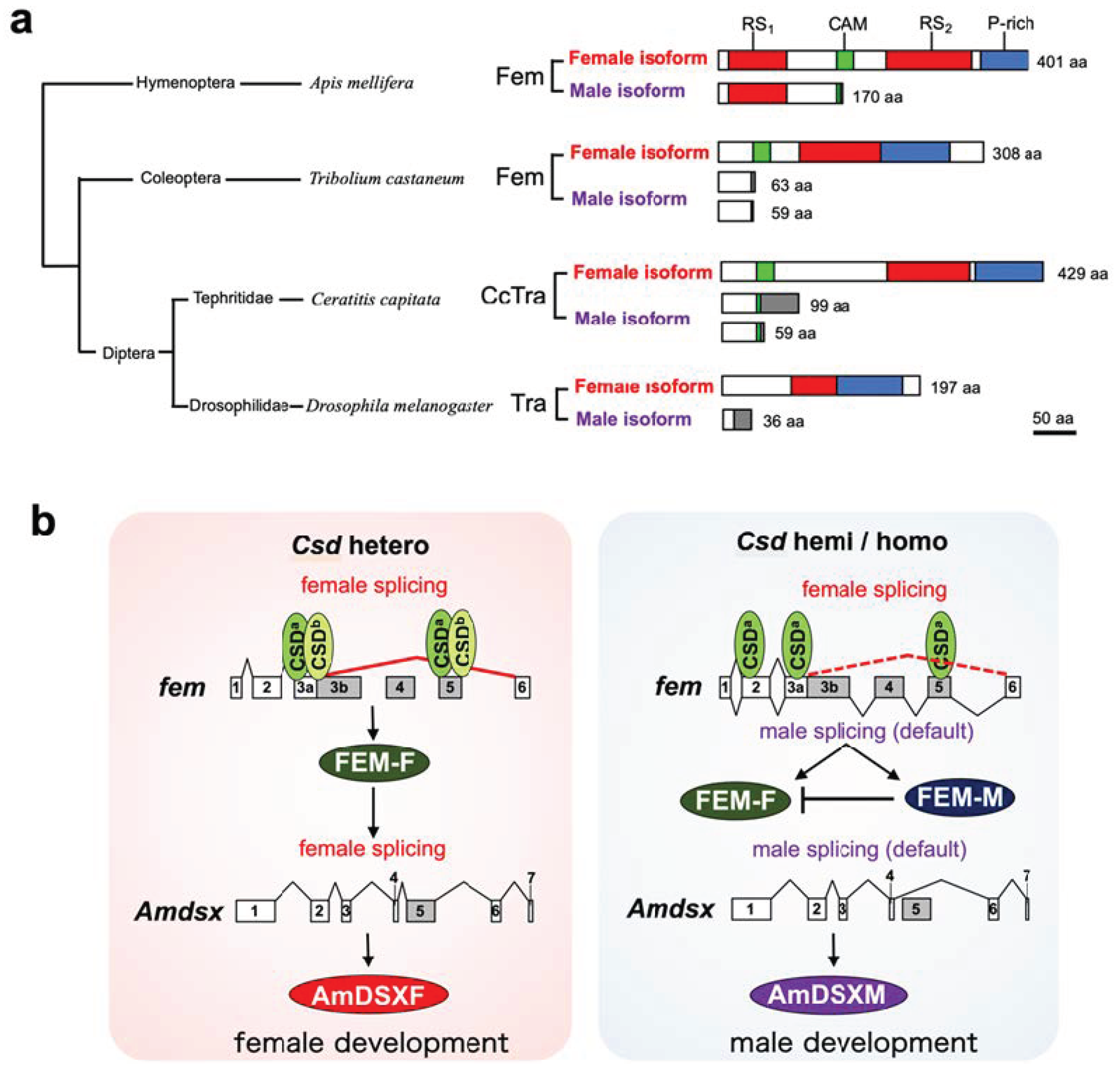
:1. Introduction
2. Materials and Methods
2.1. Genomic DNA Extraction
2.2. RNA Extraction and RT-PCR
2.3. Plasmid Construction
2.4. DNA Transfection of Cells
2.5. Quantitative RT-PCR
2.6. RNA Immunoprecipitation qPCR (RIP-qPCR) Analysis
3. Results
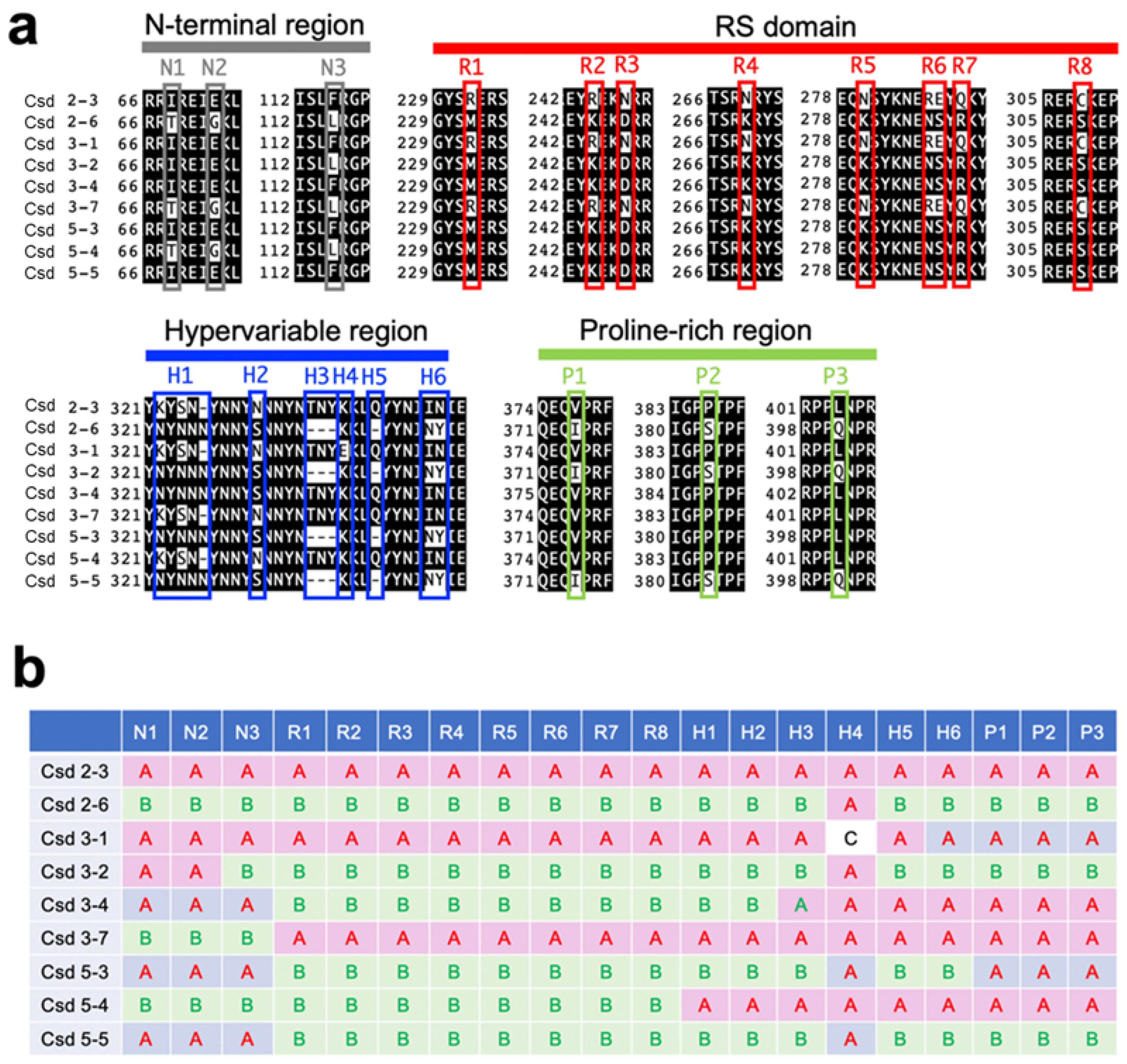
3.1. Acquisition of csd Alleles
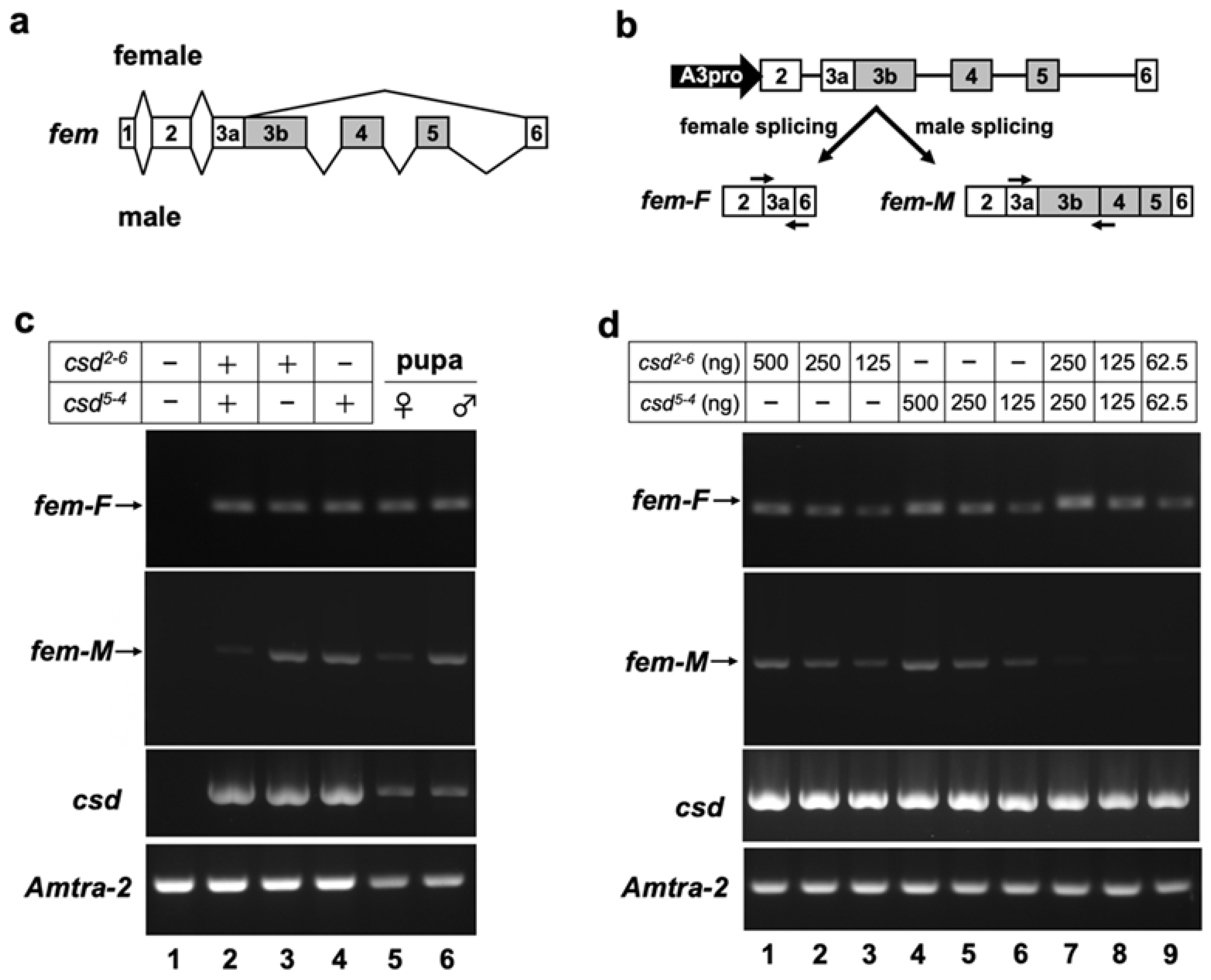
3.2. Heterogeneous Expression of Csd Proteins Induces Female-Specific Splicing of the fem Minigene
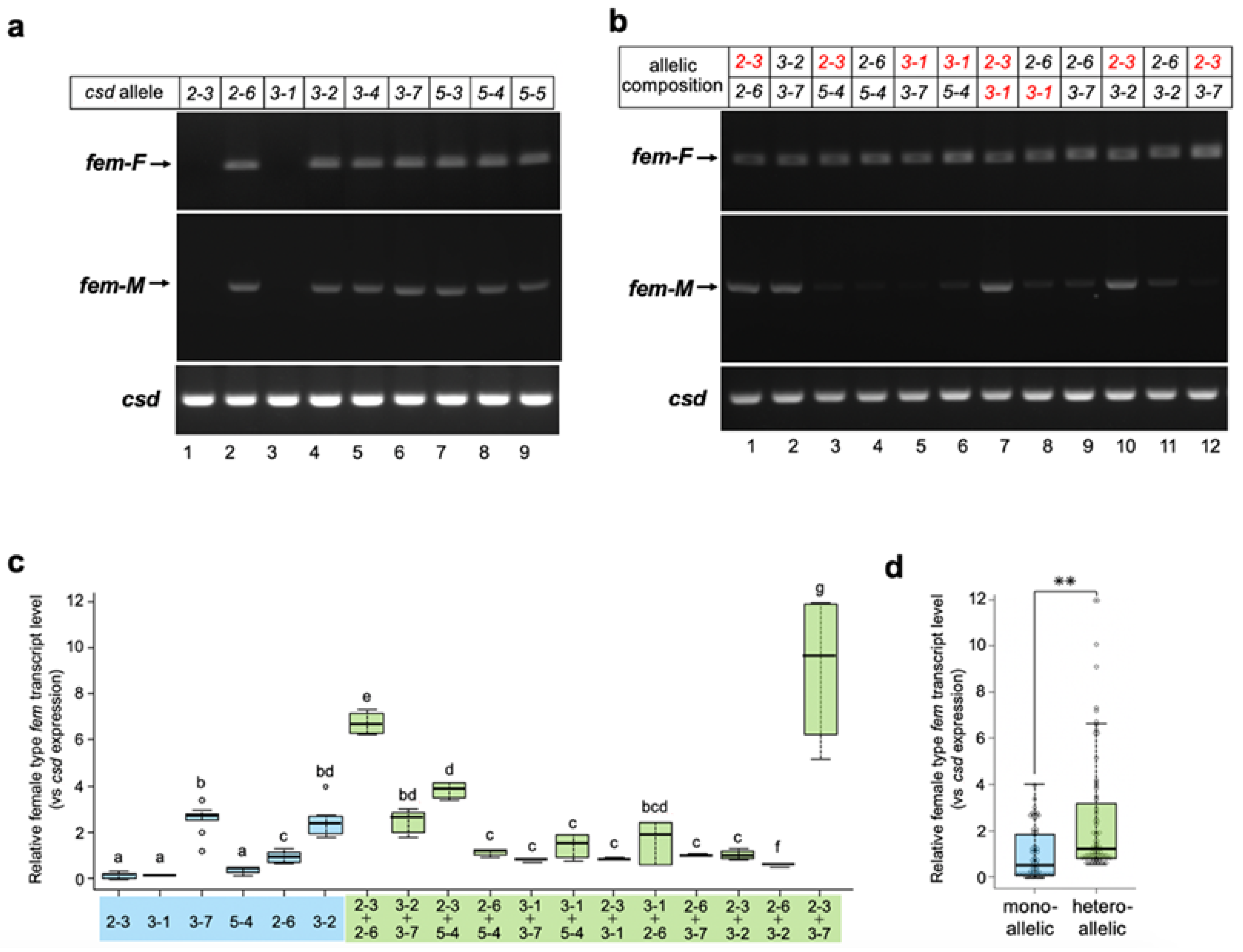
3.3. Csd Induces the Female Mode of Fem Splicing under Single-Allele Conditions
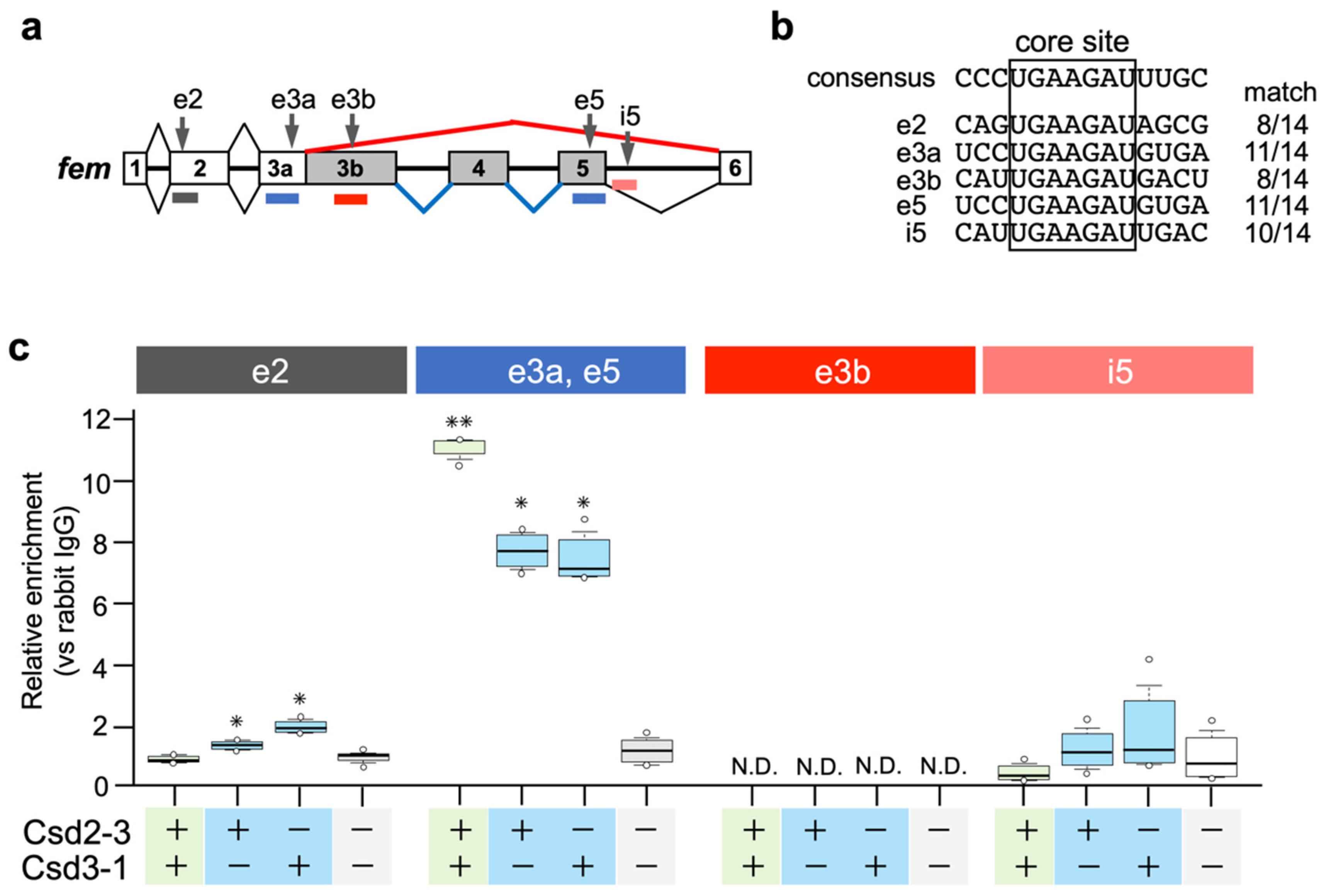
3.4. Csd Protein Specifically Enriches fem Pre-mRNA Even under Monoallelic Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kashimada, K.; Koopman, P. Sry: The master switch in mammalian sex determination. Development 2010, 137, 3921–3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridges, C.B. Triploid intersexes in Drosophila melanogaster. Science 1921, 16, 252–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, J.W.; Quintero, J.J. Indirect effects of ploidy suggest X chromosome dose, not the X:A ratio, signals sex in Drosophila. PLoS Biol. 2007, 5, e332. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Kozhina, E.; Mahadevaraju, S.; Yang, D.; Avila, F.W.; Erickson, J.W. Maternal Groucho and bHLH repressors amplify the dose-sensitive X chromosome signal in Drosophila sex determination. Dev. Biol. 2008, 323, 248–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, H. The role of the W-chromosome in the sex determination of Bombyx mori. Jpn. J. Genet. 1933, 8, 245–247. [Google Scholar]
- Kiuchi, T.; Koga, H.; Kawamoto, M.; Shoji, K.; Sakai, H.; Arai, Y.; Ishihara, G.; Kawaoka, S.; Sugano, S.; Shimada, T.; et al. A single female-specific piRNA is the primary determiner of sex in the silkworm. Nature 2014, 509, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Cook, M. Sex determination in the hymenoptera: A review of models and evidence. Heredity 1993, 71, 421–435. [Google Scholar] [CrossRef] [Green Version]
- Whiting, P.W. Multiple alleles in complementary sex determination of Habrobracon. Genetics 1943, 28, 365. [Google Scholar] [CrossRef]
- Whiting, P.W. Genetics of Habrobracon. Adv. Genet. 1961, 10, 295–348. [Google Scholar]
- Adams, J.; Rothman, E.D.; Kerr, W.E.; Paulino, Z.L. Estimation of the number of sex alleles and queen matings from diploid male frequencies in a population of Apis mellifera. Genetics 1977, 86, 583–596. [Google Scholar] [CrossRef]
- Naito, T.; Suzuki, H. Sex determination in the sawfly, Athalia rosae ruficornis (Hymenoptera): Occurrence of triploid males. J. Hered. 1991, 82, 101–104. [Google Scholar] [CrossRef]
- Beye, M.; Hasselmann, M.; Fondrk, M.K.; Page, R.E.; Omholt, S.W. The gene csd is the primary signal for sexual development in the honeybee and encodes an SR-type protein. Cell 2003, 114, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasselmann, M.; Gempe, T.; Schiøtt, M.; Nunes-Silva, C.G.; Otte, M.; Beye, M. Evidence for the evolutionary nascence of a novel sex determination pathway in honeybees. Nature 2008, 454, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Koch, V.; Nissen, I.; Schmitt, B.D.; Beye, M. Independent evolutionary origin of fem paralogous genes and complementary sex determination in hymenopteran insects. PLoS ONE 2014, 9, e91883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gempe, T.; Hasselmann, M.; Schiøtt, M.; Hause, G.; Otte, M.; Beye, M. Sex determination in honeybees: Two separate mechanisms induce and maintain the female pathway. PLoS Biol. 2009, 7, e1000222. [Google Scholar] [CrossRef] [Green Version]
- McAfee, A.; Pettis, J.S.; Tarpy, D.R.; Foster, L.J. Feminizer and doublesex knock-outs cause honey bees to switch sexes. PLoS Biol. 2019, 17, e30002562019. [Google Scholar] [CrossRef] [Green Version]
- Nissen, I.; Müller, M.; Beye, M. The Am-tra2 gene is an essential regulator of female splice regulation at two levels of the sex determination hierarchy of the honeybee. Genetics 2012, 192, 1015–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagamitsu, S.; Takase, D.; Aoki, F.; Suzuki, M.G. Identification of the Doublesex protein binding sites that activate expression of lozenge in the female genital disc in Drosophila melanogaster. Mech. Dev. 2017, 143, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S. A plaque assay and cloning of Bombyx mori nuclear polyhedrosis virus. J. Sericult. Sci. 1984, 53, 547–548. [Google Scholar]
- Suzuki, M.G.; Suzuki, K.; Aoki, F.; Ajimura, M. Effect of RNAi-mediated knockdown of the Bombyx mori transformer-2 gene on the sex-specific splicing of Bmdsx pre-mRNA. Int. J. Dev. Biol. 2012, 56, 693–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, A.; Aoki, F.; Suzuki, M.G. Conserved domains in Transformer protein act complementary to regulate sex-specific splicing of its own pre-mRNA. Sex Dev. 2018, 12, 189–190. [Google Scholar] [CrossRef] [PubMed]
- Yuzawa, T.; Matsuoka, M.; Sumitani, M.; Aoki, F.; Sezutsu, H.; Suzuki, M.G. Transgenic and knockout analyses of Masculinizer and doublesex illuminated the unique functions of doublesex in germ cell sexual development of the silkworm, Bombyx mori. BMC Dev. Biol. 2020, 20, 19. [Google Scholar] [CrossRef] [PubMed]
- Esnault, S.; Malter, J.S. Hyaluronic acid or TNF-alpha plus fibronectin triggers granulocyte macrophage-colony-stimulating factor mRNA stabilization in eosinophils yet engages differential intracellular pathways and mRNA binding proteins. J. Immunol. 2003, 171, 6780–6787. [Google Scholar] [CrossRef] [Green Version]
- Gempe, T.; Beye, M. Function and evolution of sex determination mechanisms, genes and pathways in insects. BioEssays 2011, 33, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshijima, K.; Inoue, K.; Higuchi, I.; Sakamoto, H.; Shimura, Y. Control of doublesex alternative splicing by transformer and transformer-2 in Drosophila. Science 1991, 252, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.G.; Kobayashi, S.; Aoki, F. Male-specific splicing of the silkworm Imp gene is maintained by an autoregulatory mechanism. Mech. Dev. 2014, 131, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Hedley, M.L.; Maniatis, T. Sex-specific splicing and polyadenylation of dsx pre-mRNA requires a sequence that binds specifically to tra-2 protein in vitro. Cell 1991, 65, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, V.; Ryner, L.C.; Baker, B.S. Regulation of sex-specific selection of fruitless 5′ splice sites by transformer and transformer-2. Mol. Cell. Biol. 1998, 18, 450–458. [Google Scholar] [CrossRef] [Green Version]
- Bertossa, R.C.; van de Zande, L.; Beukeboom, L.W. The Fruitless gene in Nasonia displays complex sex-specific splicing and contains new zinc finger domains. Mol. Biol. Evol. 2009, 26, 1557–1569. [Google Scholar] [CrossRef] [Green Version]
- Bopp, D.; Saccone, G.; Beye, M. Sex determination in insects: Variations on a common theme. Sex Dev. 2014, 8, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Manley, J.L.; Tacke, R. SR proteins and splicing control. Genes Dev. 1996, 10, 569–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graveley, B.R. Sorting out the complexity of SR protein functions. RNA 2000, 6, 1197–1211. [Google Scholar] [CrossRef] [Green Version]
- Kay, B.K.; Williamson, M.P.; Sudol, M. The importance of being proline: The interaction of proline-rich motifs in signaling proteins with their cognate domains. FASEB J. 2000, 14, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Hediger, M.; Henggeler, C.; Meier, N.; Perez, R.; Saccone, G.; Bopp, D. Molecular characterization of the key switch F provides a basis for understanding the rapid divergence of the sex-determining pathway in the housefly. Genetics 2010, 184, 155–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woyke, J. What happens to diploid drone larvae in a honey bee colony? J. Apic. Res. 1963, 2, 73–75. [Google Scholar] [CrossRef]
- Santomauro, G.; Oldham, N.; Boland, W.; Enbels, W. Cannibalism of diploid drone larvae in the honey bee (Apis mellifera) is released by odd pattern of cuticular substances. J. Apic. Res. 2004, 43, 69–74. [Google Scholar] [CrossRef]
- Wexler, J.; Delaney, E.K.; Belles, X.; Schal, C.; Wada-Katsumata, A.; Amicucci, M.J.; Kopp, A. Hemimetabolous insects elucidate the origin of sexual development via alternative splicing. Elife 2019, 8, e47490. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Sequence | Denaturation | Annealing | Elongation | N° Cycles |
|---|---|---|---|---|---|---|
| fem-F | femFF1 | ACATTTATATTATCTGAAAAATTAG | 98 °C | 57 °C | 68 °C | 35 |
| femFR1 | GCTTAGATCCTTCTCCCGTTC | 10 s | 30 s | 30 s | ||
| fem-M | femMF1 | ATTAGAATCTTCAGATGGTAC | 98 °C | 57 °C | 68 °C | 35 |
| femMR1 | TATGTAAAATTTAATATATTGCAC | 10 s | 30 s | 30 s | ||
| csd | csdF1 | ATGAAACGAAATATATCAAATTATTC | 98 °C | 55 °C | 68 °C | 30 |
| csdR1 | TCATTGATGCGTAGGTCCAAATC | 10 s | 30 s | 90 s | ||
| Amtra-2 | Amtra2F | CAATGAGTGACATTGAGCGAAG | 98 °C | 55 °C | 68 °C | 30 |
| Amtra2R | TTAATATCGACGTGGTGAATAAG | 10 s | 30 s | 60 s |
| Target | Primers | Sequence |
|---|---|---|
| e2 | FemRIPF0 | TTAGACAATCACGCAGTGAAGATAG |
| FemRIPR0 | CCCGTTCTTGTTGTATCATCCATTC | |
| e3a and e5 | FemRIPF1 | GGACCAGAAGATACTCAAGTTAGTGC |
| FemRIPR1 | CCCGTTCTTCTTTTGAGCATCACAT | |
| e3b | FemRIPF2 | TCAGCAGAACTCGTCAAAATGTATGT |
| FemRIPR2 | GAAACAGGCCCGGAATGCAAAAG | |
| i5 | FemRIPF3 | ACAGAACTCATCATTGAAGATTGAC |
| FemRIPR3 | TTAAATATAATTGGCGAGTTTTTGC | |
| EF-1α | ArEF-1qF | TCTGTCGTGGCATCTTGAGC |
| ArEF-1qF | TCTCCTGGGCTTCCTTCTCA | |
| fem-F | femFqF1 | AACATTTATATTATCTGAAAAATTAG |
| femFqR1 | GCTTAGATCCTTCTCCCGTTC | |
| csd | csdqF1 | ATGAAACGAAATATATCAAATTATTC |
| csdqR1 | TCATTGATGCGTAGGTCCAAATC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, Y.; Yamada, T.; Suzuki, M.G. In Vitro Comparison of Sex-Specific Splicing Efficiencies of fem Pre-mRNA under Monoallelic and Heteroallelic Conditions of csd, a Master Sex-Determining Gene in the Honeybee. J. Dev. Biol. 2023, 11, 10. https://doi.org/10.3390/jdb11010010
Suzuki Y, Yamada T, Suzuki MG. In Vitro Comparison of Sex-Specific Splicing Efficiencies of fem Pre-mRNA under Monoallelic and Heteroallelic Conditions of csd, a Master Sex-Determining Gene in the Honeybee. Journal of Developmental Biology. 2023; 11(1):10. https://doi.org/10.3390/jdb11010010
Chicago/Turabian StyleSuzuki, Yukihiro, Takafumi Yamada, and Masataka G. Suzuki. 2023. "In Vitro Comparison of Sex-Specific Splicing Efficiencies of fem Pre-mRNA under Monoallelic and Heteroallelic Conditions of csd, a Master Sex-Determining Gene in the Honeybee" Journal of Developmental Biology 11, no. 1: 10. https://doi.org/10.3390/jdb11010010