Investigating the Systems-Level Effect of Pueraria lobata for Menopause-Related Metabolic Diseases Using an Ovariectomized Rat Model and Network Pharmacological Analysis

 , , and
, , and
Abstract
:1. Introduction
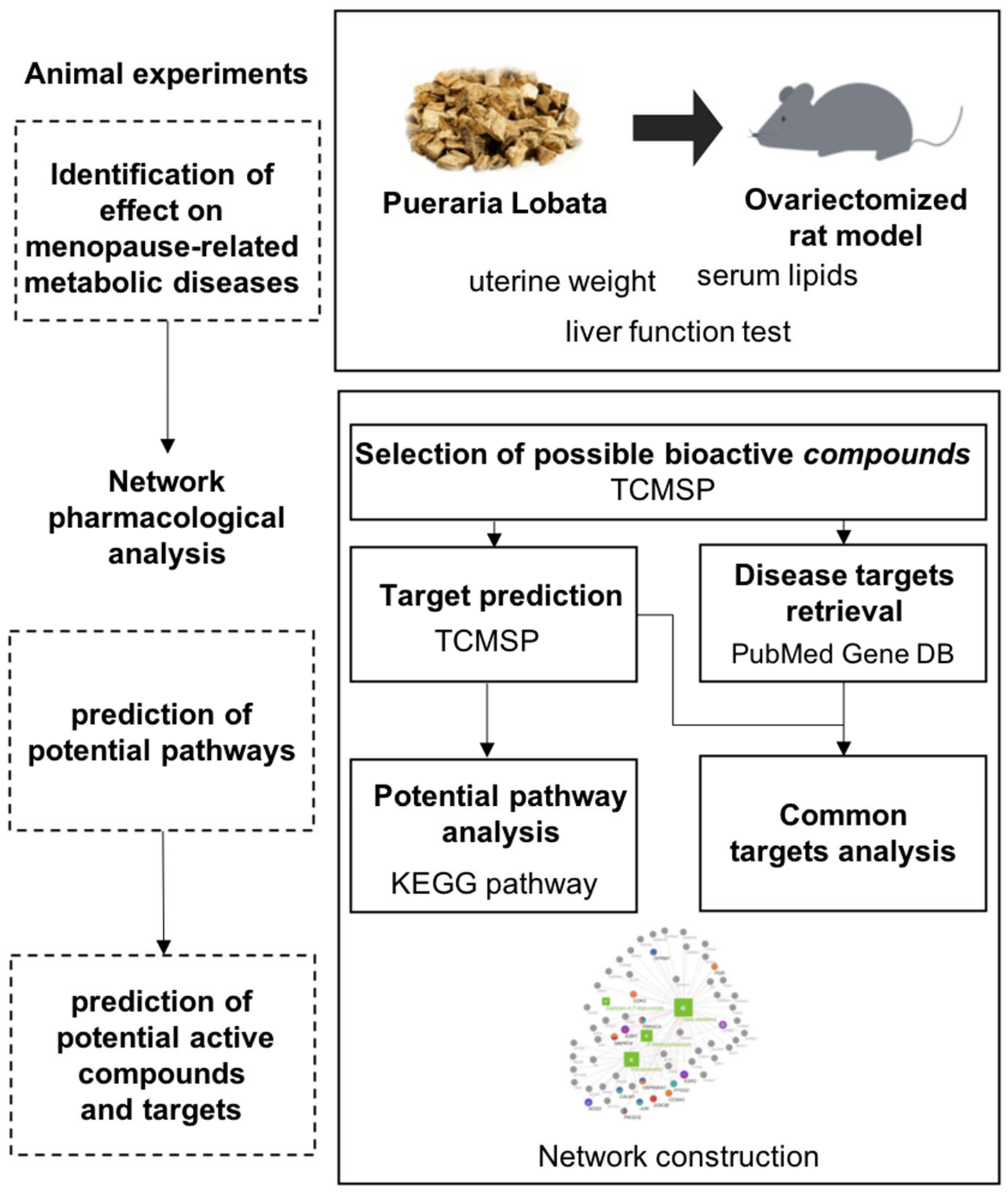
2. Materials and Methods
2.1. Preparation of the Herbal Formulation
2.2. Experimental Animals and Experimental Design
2.3. Blood and Organ Dissection
2.4. Serum Analyses
2.5. Measurement of PPAR-γ mRNA Expression
2.5.1. Tissues and Total RNA Extraction
2.5.2. cDNA Preparation
2.5.3. Real-Time Polymerase Chain Reaction
2.6. Network Pharmacological Analysis
2.7. Statistical Processing
3. Results
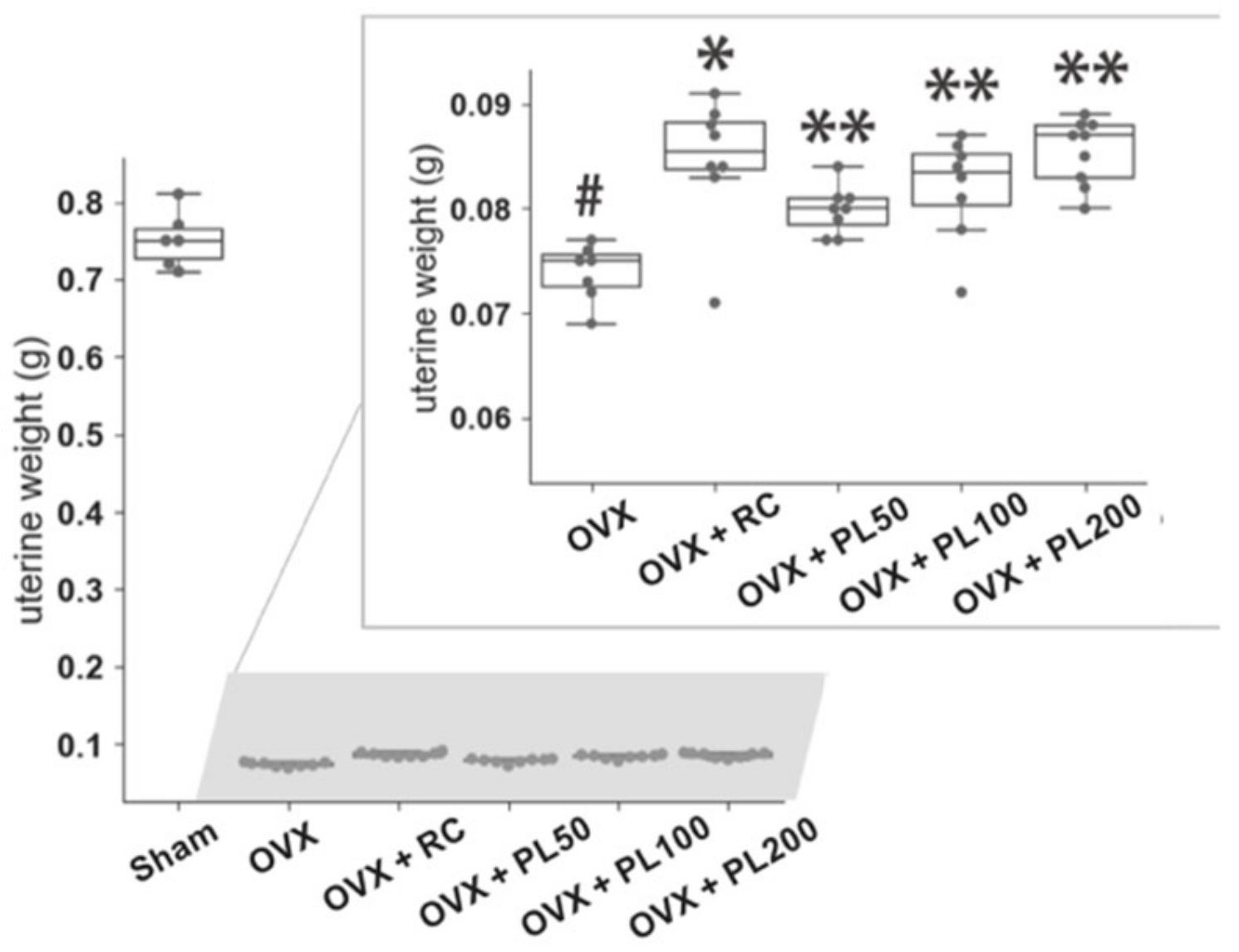
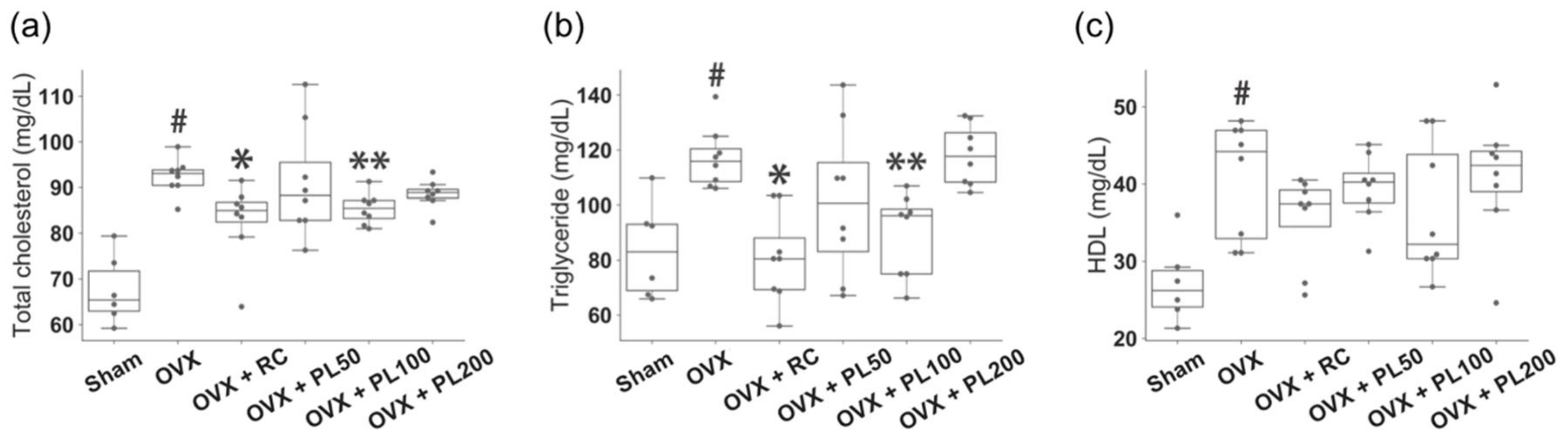
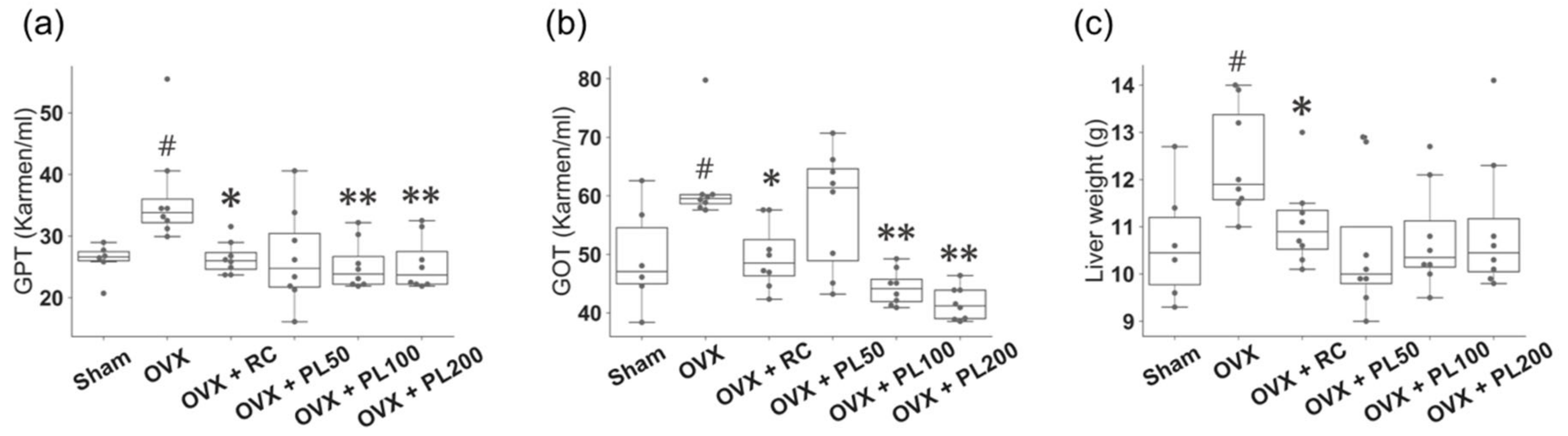
3.1. Animal Experiments
3.1.1. Uterine Weight
3.1.2. Serum Lipids
3.1.3. Glutamate Oxaloacetate Transaminase (GOT), Glutamate Pyruvate Transaminase (GPT), and Liver Weight
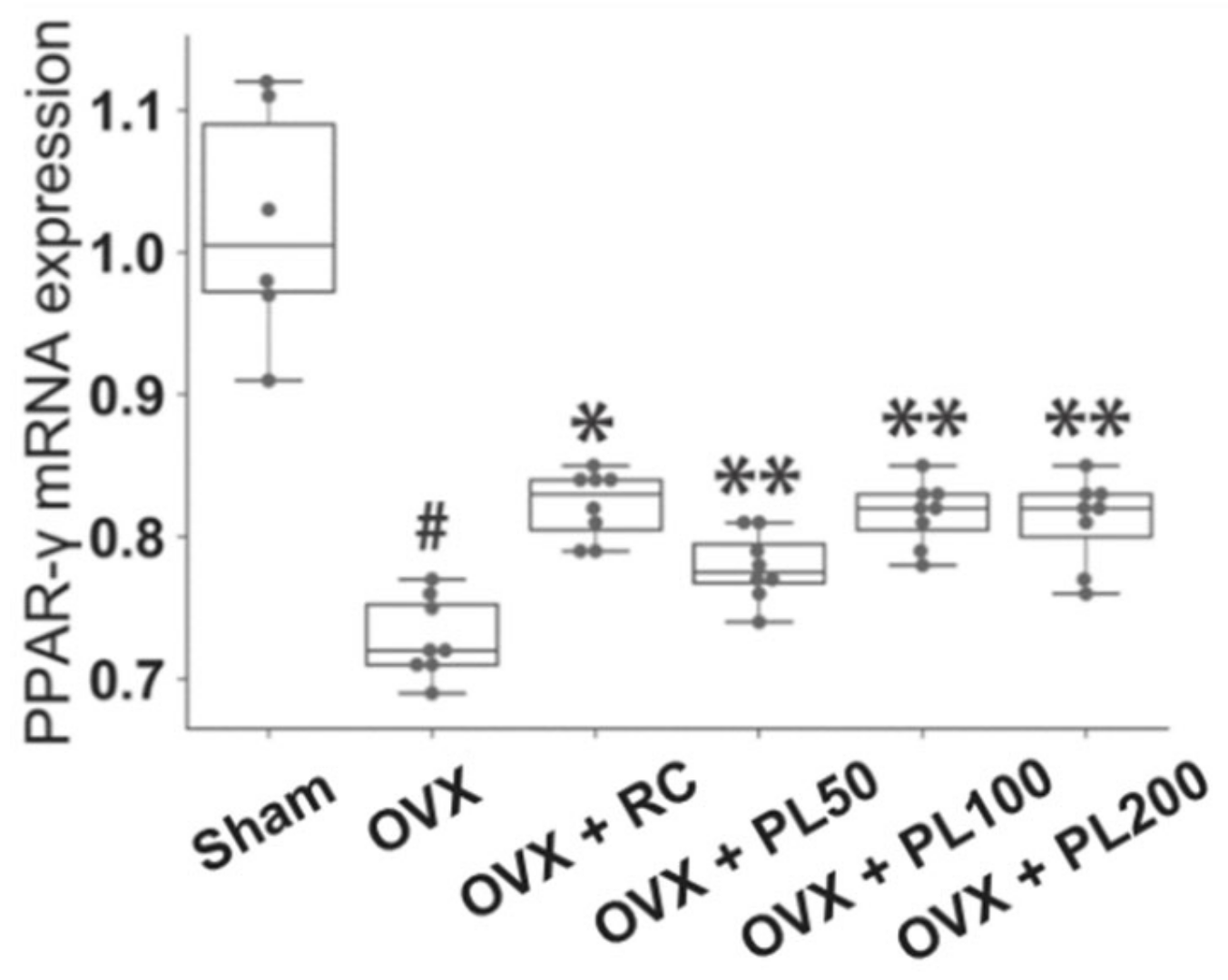
3.1.4. Peroxisome Proliferator-Activated Receptor Gamma Messenger RNA Expression
3.2. Network Pharmacological Analyses
3.2.1. Selection of Potential Bioactive Compounds of Pueraria lobata
3.2.2. Target Analysis
3.2.3. Identifying Potential Pathways of Pueraria lobata
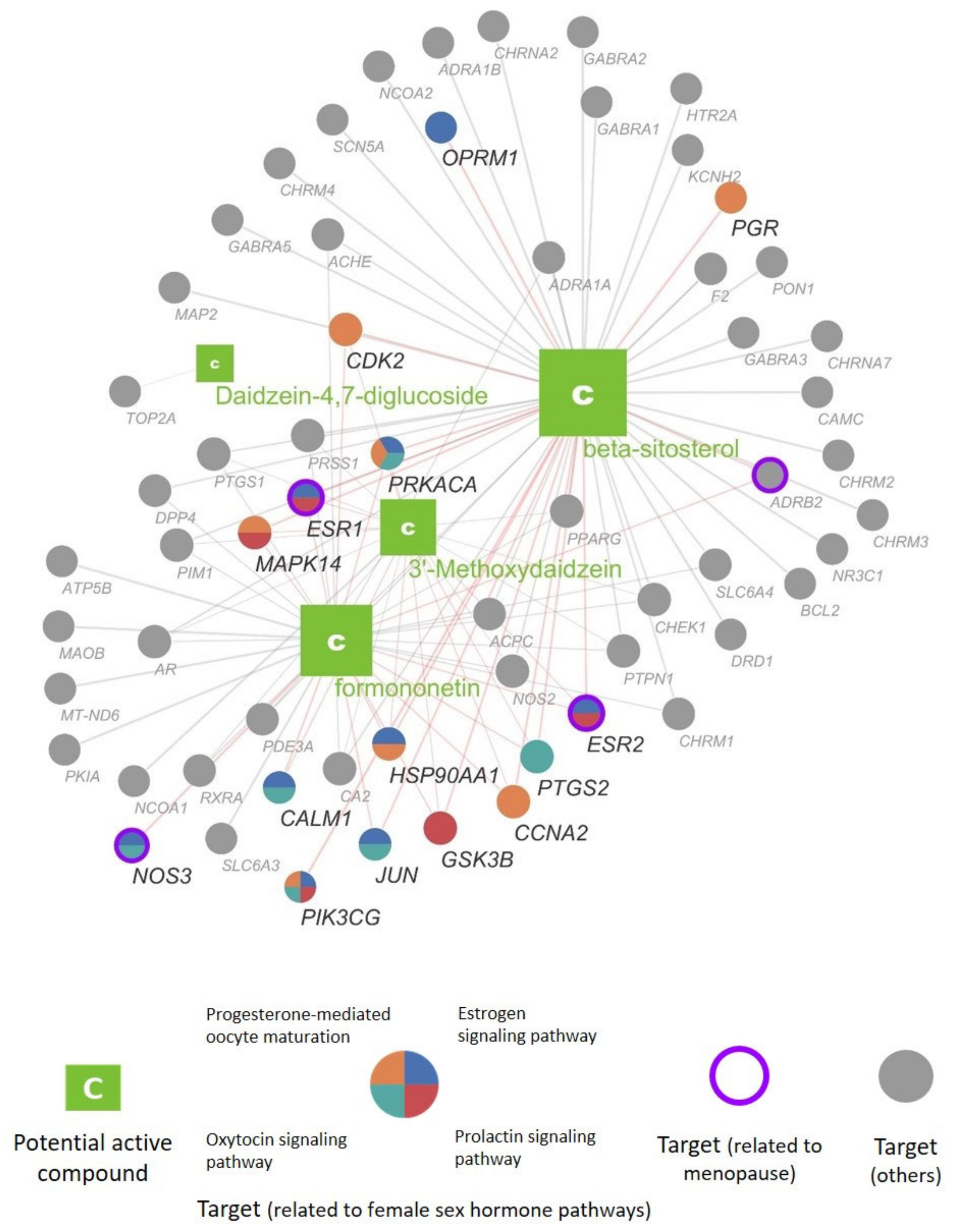
3.2.4. Construction of the Compound—Target Network of Pueraria lobata
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mauvais-Jarvis, F. Estrogen and androgen receptors: Regulators of fuel homeostasis and emerging targets for diabetes and obesity. Trends Endocrinol. Metab. 2011, 22, 24–33. [Google Scholar] [CrossRef]
- Zapantis, G.; Santoro, N. The menopausal transition: Characteristics and management. Best Pract. Res. Clin. Endocrinol. Metab. 2003, 17, 33–52. [Google Scholar] [CrossRef]
- Hallajzadeh, J.; Khoramdad, M.; Izadi, N.; Karamzad, N.; Almasi-Hashiani, A.; Ayubi, E.; Qorbani, M.; Pakzad, R.; Hasanzadeh, A.; Sullman, M.J.M.; et al. Metabolic syndrome and its components in premenopausal and postmenopausal women: A comprehensive systematic review and meta-analysis on observational studies. Menopause 2018, 25, 1155–1164. [Google Scholar] [CrossRef]
- Stefanska, A.; Bergmann, K.; Sypniewska, G. Metabolic Syndrome and Menopause: Pathophysiology, Clinical and Diagnostic Significance. In Advances in Clinical Chemistry; Academic Press: Cambridge, MA, USA, 2015; ISBN 9780128033142. [Google Scholar]
- Park, Y.W.; Zhu, S.; Palaniappan, L.; Heshka, S.; Carnethon, M.R.; Heymsfield, S.B. The metabolic syndrome: Prevalence and associated risk factor findings in the US population from the Third National Health and Nutrition Examination Survey, 1988–1994. Arch. Intern. Med. 2003, 163, 427–436. [Google Scholar] [CrossRef]
- Jouyandeh, Z.; Nayebzadeh, F.; Qorbani, M.; Asadi, M. Metabolic syndrome and menopause. J. Diabetes Metab. Disord. 2013, 12, 1. [Google Scholar] [CrossRef]
- Chang, W.-C.; Wang, J.-H.; Ding, D.-C. Hormone therapy in postmenopausal women associated with risk of stroke and venous thromboembolism: A population-based cohort study in Taiwan. Menopause 2019, 26, 197–202. [Google Scholar] [CrossRef]
- Trimarco, V.; Rozza, F.; Izzo, R.; de Leo, V.; Cappelli, V.; Riccardi, C.; di Carlo, C. Effects of a new combination of nutraceuticals on postmenopausal symptoms and metabolic profile: A crossover, randomized, double-blind trial. Int. J. Womens. Health 2016, 8, 581–587. [Google Scholar] [CrossRef]
- Jun, J.H.; Lee, H.W.; Zhang, J.; Yang, F.; Lee, M.S. Herbal medicine (Danggui Liuhuang decoction) for managing menopausal symptoms: A protocol of systematic review of randomized clinical trials. Medicine (Baltimore) 2018, 97, e9735. [Google Scholar] [CrossRef]
- Kargozar, R.; Azizi, H.; Salari, R. A review of effective herbal medicines in controlling menopausal symptoms. Electron. Physician 2017, 9, 5826–5833. [Google Scholar] [CrossRef]
- Franco, O.H.; Chowdhury, R.; Troup, J.; Voortman, T.; Kunutsor, S.; Kavousi, M.; Oliver-Williams, C.; Muka, T. Use of plant-based therapies and menopausal symptoms: A systematic review and meta-analysis. JAMA J. Am. Med. Assoc. 2016, 315, 2554–2563. [Google Scholar] [CrossRef]
- Liu, Z.M.; Ho, S.C.; Chen, Y.M.; Ho, S.; To, K.; Tomlinson, B.; Woo, J. Whole soy, but not purified daidzein, had a favorable effect on improvement of cardiovascular risks: A 6-month randomized, double-blind, and placebo-controlled trial in equol-producing postmenopausal women. Mol. Nutr. Food Res. 2014, 58, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Park, I.S.; Lee, H.W.; Ryuk, J.A.; Ko, B.S. Effects of an aqueous extract of dangguijagyagsan on serum lipid levels and blood flow improvement in ovariectomized rats. Evid. Based Complement. Altern. Med. 2014, 2014, 497836. [Google Scholar] [CrossRef] [PubMed]
- Go, H.; Ryuk, J.A.; Lee, H.W.; Ko, B.S. Palmiwon attenuates hepatic lipid accumulation and hyperlipidemia in a menopausal rat model. Menopause 2015, 4, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Li, G.Q.; Li, K.M.; Razmovski-Naumovski, V.; Chan, K. Kudzu root: Traditional uses and potential medicinal benefits in diabetes and cardiovascular diseases. J. Ethnopharmacol. 2011, 134, 584–607. [Google Scholar] [CrossRef]
- Lee, M.-K.; Cho, S.-Y.; Jang, J.-Y.; Choi, M.-S.; Jeon, S.-M.; Jang, M.K.; Kim, M.-J.; Park, Y.B. Effects of Puerariae Flos and Puerariae Radix Extracts on Antioxidant Enzymes in Ethanol-Treated Rats. Am. J. Chin. Med. 2003, 29, 343–354. [Google Scholar] [CrossRef]
- Zhang, R.; Hu, Y.; Yuan, J.; Wu, D. Effects of Puerariae radix extract on the increasing intestinal permeability in rat with alcohol-induced liver injury. J. Ethnopharmacol. 2009, 126, 207–214. [Google Scholar] [CrossRef]
- Luo, Y.; Zheng, S.; Ding, Y.; Dai, Y.; Zhou, Y.; Xiang, R.; Bay-Jensen, A.C.; Karsdal, M.A.; Qvist, P.; Zheng, Q. Preventive effects of kudzu root on bone loss and cartilage degradation in ovariectomized rat. Am. J. Transl. Res. 2017, 9, 5180. [Google Scholar]
- Jung, H.W.; Kang, A.N.; Kang, S.Y.; Park, Y.K.; Song, M.Y. The root extract of Pueraria lobata and its main compound, puerarin, prevent obesity by increasing the energy metabolism in skeletal muscle. Nutrients 2017, 9, 33. [Google Scholar] [CrossRef]
- Prasain, J.K.; Jones, K.; Kirk, M.; Wilson, L.; Smith-Johnson, M.; Weaver, C.; Barnes, S. Profiling and quantification of isoflavonoids in kudzu dietary supplements by high-performance liquid chromatography and electrospray ionization tandem mass spectrometry. J. Agric. Food Chem. 2003, 51, 4213–4218. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Jo, M.S.; Lee, D.; Baek, S.E.; Baek, J.; Yu, J.S.; Jo, J.; Yun, H.; Kang, K.S.; Yoo, J.E.; et al. Dual effects of isoflavonoids from Pueraria lobata roots on estrogenic activity and anti-proliferation of MCF-7 human breast carcinoma cells. Bioorg. Chem. 2019, 83, 135–144. [Google Scholar] [CrossRef]
- Liu, Z.H.; Sun, X.B. Network pharmacology: New opportunity for the modernization of traditional Chinese medicine. Yaoxue Xuebao 2012, 47, 696–703. [Google Scholar]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, W.; Huang, C.; Li, Y.; Yu, H.; Wang, Y.; Duan, J.; Ling, Y. A novel chemometric method for the prediction of human oral bioavailability. Int. J. Mol. Sci. 2012, 13, 6964–6982. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef]
- Maglott, D.; Ostell, J.; Pruitt, K.D.; Tatusova, T. Entrez gene: Gene-centered information at NCBI. Nucleic Acids Res. 2011, 39, D52–D57. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Yu, H.; Chen, J.; Xu, X.; Li, Y.; Zhao, H.; Fang, Y.; Li, X.; Zhou, W.; Wang, W.; Wang, Y. A systematic prediction of multiple drug-target interactions from chemical, genomic, and pharmacological data. PLoS ONE 2012, 7, e37608. [Google Scholar] [CrossRef]
- Barros, R.P.A.; Machado, U.F.; Warner, M.; Gustafsson, J.-A. Muscle GLUT4 regulation by estrogen receptors ERbeta and ER. Proc. Natl. Acad. Sci. USA 2006, 103, 1605–1608. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, M.; Fang, G.; Tang, Y.; Chen, Z.; Liu, X. Hypocholesterolemic effect of capsaicinoids by increased bile acids excretion in ovariectomized rats. Mol. Nutr. Food Res. 2013, 57, 1080–1088. [Google Scholar] [CrossRef]
- Katsuta, O.; Hiratsuka, H.; Matsumoto, J.; Tsuchitani, M.; Umemura, T.; Marumo, F. Ovariectomy enhances cadmium-induced nephrotoxicity and hepatotoxicity in rats. Toxicol. Appl. Pharmacol. 1993, 119, 267–274. [Google Scholar] [CrossRef]
- Gouda, H.N.; Sagoo, G.S.; Harding, A.H.; Yates, J.; Sandhu, M.S.; Higgins, J.P.T. The association between the peroxisome proliferator-activated receptor-γ2 (PPARG2) Pro12Ala gene variant and type 2 diabetes mellitus: A HuGE review and meta-analysis. Am. J. Epidemiol. 2010, 171, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yao, J.; Howe, A.A.; Menke, B.M.; Sivitz, W.I.; Spector, A.A.; Norris, A.W. Peroxisome Proliferator-Activated Receptor γ Decouples Fatty Acid Uptake from Lipid Inhibition of Insulin Signaling in Skeletal Muscle. Mol. Endocrinol. 2012, 26, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Norris, A.W.; Chen, L.; Fisher, S.J.; Szanto, I.; Ristow, M.; Jozsi, A.C.; Hirshman, M.F.; Rosen, E.D.; Goodyear, L.J.; Gonzalez, F.J.; et al. Muscle-specific PPARγ-deficient mice develop increased adiposity and insulin resistance but respond to thiazolidinediones. J. Clin. Investig. 2003, 112, 608–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hevener, A.L.; He, W.; Barak, Y.; Le, J.; Bandyopadhyay, G.; Olson, P.; Wilkes, J.; Evans, R.M.; Olefsky, J. Muscle-specific Pparg deletion causes insulin resistance. Nat. Med. 2003, 9, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Bai, Y.H.; Wang, S.T.; Zhu, Z.M.; Zhang, Y.W. Research on antioxidant effects and estrogenic effect of formononetin from Trifolium pratense (red clover). Phytomedicine 2009, 16, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Saeidnia, S. The Story of Beta-sitosterol—A Review. Eur. J. Med. Plants 2014, 4, 590. [Google Scholar] [CrossRef]
- Farquhar, J.W.; Smith, R.E.; Dempsey, M.E. The Effect of Beta Sitosterol on the Serum Lipids of Young Men with Arteriosclerotic Heart Disease. Circulation 1956, 14, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.H.; Helferich, W.G.; Clausen, L.M.; Allred, K.F.; Almada, A.L. β-Sitosterol, β-Sitosterol Glucoside, and a Mixture of β-Sitosterol and β-Sitosterol Glucoside Modulate the Growth of Estrogen-Responsive Breast Cancer Cells in Vitro and in Ovariectomized Athymic Mice. J. Nutr. 2004, 134, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Umehara, K.; Nemoto, K.; Matsushita, A.; Terada, E.; Monthakantirat, O.; De-Eknamkul, W.; Miyase, T.; Warashina, T.; Degawa, M.; Noguchi, H. Flavonoids from the heartwood of the Thai medicinal plant Dalbergia parviflora and their effects on estrogenic-responsive human breast cancer cells. J. Nat. Prod. 2009, 72, 2163–2168. [Google Scholar] [CrossRef]
- Casanova, M.; You, L.; Gaido, K.W.; Archibeque-Engle, S.; Janszen, D.B.; Heck, H.D.A. Developmental effects of dietary phytoestrogens in Sprague-Dawley rats and interactions of genistein and daidzein with rat estrogen receptors α and β in vitro. Toxicol. Sci. 1999, 51, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Kuźbicka, K.; Rachoń, D.; Woziwodzka, A.; Rybicka, M.; Bielawski, K.P. Associations of ESR1 and ESR2 gene polymorphisms with metabolic syndrome and its components in postmenopausal women. Maturitas 2018, 115, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Clapauch, R.; Mourão, A.F.; Mecenas, A.S.; Maranhão, P.A.; Rossini, A.; Bouskela, E. Endothelial function and insulin resistance in early postmenopausal women with cardiovascular risk factors: Importance of ESR1 and NOS3 polymorphisms. PLoS ONE 2014, 9, e103444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jocken, J.W.E.; Blaak, E.E.; Schiffelers, S.; Arner, P.; Van Baak, M.A.; Saris, W.H.M. Association of a beta-2 adrenoceptor (ADRB2) gene variant with a blunted in vivo lipolysis and fat oxidation. Int. J. Obes. 2007, 31, 813. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sense (5′-3′) | Antisense (5′-3′) |
|---|---|---|
| Peroxisome proliferator-activated receptor gamma (PPAR-γ) | TCPLAPLGCTCTGTCATC | CATCTGTACTPLTPLPLACA |
| Compound Name | MW | OB (%) | DL | AlogP | Hdon | Hacc | Caco-2 |
|---|---|---|---|---|---|---|---|
| Formononetin * | 268.28 | 69.67 | 0.21 | 2.58 | 1 | 4 | 0.78 |
| Sitogluside | 576.95 | 20.63 | 0.62 | 6.34 | 4 | 6 | −0.14 |
| Beta-sitosterol * | 414.79 | 36.91 | 0.75 | 8.08 | 1 | 1 | 1.32 |
| Daidzein | 254.25 | 19.44 | 0.19 | 2.33 | 2 | 4 | 0.59 |
| Ononin | 430.44 | 11.52 | 0.78 | 0.68 | 4 | 9 | −0.74 |
| Docosanoate | 340.66 | 15.69 | 0.26 | 9.11 | 1 | 2 | 1.21 |
| Lupenone | 424.78 | 11.66 | 0.78 | 7.36 | 0 | 1 | 1.48 |
| Genistein | 270.25 | 17.93 | 0.21 | 2.07 | 3 | 5 | 0.43 |
| Lignoceric acid | 368.72 | 14.9 | 0.33 | 10.02 | 1 | 2 | 1.24 |
| Scoparone | 206.21 | 74.75 | 0.09 | 1.87 | 0 | 4 | 0.85 |
| (R)-allantoin | 158.14 | 96.9 | 0.03 | −1.76 | 5 | 7 | −0.99 |
| 3′-Methoxydaidzein * | 284.28 | 48.57 | 0.24 | 2.32 | 2 | 5 | 0.56 |
| Daidzein-4,7-diglucoside * | 578.57 | 47.27 | 0.67 | −1.48 | 8 | 14 | −2.53 |
| Soyasapogenol b | 458.8 | 16.73 | 0.75 | 5.11 | 3 | 3 | 0.43 |
| Puerarin | 416.41 | 24.03 | 0.69 | −0.06 | 6 | 9 | −1.15 |
| 7,8,4′-Trihydroxyisoflavone | 270.25 | 20.67 | 0.22 | 2.07 | 3 | 5 | 0.45 |
| Daidzin | 416.41 | 14.32 | 0.73 | 0.43 | 5 | 9 | −1 |
| Sophoradiol | 442.8 | 17.42 | 0.76 | 6.2 | 2 | 2 | 0.95 |
| Predicted Targets of the Potential Bioactive Compounds of PL | ||||||||
|---|---|---|---|---|---|---|---|---|
| ESR1 | ESR2 | JUN | MT-ND6 | ATP5B | NOS2 | PTGS1 | F2 | CHRM1 |
| AR | PPARG | PTGS2 | NOS3 | CA2 | RXRA | ACHE | PDE3A | ADRA1A |
| PTPN1 | SLC6A3 | ADRB2 | SLC6A4 | DPP4 | MAPK14 | GSK3B | HSP90AA1 | CDK2 |
| ACPC | MAOB | CHEK1 | PRKACA | PRSS1 | PIM1 | CCNA2 | PKIA | CALM1 |
| BCL2 | PON1 | MAP2 | DRD1 | CHRM3 | KCNH2 | SCN5A | GABRA2 | CHRM4 |
| HTR2A | GABRA5 | GABRA3 | PGR | CHRM2 | ADRA1B | CHRNA2 | OPRM1 | NR3C1 |
| GABRA1 | PIK3CG | CHRNA7 | CAMC | NCOA2 | NCOA2 | TOP2A | ||
| Pathway | Combined Score | Adjusted p-Value | Genes (Targets) |
|---|---|---|---|
| Neuroactive ligand-receptor interaction † | 88.66 | 4.45 × 10−11 | GABRA2;CHRM2;GABRA1;CHRM3;PRSS1;CHRNA2;CHRM1;GABRA5;CHRM4;CHRNA7;GABRA3;ADRB2;OPRM1;HTR2A;F2;ADRA1B;NR3C1;ADRA1A;DRD1 |
| Calcium signaling pathway † | 62.88 | 1.39 × 10−7 | CHRM2;CHRM3;CHRM1;NOS2;NOS3;CHRNA7;ADRB2;HTR2A;ADRA1B;ADRA1A;DRD1;CALM1;PRKACA |
| Cholinergic synapse | 46.9 | 1.19 × 10−5 | CHRM2;CHRM3;ACHE;CHRM1;CHRM4;CHRNA7;BCL2;PRKACA;PIK3CG |
| Estrogen signaling pathway *† | 46.29 | 6.38 × 10−6 | HSP90AA1;JUN;NOS3;OPRM1;CALM1;PRKACA;ESR1;ESR2;PIK3CG |
| Pathways in cancer | 38.78 | 6.59 × 10−4 | AR;GSK3B;JUN;HSP90AA1;RXRA;NOS2;CDK2;BCL2;PPARG;PRKACA;PTGS2;PIK3CG |
| Morphine addiction | 38.13 | 2.63 × 10−5 | GABRA2;GABRA1;GABRA5;GABRA3;PDE3A;OPRM1;DRD1;PRKACA |
| Adrenergic signaling in cardiomyocytes † | 36.69 | 7.71 × 10−5 | BCL2;ADRB2;SCN5A;MAPK14;CALM1;PRKACA;ADRA1B;ADRA1A;PIK3CG |
| cAMP signaling pathway † | 32.7 | 4.14 × 10−4 | CHRM2;JUN;CHRM1;PDE3A;ADRB2;DRD1;CALM1;PRKACA;PIK3CG |
| Progesterone-mediated oocyte maturation * | 30.5 | 3.51 × 10−4 | CCNA2;HSP90AA1;CDK2;PGR;MAPK14;PRKACA;PIK3CG |
| Retrograde endocannabinoid signaling | 29.53 | 3.54 × 10−4 | GABRA2;GABRA1;GABRA5;GABRA3;MAPK14;PRKACA;PTGS2 |
| Amphetamine addiction | 27.59 | 3.54 × 10−4 | JUN;MAOB;DRD1;CALM1;PRKACA;SLC6A3 |
| Thyroid hormone signaling pathway † | 25.81 | 6.36 × 10−4 | NCOA1;NCOA2;GSK3B;RXRA;PRKACA;ESR1;PIK3CG |
| Dopaminergic synapse | 24.97 | 7.51 × 10−4 | GSK3B;MAOB;DRD1;MAPK14;CALM1;PRKACA;SLC6A3 |
| Epstein–Barr virus infection | 24.92 | 1.47 × 10−3 | CCNA2;GSK3B;JUN;CDK2;BCL2;MAPK14;PRKACA;PIK3CG |
| Taste transduction | 22.57 | 6.59 × 10−4 | GABRA2;GABRA1;CHRM3;GABRA5;GABRA3;PRKACA |
| Prostate cancer | 22.07 | 7.51 × 10−4 | GSK3B;AR;HSP90AA1;CDK2;BCL2;PIK3CG |
| AGE-RAGE signaling pathway in diabetic complications † | 21.27 | 1.32 × 10−3 | JUN;NOS3;PIM1;BCL2;MAPK14;PIK3CG |
| Small cell lung cancer | 21.1 | 7.45 × 10−4 | RXRA;NOS2;CDK2;BCL2;PTGS2;PIK3CG |
| PI3K-Akt signaling pathway † | 20.52 | 7.80 × 10−3 | CHRM2;GSK3B;HSP90AA1;CHRM1;RXRA;NOS3;CDK2;BCL2;PIK3CG |
| Salivary secretion † | 19.81 | 7.51 × 10−4 | CHRM3;ADRB2;CALM1;ADRA1B;PRKACA;ADRA1A |
| cGMP-PKG signaling pathway † | 19.71 | 2.42 × 10−3 | NOS3;PDE3A;ADRB2;CALM1;ADRA1B;ADRA1A;PIK3CG |
| Nicotine addiction | 18.71 | 3.80 × 10−4 | GABRA2;GABRA1;GABRA5;CHRNA7;GABRA3 |
| Serotonergic synapse | 16.7 | 2.05 × 10−3 | MAOB;HTR2A;PRKACA;PTGS2;SLC6A4;PTGS1 |
| Neurotrophin signaling pathway | 16.19 | 2.59 × 10−3 | GSK3B;JUN;BCL2;MAPK14;CALM1;PIK3CG |
| Regulation of lipolysis in adipocytes † | 15.31 | 8.42 × 10−4 | ADRB2;PTGS2;PRKACA;PIK3CG;PTGS1 |
| Cocaine addiction | 14.94 | 6.59 × 10−4 | JUN;MAOB;DRD1;PRKACA;SLC6A3 |
| Oxytocin signaling pathway *† | 14.46 | 8.60 × 10−3 | JUN;NOS3;CALM1;PRKACA;PTGS2;PIK3CG |
| Prolactin signaling pathway *† | 14.06 | 2.12 × 10−3 | GSK3B;MAPK14;ESR1;ESR2;PIK3CG |
| Inflammatory mediator regulation of TRP channels | 13.19 | 6.91 × 10−3 | HTR2A;MAPK14;CALM1;PRKACA;PIK3CG |
| Alcoholism | 12.61 | 1.34 × 10−2 | MAOB;PKIA;DRD1;CALM1;PRKACA;SLC6A3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.H.; Baek, S.-E.; Lee, W.-Y.; Baek, J.Y.; Trinh, T.A.; Park, D.H.; Lee, H.L.; Kang, K.S.; Kim, C.-E.; Yoo, J.-E. Investigating the Systems-Level Effect of Pueraria lobata for Menopause-Related Metabolic Diseases Using an Ovariectomized Rat Model and Network Pharmacological Analysis. Biomolecules 2019, 9, 747. https://doi.org/10.3390/biom9110747
Oh JH, Baek S-E, Lee W-Y, Baek JY, Trinh TA, Park DH, Lee HL, Kang KS, Kim C-E, Yoo J-E. Investigating the Systems-Level Effect of Pueraria lobata for Menopause-Related Metabolic Diseases Using an Ovariectomized Rat Model and Network Pharmacological Analysis. Biomolecules. 2019; 9(11):747. https://doi.org/10.3390/biom9110747
Chicago/Turabian StyleOh, Ji Hong, Seon-Eun Baek, Won-Yung Lee, Ji Yun Baek, Tuy An Trinh, Do Hwi Park, Hye Lim Lee, Ki Sung Kang, Chang-Eop Kim, and Jeong-Eun Yoo. 2019. "Investigating the Systems-Level Effect of Pueraria lobata for Menopause-Related Metabolic Diseases Using an Ovariectomized Rat Model and Network Pharmacological Analysis" Biomolecules 9, no. 11: 747. https://doi.org/10.3390/biom9110747