Stimulation of the Production of Prostaglandin E2 by Ethyl Gallate, a Natural Phenolic Compound Richly Contained in Longan


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. In Vitro Cell Culture Experiments
2.3. In Vivo Animal Experiments
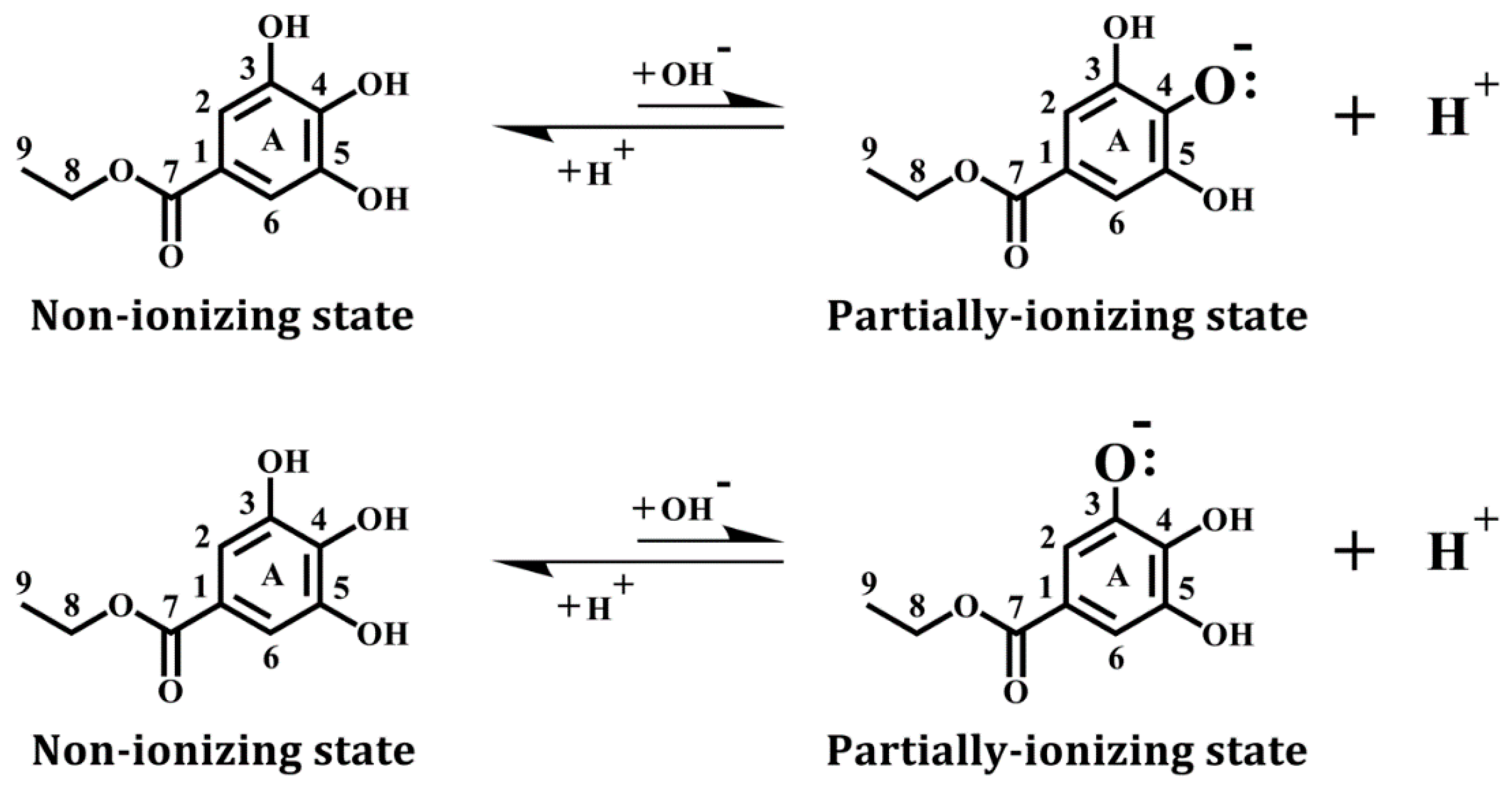
2.4. Molecular Docking Analysis of the Binding of Ethyl Gallate with COX Enzymes
2.4.1. Protein Processing
2.4.2. Ligand Processing
2.4.3. Flexible Docking
2.4.4. Calculation of Binding Energy
3. Results
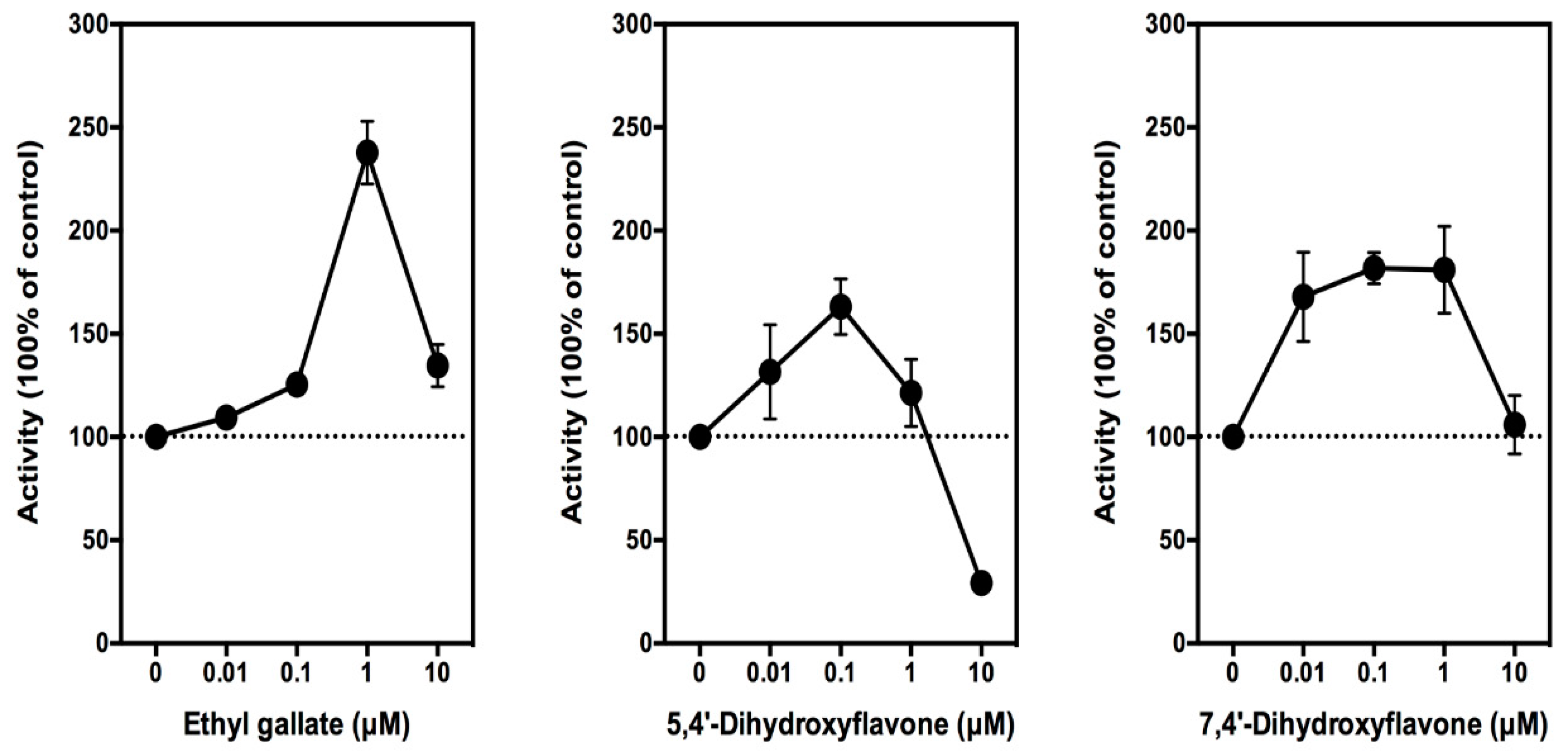
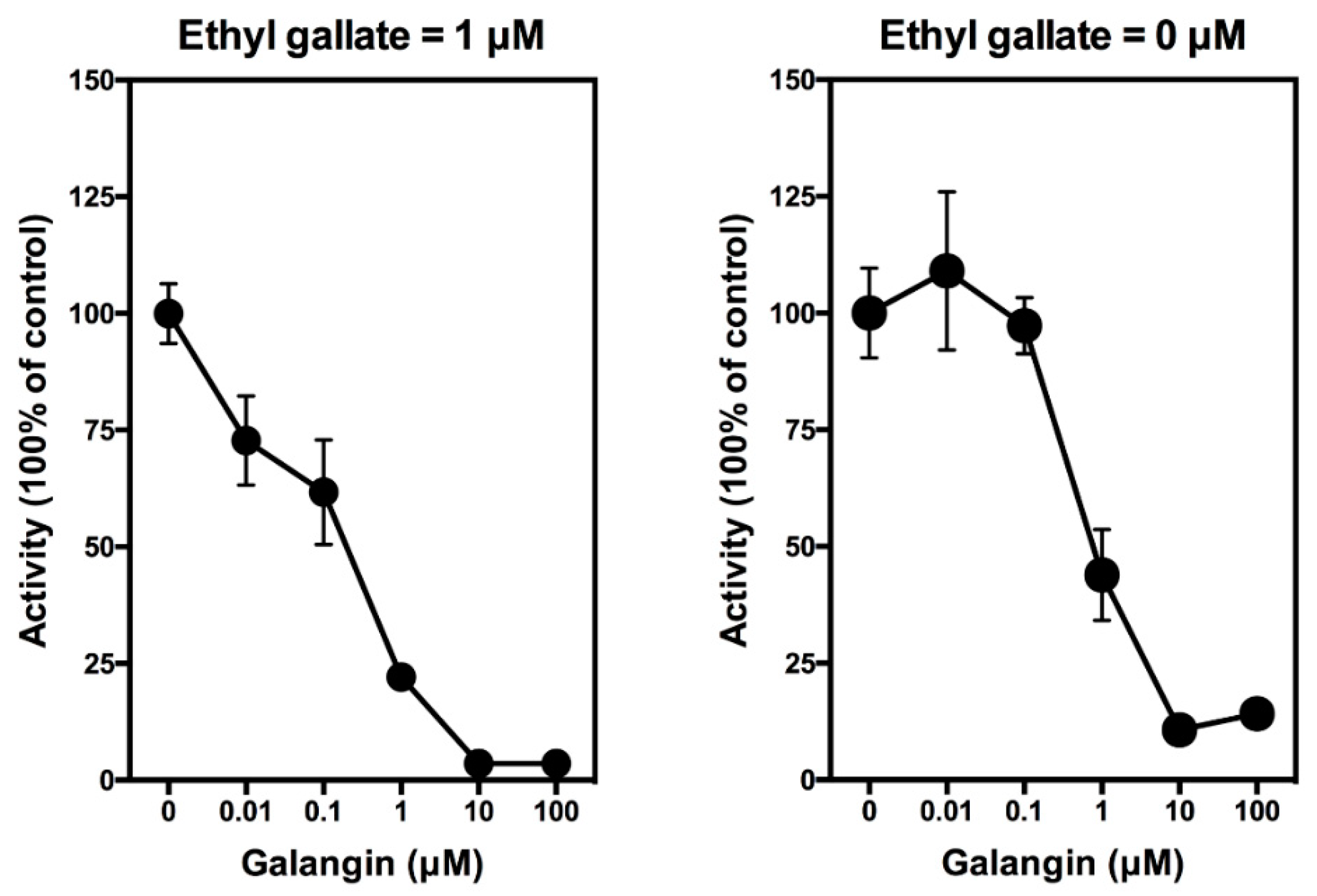
3.1. Effect of Ethyl Gallate on PGE2 Production In Vitro and In Vivo
3.1.1. In Vitro Studies
3.1.2. In Vivo Studies
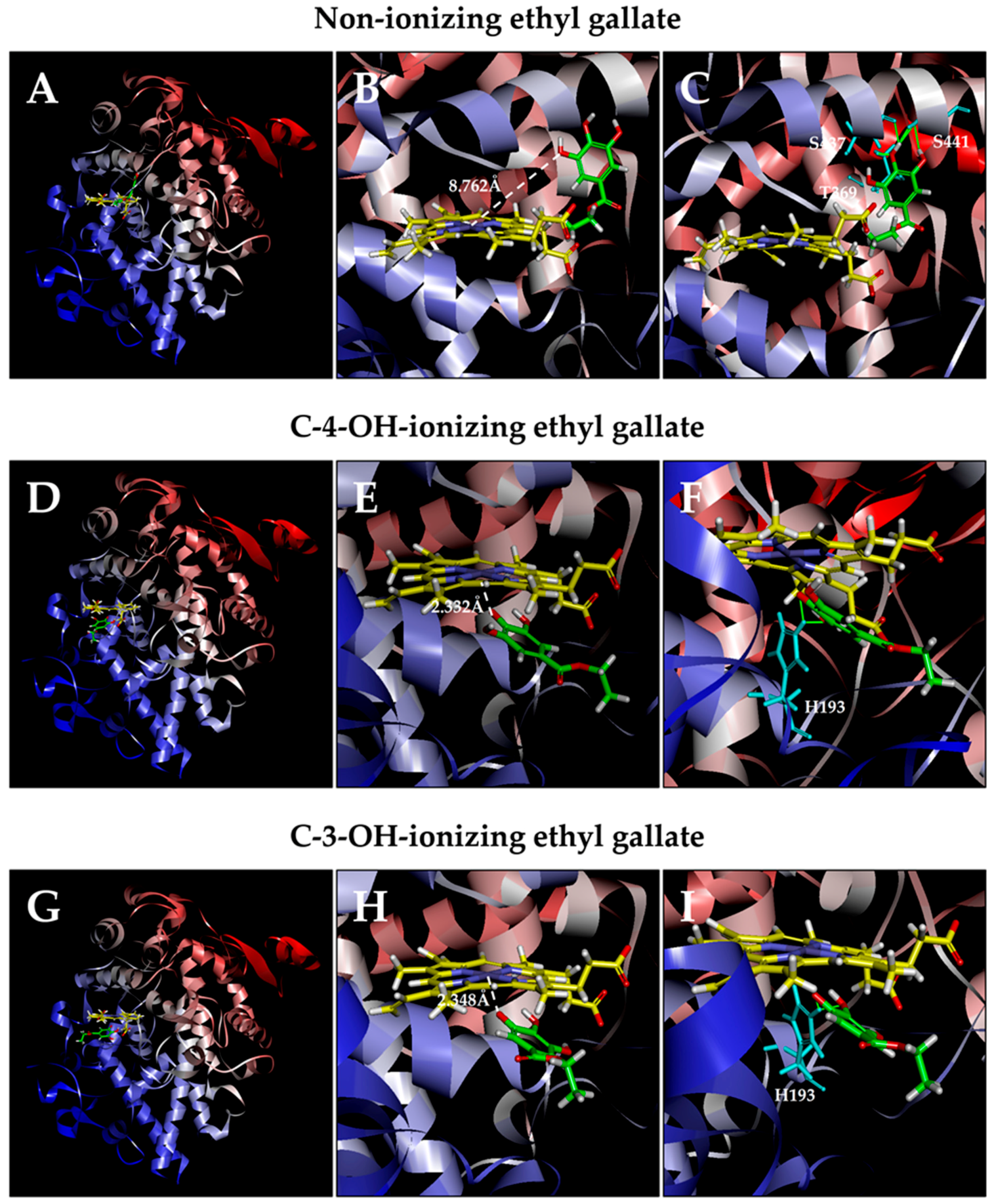
3.2. Computational Docking Analysis of Ethyl Gallate Binding Inside the Peroxidase Active Sites of COX-1 and COX-2
3.2.1. COX-1
3.2.2. COX-2
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Marnett, L.J. Cyclooxygenase mechanisms. Curr. Opin. Chem. Biol. 2000, 4, 545–552. [Google Scholar] [CrossRef]
- Fa, F. Cyclooxygenase enzymes: regulation and function. Curr. Pharmaceut. Des. 2004, 10, 577–588. [Google Scholar]
- Mitchell, J.A.; Kirkby, N.S. Eicosanoids, prostacyclin and cyclooxygenase in the cardiovascular system. Br. J. Pharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Duggan, K.C.; Walters, M.J.; Musee, J.; Harp, J.M.; Kiefer, J.R.; Oates, J.A.; Marnett, L.J. Molecular basis for cyclooxygenase inhibition by the non-steroidal anti-inflammatory drug naproxen. J. Biol. Chem. 2010, 285, 34950–34959. [Google Scholar] [CrossRef] [PubMed]
- Blobaum, A.L.; Marnett, L.J. Structural and functional basis of cyclooxygenase inhibition. J. Med. Chem. 2007, 50, 1425–1441. [Google Scholar] [CrossRef] [PubMed]
- Marnett, L.J.; Rowlinson, S.W.; Goodwin, D.C.; Kalgutkar, A.S.; Lanzo, C.A. Arachidonic acid oxygenation by COX-1 and COX-2. Mechanisms of catalysis and inhibition. J. Biol. Chem. 1999, 274, 22903–22906. [Google Scholar] [CrossRef] [PubMed]
- Kurumbail, R.G.; Kiefer, J.R.; Marnett, L.J. Cyclooxygenase enzymes: catalysis and inhibition. Curr. Opin. Struct. Biol. 2001, 11, 752–760. [Google Scholar] [CrossRef]
- Chen, H. The Production and Uses of litchis in China. South China Agricultural University. Available online: http://ir4.rutgers.edu/GMUS/presentation%20pdf/day1Chen.pdf (accessed on 4 July 2018).
- Menzel, C.M.; Waite, G.K. Litchi and Longan: Botany, Production and Uses; CABI: Wallingford, UK, 2005; ISBN 0-85199-696-5. [Google Scholar]
- Spencer, P.S.; Palmer, V.S. The enigma of litchi toxicity: an emerging health concern in southern Asia. Lancet Glob. Health 2017, 5, e383–e384. [Google Scholar] [CrossRef]
- Paireau, J.; Tuan, N.H.; Lefrançois, R.; Buckwalter, M.R.; Nghia, N.D.; Hien, N.T.; Lortholary, O.; Poirée, S.; Manuguerra, J.C.; Gessain, A.; et al. Litchi-associated acute encephalitis in children, Northern Vietnam, 2004–2009. Emerg. Infect. Dis. 2012, 18, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.; Kumar, A.; Thomas, J.D.; Laserson, K.F.; Bhushan, G.; Carter, M.D.; Chhabra, M.; Mittal, V.; Khare, S.; Sejvar, J.J.; et al. Association of acute toxic encephalopathy with litchi consumption in an outbreak in Muzaffarpur, India, 2014: a case-control study. Lancet Glob. Health 2017, 5, e458–e466. [Google Scholar] [CrossRef]
- Islam, M.S. Outbreak of sudden death with acute encephalitis syndrome among children associated with exposure to lychee orchards in Northern Bangladesh, 2012. Am. J. Trop. Med. Hyg. 2017, 97, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Xu, L.; Wu, P.; Xie, H.; Jiang, Y.; Chen, F.; Wei, X. Polyphenols from longan seeds and their radical-scavenging activity. Food Chem. 2009, 116, 433–436. [Google Scholar] [CrossRef]
- Tseng, H.C.; Wu, W.T.; Huang, H.S.; Wu, M.C. Antimicrobial activities of various fractions of longan (Dimocarpus longan Lour. Fen Ke) seed extract. Int. J. Food Sci. Nutr. 2014, 65, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Soong, Y.Y.; Barlow, P.J. Isolation and structure elucidation of phenolic compounds from longan (Dimocarpus longan Lour.) seed by high-performance liquid chromatography-electrospray ionization mass spectrometry. J. Chromat. A 2005, 1085, 270–277. [Google Scholar] [CrossRef]
- Chen, J.; Ge, Z.Z.; Zhu, W.; Xu, Z.; Li, C.M. Screening of key antioxidant compounds of longan (Dimocarpus longan Lour.) seed extract by combining online fishing/knockout, activity evaluation, Fourier transform ion cyclotron resonance mass spectrometry, and high-performance liquid chromatography electrospray ionization mass spectrometry methods. J. Agric. Food Chem. 2014, 62, 9744–9750. [Google Scholar] [PubMed]
- Zhang, Z.; Liao, L.; Moored, J.; Wu, T.; Wang, Z. Antioxidant phenolic compounds from walnut kernels (Juglans regia L.). Food Chem. 2009, 113, 160–165. [Google Scholar] [CrossRef]
- Marzouk, M.S.; El-Toumy, S.A.; Moharram, F.A.; Shalaby, N.M.; Ahmed, A.A. Pharmacologically active ellagitannins from Terminalia myriocarpa. Planta Medica 2002, 68, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Suárez, R.; Gómez-Cordovés, C.; Bartolomé, B. Simultaneous determination of nonanthocyanin phenolic compounds in red wines by HPLC-DAD/ESI-MS. Am. J. Enol. Vitic. 2005, 56, 139–147. [Google Scholar]
- Bai, H.W.; Zhu, B.T. Strong activation of cyclooxygenase I and II catalytic activity by dietary bioflavonoids. J. Lipid Res. 2008, 49, 2557–2570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Bai, H.W.; Zhu, B.T. Structural basis for certain naturally occurring bioflavonoids to function as reducing cosubstrates of cyclooxygenase I and II. PLoS ONE 2010, 5, e12316. [Google Scholar]
- Bai, H.W.; Zhu, B.T. Myricetin and quercetin are naturally occurring cosubstrates of cyclooxygenases in vivo. Prostaglandins Leukot. Essent. Fatty Acids 2010, 82, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.R.; Sui, H.C.; Zhu, B.T. Ellagic acid, a plant phenolic compound, activates cyclooxygenase-mediated prostaglandin production. Exp. Ther. Med. accepted.
- Gupta, K.; Selinsky, B.S.; Kaub, C.J.; Katz, A.K.; Loll, P.J. The 2.0 Å resolution crystal structure of prostaglandin H2 synthase-1: Structural insights into an unusual peroxidase. J. Mol. Biol. 2004, 335, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Feig, M.; Onufriev, A.; Lee, M.S.; Im, W.; Case, D.A.; Brooks, C.L. Performance comparison of generalized Born and Poisson methods in the calculation of electrostatic solvation energies for protein structures. J. Comp. Chem. 2004, 25, 265–284. [Google Scholar] [CrossRef] [PubMed]
- Uciechowska, U.; Schemies, J.; Scharfe, M.; Lawson, M.; Wichapong, K.; Jung, M.; Sippl, W. Binding free energy calculations and biological testing of novel thiobarbiturates as inhibitors of the human NAD+ dependent histone deacetylase Sirt2. MedChemComm 2012, 3, 167–173. [Google Scholar] [CrossRef]
- Pouplana, R.; Lozano, J.J.; Ruiz, J. Molecular modelling of the differential interaction between several non-steroidal anti-inflammatory drugs and human prostaglandin endoperoxide H synthase-2 (h-PGHS-2). J. Mol. Graph. Model. 2002, 20, 329–343. [Google Scholar] [CrossRef]
- Zhu, B.T.; Bai, H.W.; Rao, S.; Sui, H.C. Galangin inhibits cyclooxygenase by blocking the function of the reducing cosubstrate at the peroxidase site. FASEB J. under review.
- Prasad, K.N.; Yang, B.; Yang, S.; Chen, Y.; Zhao, M.; Ashraf, M.; Jiang, Y. Identification of phenolic compounds and appraisal of antioxidant and antityrosinase activities from litchi (Litchi sinensis Sonn.) seeds. Food Chem. 2009, 116, 1–7. [Google Scholar] [CrossRef]
- Queiroz, E.R.; Abreu, C.M.P.; Oliveira, K.S.; Ramos, V.O.; Fráguas, R.M. Bioactive phytochemicals and antioxidant activity in fresh and dried lychee fractions. Rev. Ciênc. Agron. 2015, 46, 163–169. [Google Scholar] [CrossRef]
- Rangkadilok, N.; Sitthimonchai, S.; Worasuttayangkurn, L.; Mahidol, C.; Ruchirawat, M.; Satayavivad, J. Evaluation of free radical scavenging and antityrosinase activities of standardized longan fruit extract. Food Chem. Toxicol. 2007, 45, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Vattem, D.A.; Shetty, K. Biological function of ellagic acid: A review. J. Food Biochem. 2005, 29, 234–266. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Protein | Binding Energy Value ΔGbinding (kcal/mol) | ||
|---|---|---|---|
| No ionization | C-4-OH ionization | C-3-OH ionization | |
| COX-1 protein | −3.652 | −94.444 | −108.779 |
| COX-2 protein | −6.398 | −100.929 | −132.742 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.R.; Sui, H.C.; Ding, Y.Y.; Zhu, B.T. Stimulation of the Production of Prostaglandin E2 by Ethyl Gallate, a Natural Phenolic Compound Richly Contained in Longan. Biomolecules 2018, 8, 91. https://doi.org/10.3390/biom8030091
Wang HR, Sui HC, Ding YY, Zhu BT. Stimulation of the Production of Prostaglandin E2 by Ethyl Gallate, a Natural Phenolic Compound Richly Contained in Longan. Biomolecules. 2018; 8(3):91. https://doi.org/10.3390/biom8030091
Chicago/Turabian StyleWang, Hui Rong, Hao Chen Sui, Yan Yan Ding, and Bao Ting Zhu. 2018. "Stimulation of the Production of Prostaglandin E2 by Ethyl Gallate, a Natural Phenolic Compound Richly Contained in Longan" Biomolecules 8, no. 3: 91. https://doi.org/10.3390/biom8030091