Recombinant Fusion Protein Joining E Protein Domain III of Tick-Borne Encephalitis Virus and HSP70 of Yersinia pseudotuberculosis as an Antigen for the TI-Complexes

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Chimeric Plasmids
2.2. Expression of the Recombinant Plasmids
2.3. Recombinant Protein Purification
2.4. Fluorescence Spectroscopy
2.5. Calorimetry
2.6. Preparation of TI-Complexes
2.7. Animals and Immunization
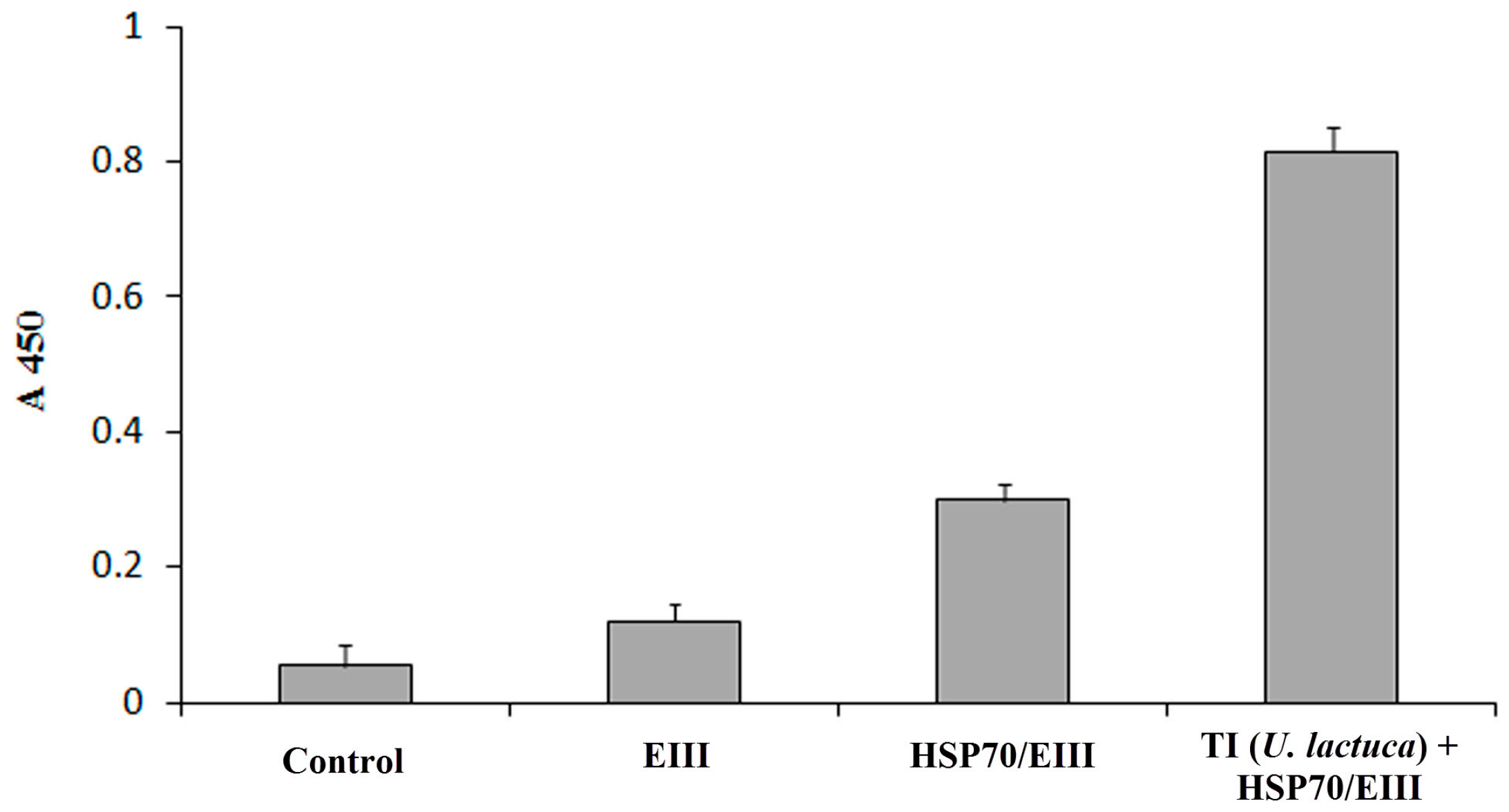
2.8. ELISA and Statistical Analyses
2.9. Computational Analysis of the Intrinsic Disorder Predisposition of the Chimeric HSP70/EIII Protein
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haditsch, M.; Kunze, U. Tick-borne encephalitis: A disease neglected by travel medicine. Travel Med. Infect. Dis. 2013, 11, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Kollaritsch, H.; Paulke-Korinek, M.; Holzmann, H.; Hombach, J.; Bjorvatn, B.; Barrett, A. Vaccines and vaccination against tick-borne encephalitis. Expert Rev. Vaccines 2012, 11, 1103–1119. [Google Scholar] [CrossRef] [PubMed]
- Ershova, A.S.; Gra, O.A.; Lyaschuk, A.M.; Grunina, T.M.; Tkachuk, A.P.; Bartov, M.S.; Savina, D.M.; Sergienko, O.V.; Galushkina, Z.M.; Gudov, V.P.; et al. Recombinant domains III of tick-borne encephalitis virus envelope protein in combination with dextran and CpGs induce immune response and partial protectiveness against TBE virus infection in mice. BMC Infect. Dis. 2016, 16, 544. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.S. Application of radiation technology in vaccines development. Clin. Exp. Vaccine Res. 2015, 4, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A structural perspective of the flavivirus life cycle. Nat. Rev. Microbiol. 2005, 3, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Füzik, T.; Formanová, P.; Růžek, D.; Yoshii, K.; Niedrig, M.; Plevka, P. Structure of tick-borne encephalitis virus and its neutralization by a monoclonal antibody. Nat. Commun. 2018, 9, 436. [Google Scholar] [CrossRef] [PubMed]
- Bressanelli, S.; Stiasny, K.; Allison, S.L.; Stura, E.A.; Duquerroy, S.; Lescar, J.; Heinz, F.X.; Rey, F.A. Structure of a flavivirus envelope glycoprotein in its low-pH-induced membrane fusion conformation. EMBO J. 2004, 23, 728–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenkova, A.M.; Chopenko, N.S.; Davydova, L.A.; Mazeika, A.N.; Bystritskaya, E.P.; Portnyagina, O.Y.; Anastyuk, S.D.; Kulbatskii, D.S.; Lyukmanova, E.N.; Dolgikh, D.A.; et al. Engineering of chimeric protein based on E protein domain III of tick- borne encephalitis virus and OmpF porin of Yersinia pseudotuberculosis. Protein Pept. Lett. 2017, 24, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Ciocca, D.R.; Cayado-Gutierrez, N.; Maccioni, M.; Cuello-Carrion, F.D. Heat shock proteins (HSPs) based anti-cancer vaccines. Curr. Mol. Med. 2012, 12, 1183–1197. [Google Scholar] [CrossRef] [PubMed]
- Sanina, N.M.; Kostetsky, E.Y.; Shnyrov, V.L.; Tsybulsky, A.V.; Novikova, O.D.; Portniagina, O.Y.; Vorobieva, N.S.; Mazeika, A.N.; Bogdanov, M.V. The influence of monogalactosyldiacylglycerols from different marine macrophytes on immunogenicity and conformation of protein antigen of tubular immunostimulating complex. Biochimie 2012, 94, 1048–1056. [Google Scholar] [CrossRef] [PubMed]
- Sanina, N.; Davydova, L.; Chopenko, N.; Kostetsky, E.; Shnyrov, V. Modulation of immunogenicity and conformation of HA1 subunit of influenza A virus H1/N1 hemagglutinin in tubular immunostimulating complexes. Int. J. Mol. Sci. 2017, 18, 1895. [Google Scholar] [CrossRef] [PubMed]
- Vorobieva, N.; Sanina, N.; Vorontsov, V.; Kostetsky, E.; Mazeika, A.; Tsybulsky, A.; Kim, N.; Shnyrov, V. On the possibility of lipid-induced regulation of conformation and immunogenicity of influenza a virus H1/N1 hemagglutinin as antigen of TI-complexes. J. Mol. Microbiol. Biotechnol. 2014, 24, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Belikov, S.I.; Kondratov, I.G.; Potapova, U.V.; Leonova, G.N. The relationship between the structure of the tick-borne encephalitis virus strains and their pathogenic properties. PLoS ONE 2014, 9, e94946. [Google Scholar] [CrossRef] [PubMed]
- Golotin, V.A.; Balabanova, L.A.; Noskova, Y.A.; Slepchenko, L.V.; Bakunina, I.Y.; Vorobieva, N.S.; Terenteva, N.A.; Rasskazov, V.A. Optimization of cold-adapted alpha-galactosidase expression in Escherichia coli. Protein Expr. Purif. 2016, 123, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kostetsky, E.Y.; Sanina, N.M.; Mazeika, A.N.; Tsybulsky, A.V.; Vorobyeva, N.S.; Shnyrov, V.L. Tubular immunostimulating complex based on cucumarioside A2-2 and monogalactosyldiacylglycerol from marine macrophytes. J. Nanobiotechnol. 2011, 9, 9–35. [Google Scholar] [CrossRef] [PubMed]
- Sanina, N.M.; Goncharova, S.N.; Kostetsky, E.Y. Seasonal change of fatty acid composition and thermotropic behavior of polar lipids from marine macrophytes. Phytochemistry 2008, 69, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Portnyagina, O.Y.; Novikova, O.D.; Vostrikova, O.P.; Solov’eva, T.F. Dynamics of immune response to porine from Yersinia pseudotuberculosis outer membrane. Bull. Exp. Biol. Med. 1999, 128, 437–440. [Google Scholar] [CrossRef]
- Xue, B.; Dunbrack, R.L.; Williams, R.W.; Dunker, A.K.; Uversky, V.N. PONDR-FIT: A meta-predictor of intrinsically disordered amino acids. Biochim. Biophys. Acta 2010, 1804, 996–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, P.; Obradovic, Z.; Li, X.; Garner, E.C.; Brown, C.J.; Dunker, A.K. Sequence complexity of disordered protein. Proteins 2001, 42, 38–48. [Google Scholar] [CrossRef]
- Obradovic, Z.; Peng, K.; Vucetic, S.; Radivojac, P.; Dunker, A.K. Exploiting heterogeneous sequence properties improves prediction of protein disorder. Proteins 2005, 61, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, Z.; Peng, K.; Vucetic, S.; Radivojac, P.; Brown, C.J.; Dunker, A.K. Predicting intrinsic disorder from amino acid sequence. Proteins 2003, 53, 566–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosztanyi, Z.; Csizmok, V.; Tompa, P.; Simon, I. IUPred: Web server for the prediction of intrinsically unstructured regions of proteins based on estimated energy content. Bioinformatics 2005, 21, 3433–3434. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Vucetic, S.; Radivojac, P.; Brown, C.J.; Dunker, A.K.; Obradovic, Z. Optimizing long intrinsic disorder predictors with protein evolutionary information. J. Bioinform. Comput. Biol. 2005, 3, 35–60. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.L.; Kurgan, L. Comprehensive comparative assessment of in-silico predictors of disordered regions. Curr. Protein Pept. Sci. 2012, 13, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Kurgan, L. Accurate prediction of disorder in protein chains with a comprehensive and empirically designed consensus. J. Biomol. Struct. Dyn. 2014, 32, 448–464. [Google Scholar] [CrossRef] [PubMed]
- Dunker, A.K.; Lawson, J.D.; Brown, C.J.; Williams, R.M.; Romero, P.; Oh, J.S.; Oldfield, C.J.; Campen, A.M.; Ratliff, C.M.; Hipps, K.W.; et al. Intrinsically disordered protein. J. Mol. Graph. Model. 2001, 19, 26–59. [Google Scholar] [CrossRef] [Green Version]
- Prilusky, J.; Felder, C.E.; Zeev-Ben-Mordehai, T.; Rydberg, E.H.; Man, O.; Beckmann, J.S.; Silman, I.; Sussman, J.L. FoldIndex: A simple tool to predict whether a given protein sequence is intrinsically unfolded. Bioinformatics 2005, 21, 3435–3438. [Google Scholar] [CrossRef] [PubMed]
- Campen, A.; Williams, R.M.; Brown, C.J.; Meng, J.; Uversky, V.N.; Dunker, A.K. TOP-IDP-scale: A new amino acid scale measuring propensity for intrinsic disorder. Protein Pept. Lett. 2008, 15, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Dosztanyi, Z.; Csizmok, V.; Tompa, P.; Simon, I. The pairwise energy content estimated from amino acid composition discriminates between folded and intrinsically unstructured proteins. J. Mol. Biol. 2005, 347, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Walsh, I.; Giollo, M.; Di Domenico, T.; Ferrari, C.; Zimmermann, O.; Tosatto, S.C. Comprehensive large-scale assessment of intrinsic protein disorder. Bioinformatics 2015, 31, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Kurgan, L. On the complementarity of the consensus-based disorder prediction. In Proceedings of the Pacific Symposium on Biocomputing, Fairmont Orchid, HI, USA, 3–7 January 2012; pp. 176–187. [Google Scholar]
- Golotin, V.; Balabanova, L.; Likhatskaya, G.; Rasskazov, V. Recombinant production and characterization of a highly active alkaline phosphatase from marine bacterium Cobetia marina. Mar. Biotechnol. 2015, 17, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.; Takemoto, S.; Takakura, Y. Heat shock protein derivatives for delivery of antigens to antigen presenting cells. Int. J. Pharm. 2008, 354, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Suzue, K.; Young, R.A. Heat shock proteins as immunological carriers and vaccines. EXS 1996, 77, 451–465. [Google Scholar] [PubMed]
- Jiang, J.; Xie, D.; Zhang, W.; Xiao, G.; Wen, J. Fusion of Hsp70 to Mage-a1 enhances the potency of vaccine-specific immune responses. J. Transl. Med. 2013, 111, 300. [Google Scholar] [CrossRef] [PubMed]
- Mahalka, A.K.; Kirkegaard, T.; Jukola, L.T.; Jaatela, M.; Kinnunen, P.K.J. Human heat shock protein 70 (Hsp70) as a peripheral membrane protein. Biochim. Biophys. Acta 2014, 1838, 1344–1361. [Google Scholar] [CrossRef] [PubMed]
- Armijo, G.; Okerblom, J.; Cauvi, D.M.; Lopez, V.; Schlamadinger, D.E.; Kim, J.; Arispe, N.; De Maio, A. Interaction of heat shock protein 70 with membranes depends on the lipid environment. Cell Stress Chaperones 2014, 19, 877–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.Q.; Sturtevant, J.M. Thermodynamic study of yeast phosphoglycerate kinase. Biochemistry 1987, 26, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Arribas, A.; Santamaría, R.I.; Zhadan, G.G.; Villar, E.; Shnyrov, V.L. Differential scanning calorimetric study of the thermal stability of xylanase from Streptomyces halstedii JM8. Biochemistry 1994, 33, 13787–13791. [Google Scholar] [CrossRef] [PubMed]
- Garnier, C.; Protasevich, I.; Gilli, R.; Tsvetkov, P.; Lobachov, V.; Peyrot, V.; Briand, C.; Makarov, A. The two-stage process of the heat shock protein 90 thermal denaturation: Effect of calcium and magnesium. Biochem. Biophys. Res. Commun. 1998, 249, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Sanina, N.; Chopenko, N.; Mazeika, A.; Kostetsky, E. Nanoparticulate tubular immunostimulating complexes: Novel formulation of effective adjuvants and antigen delivery systems. Biomed. Res. Int. 2017. [Google Scholar] [CrossRef] [PubMed]
- Permyakov, E.A. The Method of Intrinsic Protein Luminescence; Nauka: Moscow, Russia, 2003; pp. 73–88. ISBN 5-02-006453-X. [Google Scholar]
- Reshetnyak, Y.; Burstein, E. Decomposition of protein tryptophan fluorescence spectra into log-normal components. II. The statistical proof of discreteness of tryptophan classes in proteins. Biophys. J. 2001, 81, 1710–1734. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| dnaK-X-Nco-dir | 5′-ATATCCATGGCGATGGGTAAAATTATTGGTATCGAC-3′ |
| dnaK-X-Sac-rev | 5′-TATAGAGCTCGCTTTTTTGTCTTTTACTTCTTCGAATTC-3′ |
| E3-X-Sac-L-dir | 5′-TATAGAGCTCGGGTGGTGGTGGTTCTGGTGGTGGTGGTTCTGGTGGTGGTGGTTCTGGTCTTACATACACAATGTGCGACAAGACGAAATTCAC-3′ |
| E3-X-Sal-stop-rev | 5′-TATA GTCGAC TTA GTTAAAGGCACCGCCAAGAACTGT-3′ |
| Hsp70 | HSP70/EIII | HSP70/EIII + MGDG U. lactuca | |
|---|---|---|---|
| Tm1, °C | 54.6 ± 0.1 | 54.2 ± 0.1 | 55.4 ± 0.1 |
| ΔH1, kcal/mol | 92.1 ± 0.7 | 99.5 ± 1.7 | 96.9 ± 1.4 |
| Tm2, °C | - | 58.4 ± 0.3 | 63.2 ± 0.1 |
| ΔH2, kcal/mol mol−1 | - | 79.9 ± 1.5 | 103.0 ± 1.9 |
| Tm3, °C | 66.4 ± 0.2 | 67.1 ± 0.1 | 67.7 ± 0.1 |
| ΔH3, kcal/mol | 81.5 ± 1.5 | 116.0 ± 1.5 | 170.0 ± 1.9 |
| Tm4, °C | 68.5 ± 0.4 | 70.1 ± 0.6 | 71.3 ± 0.5 |
| ΔH4, kcal/mol | 129.0 ± 1.3 | 124.0 ± 1.9 | 126.0 ± 2.6 |
| Protein | Spectral Forms (%) | |||
|---|---|---|---|---|
| S | I | II | III | |
| HSP70 | 13.6 ± 0.3 | 58.5 ± 0.5 | 12.2 ± 0.3 | 15.7 ± 0.2 |
| HSP70/EIII | 19.5 ± 0.5 | 45.7 ± 0.4 | 34.8 ± 0.5 | 0 |
| HSP70/EIII + MGDG U. lactuca | 19.3 ± 0.5 | 42.8 ± 0.5 | 0 | 37.9 ± 0.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golotin, V.; Sanina, N.; Davydova, L.; Chopenko, N.; Mazeika, A.; Roig, M.; Shnyrov, V.; Uversky, V.N.; Kostetsky, E. Recombinant Fusion Protein Joining E Protein Domain III of Tick-Borne Encephalitis Virus and HSP70 of Yersinia pseudotuberculosis as an Antigen for the TI-Complexes. Biomolecules 2018, 8, 82. https://doi.org/10.3390/biom8030082
Golotin V, Sanina N, Davydova L, Chopenko N, Mazeika A, Roig M, Shnyrov V, Uversky VN, Kostetsky E. Recombinant Fusion Protein Joining E Protein Domain III of Tick-Borne Encephalitis Virus and HSP70 of Yersinia pseudotuberculosis as an Antigen for the TI-Complexes. Biomolecules. 2018; 8(3):82. https://doi.org/10.3390/biom8030082
Chicago/Turabian StyleGolotin, Vasily, Nina Sanina, Ludmila Davydova, Natalia Chopenko, Andrey Mazeika, Manuel Roig, Valery Shnyrov, Vladimir N. Uversky, and Eduard Kostetsky. 2018. "Recombinant Fusion Protein Joining E Protein Domain III of Tick-Borne Encephalitis Virus and HSP70 of Yersinia pseudotuberculosis as an Antigen for the TI-Complexes" Biomolecules 8, no. 3: 82. https://doi.org/10.3390/biom8030082