
Transcriptional Regulation of Chemokine Expression in Ovarian Cancer

Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Systematic Name | Alternate Human Names | Tissue/Cells | Reference |
|---|---|---|---|
| CCL2 | Monocyte chemotactic protein 1 (MCP-1) | Tumor biopsies, serum and ascites | Negus et al., 1995 [18]
Milliken et al., 2002 [19] |
| CCL5 | RANTES | Tumor ascites, plasma and peritoneal fluid | Milliken et al., 2002 [19]
Negus et al., 1997 [20] |
| CCL11 | Eotaxin | Primary ovarian cancer cells obtained from ascites | Levina et al., 2009 [21]
Nolen et al., 2010 [22] |
| CCL25 | Thymus expressed chemokine (TECK) | Tumor tissue | Singh et al., 2011 [23] |
| CCL28 | Mucosae-associated epithelial chemokine (MEC) | Tumor tissue | Facciabene et al., 2011 [24] |
| CXCL1 | Growth-regulated protein α (GRO-α) | Plasma and tumor ascites | Lee et al., 2006 [25]
Yang et al., 2006 [26] |
| CXCL2 | Growth-regulated protein β (GRO-β) | Ovarian cancer cell lines | Son et al., 2007 [27]
Kavandi et al., 2012 [28] |
| CXCL8 | Interleukin 8 (IL-8) | Tumor tissue, ascites, serum and cyst fluid | Lee et al., 1996 [29]
Xu et al., 1999 [30] |
| CXCL12 | Stromal cell-derived factor (SDF-1) | Tumor biopsies, tissues and ascites | Zou et al., 2001 [31]
Scotton et al., 2002 [32] |
| CXCL16 | Transmembrane chemokine CXCL16 | Epithelial ovarian carcinoma tissue | Guo et al., 2011 [33]
Gooden et al., 2014 [34] |
| CX3CL1 | Fractalkine | Epithelial ovarian carcinoma tissue | Gaudin et al., 2011 [35] |
| XCL1/2 | Lymphotactin | Tumor ascites and ovarian cancer cell lines | Kim et al., 2012 [36] |
2. Mechanisms Regulating Chemokine Transcription in Ovarian Cancer Cells
2.1. Chemokine Regulation by NFκB and Epigenetic Acetylation
2.2. Chemokine Modulation by Hypoxia and Metabolism
2.3. Chemokine Modulation by Chemotherapeutic Interventions
3. Chemokine Transcriptional Regulation in Ovarian Cancer Cells
| Factor | Site | Sequence | Factor | Site | Sequence |
|---|---|---|---|---|---|
| SP-1 | -54/-44 | ACTCCGCCCT | c-Fos | -1465/-1457 | CTGACTCC |
| Nkx-1 | -65/-58 | CCTCCTG | p53 | -1541/-1534 | GGGCAGG |
| Elk-1 | -76/-71 | GGAAG | HOX-11 | -1571/-1564 | CCTAACG |
| GATA | -88/-82 | CTTATC | PEA3 | -1644/-1636 | AAACATCC |
| C/EBP | -112/-106 | TTGCTC | GR | -1790/-1782 | TTGTTCTC |
| ELF | -143/-130 | CTACTTCCTGGAA | AR | -1789/-1781 | TGTTCTCT |
| Hif-1 * | -127/-122 | CACAG | FOXP3 | -1959/-1950 | AAACATTTT |
| AP-1 * | -139/-131 | TTCCTGGAA | C/EBP | -1980/-1973 | TTGCACA |
| STAT1-3 * | -139/-131 | TTCCTGGAA | Pbx-1 | -2132/-2120 | AGCATGACTGGA |
| C-Ets1 | -140/-133 | CTTCCTG | FOXO-3 | -2184/-2176 | CTTATTTA |
| NF-AT | -181/-172 | GGAAAAAGT | CUTL-1 | -2309/-2303 | ATTGGT |
| E47 | -239/-232 | GTCTGGG | PR | -2358/-2351 | GAACACT |
| RP58 | -256/-245 | GTTCACATCTG | Smad3 | -2521/-2511 | GAGGCAGACA |
| HNF-1 | -654/-646 | TAATATTT | ERα | -2570/-2562 | CTGACCTC |
| TMF | -708/-701 | TATAACA | c-Jun | -2580/-2574 | CATGGG |
| HNF-3 | -742/-735 | CTATTTA | NFκB * | -2600/-2591 | GGAATTTCC |
| AP-2 | -747/-741 | GCAGGC | ZDX/BCL6 | -2632/-2621 | GGGAACTTCC |
| c-Jun | -942/-935 | TGACTTA | E47 | -2678/-2671 | ATCTGGA |
| HMG1 | -1042/-1035 | GGAAATT | ETF | -2717/-2708 | CACAGCCCC |
| IRF-3 | -1089/-1082 | GCTTTCC | GATA | -2902/-2893 | CTTTATCT |
| BTEB3 | -1287/-1278 | AGGAGGAGG | PU-1 | -3041/-3031 | TTACTTCCTC |
| NF-Y | -1315/-1307 | ATTGGGCA | YY1 | -3264/-3257 | AAAATGG |
| USF-2b | -1447/-1439 | GTCATTTG | RAR | -3429/-3421 | ATCTCACC |
| Factor | Site | Sequence | Factor | Site | Sequence |
|---|---|---|---|---|---|
| IRF-3 | -50/-43 | GCTTTCC | Elk-1 | -771/-766 | GGAAG |
| HMG I | -75/-68 | AATTTCC | FOXP3 | -791/-782 | CAACATTTT |
| MBP-1 | -78/-68 | GGGAATTTCC | MZF-1 | -810/-803 | CAGGGGA |
| NFκB * | -79/-68 | CGGGAATTTCC | TGIF | -870/-862 | TGACAACC |
| CDP * | -97/-87 | GGGATCGATC | C/EBP | -980/-974 | TTGCAC |
| E47 | -90/-83 | ATCTGGA | YY-1 | -1061/-1054 | TAAATGG |
| E2F-1 | -126/-119 | GGCGGGG | c-Ets | -1076/-1069 | CAGGAAG |
| SP3 | -128/-119 | GGGGCGGGG | AR | -1394/-1386 | TGTTCTCT |
| SP-1 * | -130/-121 | GGGGGCGGG | c-Jun | -1491/-1483 | TGACTCAT |
| R2 | -137/-131 | TCCACC | Pax | -1909/-1902 | CCTTGAC |
| LF-A1 | -247/-240 | TGGGGCA | ERα | -2057/-2050 | TGGGTCAA |
| AP-2 * | -279/-273 | GCAGGC | NF-Y | -2060/-2052 | ATTGGGTC |
| AREB6 | -296/-288 | CAGGTGGT | LEF-1 | -2807/-2799 | CTTTGTTG |
| Smad3 | -563/-553 | TTCACAGACA | HNF-1 | -2966/-2958 | TAATATTT |
| PR | -602/-595 | GAACATT | RAR | -3102/-3094 | ATGCCTTAG |
| GR | -605/-596 | GCAGAACAT | NHP-1 | -3103/-3096 | TGACCTT |
| TMF | -739/-732 | TGTTATA | PEA3 | -3110/-3102 | GGATGTAT |
| GATA | -767/-761 | GATAAG | ATF | -3452/-3443 | TGACGTAAA |
| Factor | Site | Sequence | Factor | Site | Sequence |
|---|---|---|---|---|---|
| NFκB * | -76/-67 | GGGAATTTCC | BTEB3 | -862/-853 | AAGCGGAGT |
| CREB | -83/-74 | CGGACGTCA | NF-Y | -970/-962 | GAACCAAT |
| ATF-2 | -83/-74 | CGGACGTCA | HMG I | -999/-992 | AATTTCC |
| HLF | -104/-95 | GTTACGCAA | IRF | -999/-992 | AATTTCC |
| E2F-1 | -111/-104 | GGCGGGA | NF-AT | -1001/-992 | AAAATTTCC |
| NF-1 | -113/-108 | TTGGC | CUTL1 | -1085/-1079 | ATTGAT |
| LF-A1 | -139/-132 | CGGGGCA | FOXP3 | -1115/-1106 | CTTAATTTT |
| GATA | -192/-184 | GGTTATCT | PR A | -1257/-1250 | GAACACT |
| AP2α | -198/-192 | GCAGGC | C/EBP | -1367/-1360 | TGAGCAA |
| STAT3 * | -218/-210 | TTGGGGAA | MZF1 | -1380/-1373 | CAGGGGA |
| ERα | -241/-233 | CTGACCCA | HNF-1 | -1440/-1432 | ATATTAAC |
| PEA3 | -276/-268 | GGATGTAG | TMF | -1880/-1873 | TATAACA |
| Elk-1 | -296/-292 | GAAG | E47 | -1830/-1823 | TTCTGGA |
| STAT3 * | -318/-310 | GGGATCGATC | Nkx2 | -1827/-1820 | CTGGAGG |
| p53 | -339/-332 | CTTGCCC | HNF | -2153/-2146 | TAAATGG |
| AhR | -418/-410 | GCGTGCGT | YY1 | -2153/-2146 | TAAATGG |
| c-Jun * | -437/-430 | TGACACA | HSF1 | -2409/-2401 | ATTCTAGG |
| c-Fos | -451/-443 | TGCGTCAT | ETF | -2505/-2496 | GGGGCTGTC |
| c-Ets | -473/-467 | CAGGAAG | AP3 | -2636/-2629 | GAGTTAG |
| USF-1 | -508/-499 | ACACGTGAT | Smad3 | -3112/-3102 | CAGTCAGACA |
| AREB6 | -574/-566 | AACACCTG | LEF-1 | -3101/-3093 | CAACAAAG |
| FOXJ2 | -621/-611 | AAAATAAACA | TCF-1 | -3102/-3093 | ACAACAAAG |
| AR | -673/-665 | TGTTCCAA | GR | -3256/-3247 | ACAGAACAT |
| Factor | Site | Sequence | Factor | Site | Sequence |
|---|---|---|---|---|---|
| NFκB * | -80/-70 | GGAATTTCC | E47 | -859/-852 | ATCTGGA |
| PU-1 | -83/-73 | GGAATTTCCTC | PR | -868/-861 | ACTCTTC |
| NRF * | -88/-77 | ATTCCTCTGA | HSF1 | -867/868 | CCTTGAAT |
| C/EBP * | -94/-87 | TTGCAAA | IRF | -973/-964 | TTTCCATTA |
| MZF-1 | -112/-105 | GAGGGA | RAR | -1068/-1061 | AGAGGTC |
| EBF | -118/-107 | TGCCCTGAGGG | ERα | -1067/-1060 | GAGGTCA |
| C/EBP * | -119/-112 | TTGCACA | p53 | -1258/-1251 | CTTGCCC |
| AP-1 * | -129/-121 | TGACTCAG | FOXP3 | -1304/-1295 | AAAATGAAG |
| c-Ets | -141/-132 | TAGGAAGTC | RelA | -1367/-1357 | GGCATTCCCC |
| Elk-1 | -139/-134 | GGAAG | YY1 | -1372/-1365 | AAAATGG |
| LEF-1 | -187/-179 | GATCAAAG | Smad3 | -1403/-1393 | GAAACAGACA |
| Hif-1 * | -234/-229 | GTGCG | Nkx1 | -1457/-1450 | CCTCAAG |
| GRα | -335/-327 | TTGTTCTA | AP2α | -1473/-1467 | CCAGGC |
| AREB6 | -328/-320 | AACACCTG | TCF1 | -1663/-1654 | ACAACAAAG |
| AR | -334/-326 | TGTTCTAA | NF-AT | -1687/-1677 | CTAATTTTCC |
| NF | -424/-416 | ATTGGCTC | HMGI | -1685/-1677 | AATTTTCC |
| AP3 | -535/-528 | TAAATC | HLF | -1695/-1686 | TTGTGTAAC |
| HNF-3 | -606/-599 | TAAATGT | CUTL1 | -1858/1852 | TTGGT |
| FOXO3 | -651/-641 | CTTATCTA | PEA3 | -2174/-2166 | GCACATCC |
| GATA | -651/-644 | CTTTATCT | HOX11 | -2200/-2193 | CGTTAGG |
| c-Myb | -792/-784 | CAACTGCC | RARγ | -2225/-2217 | GGCTCACC |
| C/EBP | -798/-792 | TTGCTC | AIRE | -2555/-2545 | ATGGTTATCT |
| GR | -847/-838 | CTGTTCTCT | Oct1 | -2744/-2733 | TCACTTTGCAT |
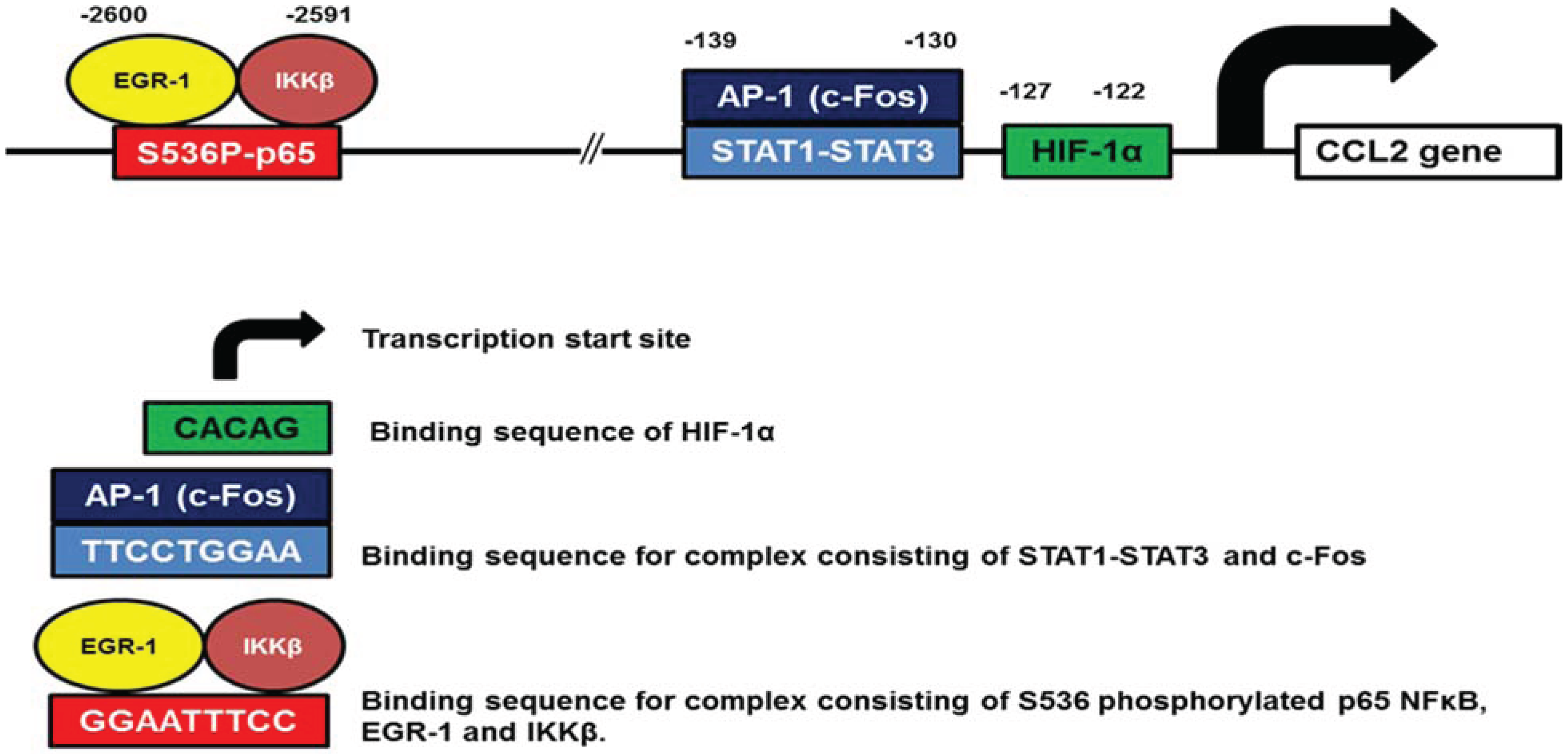
3.1. CCL2

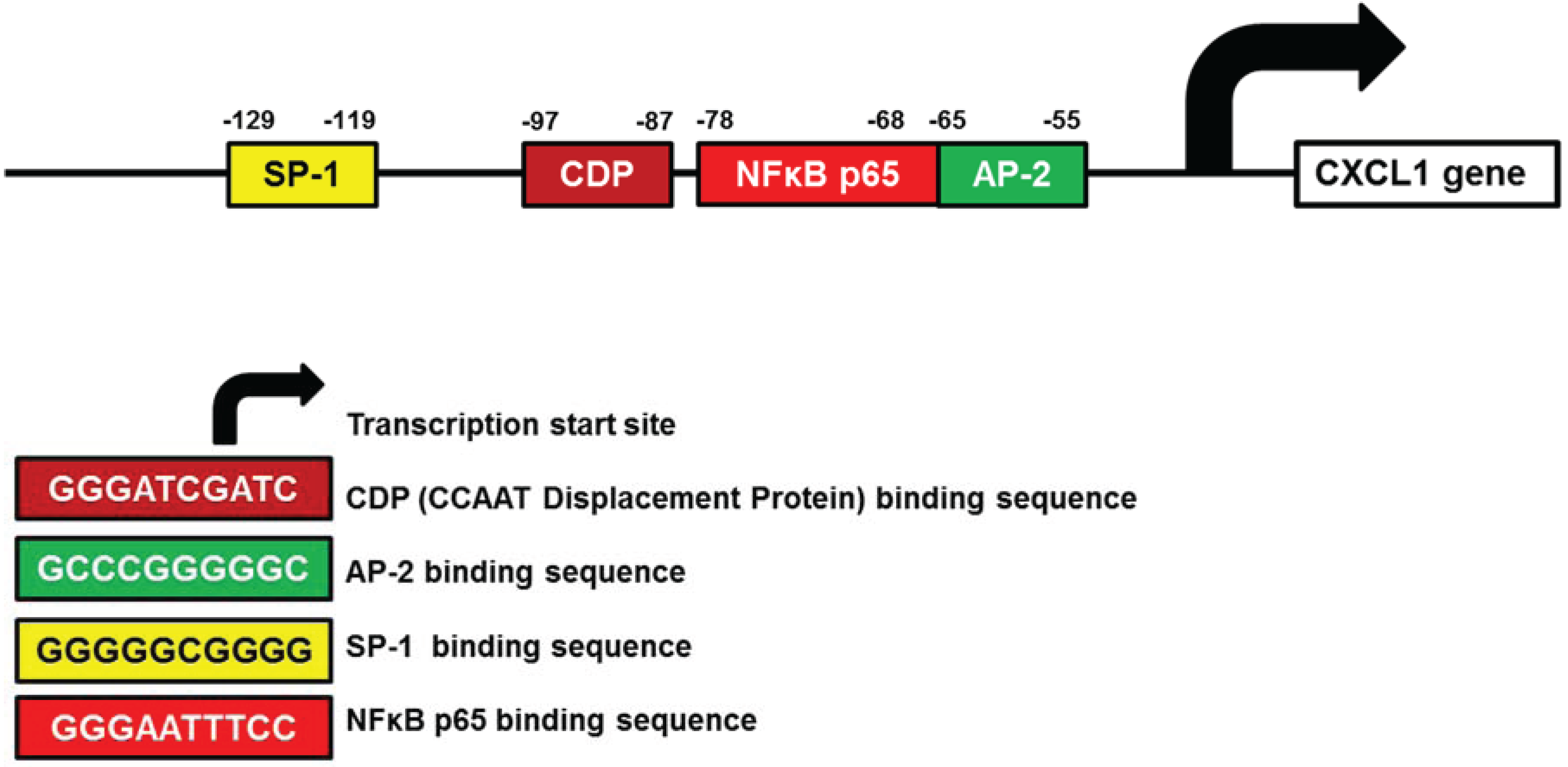
3.2. CXCL1

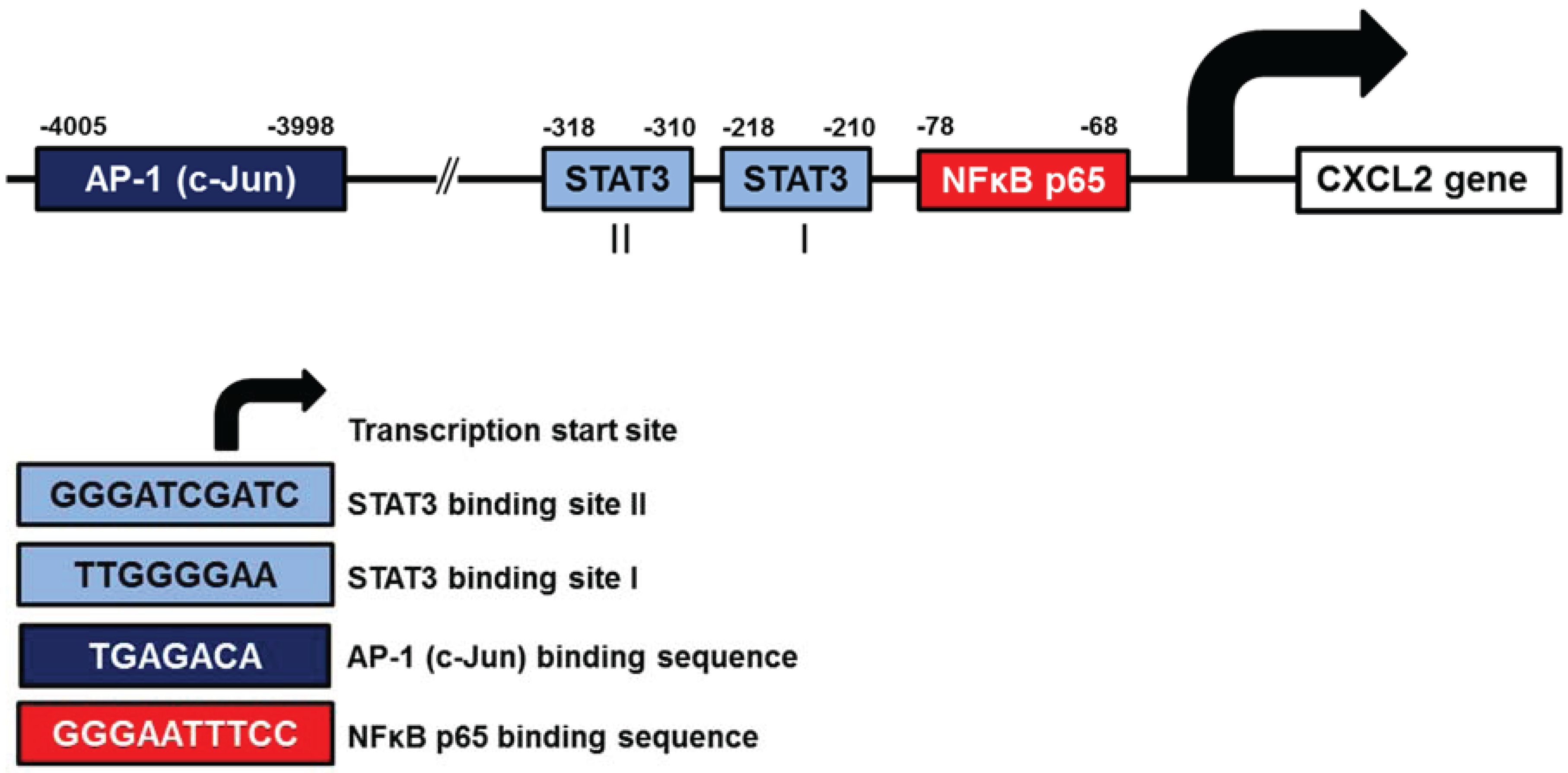
3.3. CXCL2

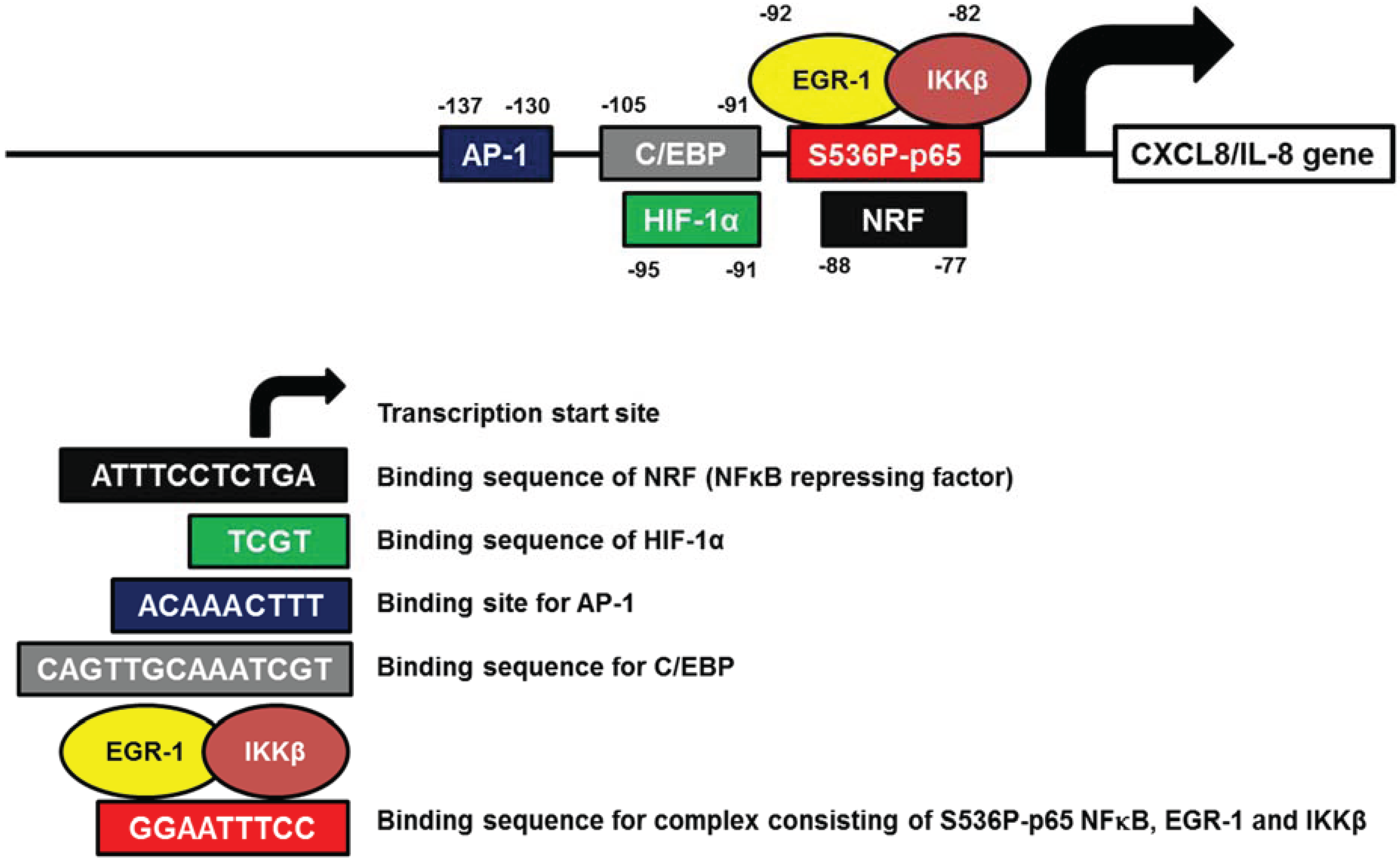
3.4. CXCL8

4. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Baggiolini, M.; Walz, A.; Kunkel, S.L. Neutrophil-activating peptide-1/interleukin 8, a novel cytokine that activates neutrophils. J. Clin. Investig. 1989, 84, 1045–1049. [Google Scholar] [CrossRef]
- Kunkel, S.L.; Strieter, R.M.; Chensue, S.W.; Basha, M.; Standiford, T.; Ham, J.; Remick, D.G. Tumor necrosis factor-alpha, interleukin-8 and chemotactic cytokines. Prog. Clin. Biol. Res. 1990, 349, 433–444. [Google Scholar] [PubMed]
- Baggiolini, M.; Dewald, B.; Moser, B. Human chemokines: An update. Annu. Rev. Immunol. 1997, 15, 675–705. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.M. Chemokines and the molecular basis of cancer metastasis. N. Engl. J. Med. 2001, 345, 833–835. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A. Chemokines and cancer. Int. J. Cancer 2006, 119, 2026–2029. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Lazennec, G.; Richmond, A. Chemokines and chemokine receptors: New insights into cancer-related inflammation. Trends Mol. Med. 2010, 16, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Rainczuk, A.; Rao, J.; Gathercole, J.; Stephens, A.N. The emerging role of CXC chemokines in epithelial ovarian cancer. Reproduction 2012, 144, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Yoshie, O. Chemokines: A new classification system and their role in immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.J.; Lolis, E. Structure, function, and inhibition of chemokines. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Yoshie, O. The chemokine superfamily revisited. Immunity 2012, 36, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Fidler, I.J. Interleukin 8: An autocrine growth factor for human ovarian cancer. Oncol. Res. 2000, 12, 97–106. [Google Scholar] [PubMed]
- Szlosarek, P.; Balkwill, F. The inflammatory cytokine network of epithelial cancer: Therapeutic implications. Novartis Found. Symp. 2004, 256, 227–237. [Google Scholar] [PubMed]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [PubMed]
- Sarvaiya, P.J.; Guo, D.; Ulasov, I.; Gabikian, P.; Lesniak, M.S. Chemokines in tumor progression and metastasis. Oncotarget 2013, 4, 2171–2185. [Google Scholar] [PubMed]
- Muralidhar, G.G.; Barbolina, M.V. Chemokine receptors in epithelial ovarian cancer. Int. J. Mol. Sci. 2013, 15, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Negus, R.P.; Stamp, G.W.; Relf, M.G.; Burke, F.; Malik, S.T.; Bernasconi, S.; Allavena, P.; Sozzani, S.; Mantovani, A.; Balkwill, F.R. The detection and localization of monocyte chemoattractant protein-1 (MCP-1) in human ovarian cancer. J. Clin. Investig. 1995, 95, 2391–2396. [Google Scholar] [CrossRef] [PubMed]
- Milliken, D.; Scotton, C.; Raju, S.; Balkwill, F.; Wilson, J. Analysis of chemokines and chemokine receptor expression in ovarian cancer ascites. Clin. Cancer Res. 2002, 8, 1108–1114. [Google Scholar] [PubMed]
- Negus, R.P.; Stamp, G.W.; Hadley, J.; Balkwill, F.R. Quantitative assessment of the leukocyte infiltrate in ovarian cancer and its relationship to the expression of C-C chemokines. Am. J. Pathol. 1997, 150, 1723–1734. [Google Scholar] [PubMed]
- Levina, V.; Nolen, B.M.; Marrangoni, A.M.; Cheng, P.; Marks, J.R.; Szczepanski, M.J.; Szajnik, M.E.; Gorelik, E.; Lokshin, A.E. Role of eotaxin-1 signaling in ovarian cancer. Clin. Cancer Res. 2009, 15, 2647–2656. [Google Scholar] [CrossRef] [PubMed]
- Nolen, B.M.; Lokshin, A.E. Targeting CCL11 in the treatment of ovarian cancer. Expert Opin. Ther. Target. 2010, 14, 157–167. [Google Scholar] [CrossRef]
- Singh, R.; Stockard, C.R.; Grizzle, W.E.; Lillard, J.W., Jr.; Singh, S. Expression and histopathological correlation of CCR9 and CCL25 in ovarian cancer. Int. J. Oncol. 2011, 39, 373–381. [Google Scholar] [PubMed]
- Facciabene, A.; Peng, X.; Hagemann, I.S.; Balint, K.; Barchetti, A.; Wang, L.; Gimotty, P.A.; Gilks, C.B.; Lal, P.; Zhang, L. Tumour hypoxia promotes tolerance and angiogenesis via CCL28 and Treg cells. Nature 2011, 475, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.; Swaby, R.F.; Liang, Y.; Yu, S.; Liu, S.; Lu, K.H.; Bast, R.C., Jr.; Mills, G.B.; Fang, X. Lysophosphatidic acid is a major regulator of growth-regulated oncogene alpha in ovarian cancer. Cancer Res. 2006, 66, 2740–2748. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Rosen, D.G.; Zhang, Z.; Bast, R.C., Jr.; Mills, G.B.; Colacino, J.A.; Mercado-Uribe, I.; Liu, J. The chemokine growth-regulated oncogene 1 (Gro-1) links RAS signaling to the senescence of stromal fibroblasts and ovarian tumorigenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 16472–16477. [Google Scholar] [CrossRef] [PubMed]
- Son, D.; Parl, A.K.; Rice, V.M.; Khabele, D. Keratinocyte chemoattractant (KC)/human growth-regulated oncogene (GRO) chemokines and pro-inflammatory chemokine networks in mouse and human ovarian epithelial cancer cells. Cancer Biol. Ther. 2007, 6, 1302–1312. [Google Scholar] [PubMed]
- Kavandi, L.; Collier, M.A.; Nguyen, H.; Syed, V. Progesterone and calcitriol attenuate inflammatory cytokines CXCL1 and CXCL2 in ovarian and endometrial cancer cells. J. Cell Biochem. 2012, 113, 3143–3152. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.F.; Schuerer-Maly, C.C.; Lofquist, A.K.; van Haaften-Day, C.; Ting, J.P.; White, C.M.; Martin, B.K.; Haskill, J.S. Taxol-dependent transcriptional activation of IL-8 expression in a subset of human ovarian cancer. Cancer Res. 1996, 56, 1303–1308. [Google Scholar] [PubMed]
- Xu, L.; Xie, K.; Mukaida, N.; Matsushima, K.; Fidler, I.J. Hypoxia-induced elevation in interleukin-8 expression by human ovarian carcinoma cells. Cancer Res. 1999, 59, 5822–5829. [Google Scholar] [PubMed]
- Zou, W.; Machelon, V.; Coulomb-L’Hermin, A.; Borvak, J.; Nome, F.; Isaeva, T.; Wei, S.; Krzysiek, R.; Durand-Gasselin, I.; Gordon, A.; et al. Stromal-derived factor-1 in human tumors recruits and alters the function of plasmacytoid precursor dendritic cells. Nat. Med. 2001, 7, 1339–1346. [Google Scholar] [CrossRef]
- Scotton, C.J.; Wilson, J.L.; Scott, K.; Stamp, G.; Wilbanks, G.D.; Fricker, S.; Bridger, G.; Balkwill, F.R. Multiple actions of the chemokine CXCL12 on epithelial tumor cells in human ovarian cancer. Cancer Res. 2002, 62, 5930–5938. [Google Scholar] [PubMed]
- Guo, L.; Cui, Z.M.; Zhang, J.; Huang, Y. Chemokine axes CXCL12/CXCR4 and CXCL16/CXCR6 correlate with lymph node metastasis in epithelial ovarian carcinoma. Chin. J. Cancer 2011, 30, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Gooden, M.J.; Wiersma, V.R.; Boerma, A.; Leffers, N.; Boezen, H.M.; ten Hoor, K.A.; Hollema, H.; Walenkamp, A.M.; Daemen, T.; Nijman, H.W.; et al. Elevated serum CXCL16 is an independent predictor of poor survival in ovarian cancer and may reflect pro-metastatic ADAM protease activity. Br. J. Cancer 2014, 110, 1535–1544. [Google Scholar] [CrossRef]
- Gaudin, F.; Nasreddine, S.; Donnadieu, A.C.; Emilie, D.; Combadière, C.; Prévot, S.; Machelon, V.; Balabanian, K. Identification of the chemokine CX3CL1 as a new regulator of malignant cell proliferation in epithelial ovarian cancer. PLOS ONE 2011, 6, e21546. [Google Scholar] [CrossRef]
- Kim, M.; Rooper, L.; Xie, J.; Rayahin, J.; Burdette, J.E.; Kajdacsy-Balla, A.A.; Barbolina, M.V. The lymphotactin receptor is expressed in epithelial ovarian carcinoma and contributes to cell migration and proliferation. Mol. Cancer Res. 2012, 10, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Robinson, J.B.; Deguzman, A.; Bucana, C.D.; Fidler, I.J. Blockade of NFκB signaling inhibits angiogenesis and tumorigenicity of human ovarian cancer cells by suppressing expression of VEGF and IL-8. Cancer Res. 2000, 60, 5334–5339. [Google Scholar] [PubMed]
- Mabuchi, S.; Ohmichi, M.; Nishio, Y.; Hayasaka, T.; Kimura, A.; Ohta, T.; Kawagoe, J.; Takahashi, K.; Yada-Hashimoto, N.; Seino-Noda, H.; et al. Inhibition of NFκB increases the efficacy of cisplatin in in vitro and in vivo ovarian cancer models. J. Biol. Chem. 2004, 279, 23477–23485. [Google Scholar] [CrossRef]
- Annunziata, C.M.; Stavnes, H.T.; Kleinberg, L.; Berner, A.; Hernandez, L.F.; Birrer, M.J.; Steinberg, S.M.; Davidson, B.; Kohn, E.C. NFκB transcription factors are coexpressed and convey a poor outcome in ovarian cancer. Cancer 2010, 116, 3276–3284. [Google Scholar] [CrossRef] [PubMed]
- Leizer, A.L.; Alvero, A.B.; Fu, H.H.; Holmberg, J.C.; Cheng, Y.; Silasi, D.; Rutherford, T.; Mor, G. Regulation of inflammation by the NFκB pathway in ovarian cancer stem cells. Am. J. Reprod. Immunol. 2011, 65, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Singha, B.; Gatla, H.R.; Manna, S.; Chang, T.P.; Sanacora, S.; Poltoratsky, V.; Vancura, A.; Vancurova, I. Proteasome inhibition increases recruitment of IκB kinase β (IKKβ), S536P-p65, and transcription factor EGR1 to interleukin-8 (IL-8) promoter, resulting in increased IL-8 production in ovarian cancer cells. J. Biol. Chem. 2014, 289, 2687–2700. [Google Scholar] [CrossRef] [PubMed]
- Mabuchi, S.; Ohmichi, M.; Nishio, Y.; Hayasaka, T.; Kimura, A.; Ohta, T.; Kawagoe, J.; Takahashi, K.; Yada-Hashimoto, N.; Seino-Noda, H.; et al. Inhibition of inhibitor of NFκB phosphorylation increases the efficacy of paclitaxel in in vitro and in vivo ovarian cancer models. Clin. Cancer Res. 2004, 10, 7645–7654. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Alvero, A.; Silasi, D.; Kelly, M.; Fest, S.; Visintin, I.; Leiser, A.; Schwartz, P.; Rutherford, T.; Mor, G. Regulation of IKKβ by miR-199a affects NFκB activity in ovarian cancer cells. Oncogene 2008, 27, 4712–4723. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, L.; Hsu, S.C.; Davidson, B.; Birrer, M.J.; Kohn, E.C.; Annunziata, C.M. Activation of NFκB signaling by inhibitor of NFκB kinase beta increases aggressiveness of ovarian cancer. Cancer Res. 2010, 70, 4005–4014. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.; Kim, M.; Hernandez, L.; Grajales, V.; Noonan, A.; Anver, M.; Davidson, B.; Annunziata, C.M. IKK-ε coordinates invasion and metastasis of ovarian cancer. Cancer Res. 2012, 72, 5494–5504. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in NFκB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed]
- Smale, S.T. Dimer-specific regulatory mechanisms within the NFκB family of transcription factors. Immunol. Rev. 2012, 246, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Natoli, G. NFκB and chromatin: Ten years on the path from basic mechanisms to candidate drugs. Immunol. Rev. 2012, 246, 183–92. [Google Scholar] [CrossRef] [PubMed]
- Strait, K.A.; Warnick, C.T.; Ford, C.D.; Dabbas, B.; Hammond, E.H.; Ilstrup, S.J. Histone deacetylase inhibitors induce G2-checkpoint arrest and apoptosis in cisplatinum-resistant ovarian cancer cells associated with overexpression of the Bcl-2-related protein Bad. Mol. Cancer Ther. 2005, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.J.; Holson, E.; Wagner, F.; Zhang, Y.; Fass, D.M.; Haggarty, S.J.; Bhaskara, S.; Hiebert, S.W.; Schreiber, S.L.; Khabele, D. The DNA damage mark pH2AX differentiates the cytotoxic effects of small molecule HDAC inhibitors in ovarian cancer cells. Cancer Biol. Ther. 2011, 12, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Zhou, H.; Li, J.; Tipton, T.; Wang, B.; Shao, G.; Gilbert, E.N.; Li, Q.; Jiang, S. Preclinical studies on histone deacetylase inhibitors as therapeutic reagents for endometrial and ovarian cancers. Future Oncol. 2011, 7, 1415–1428. [Google Scholar] [CrossRef] [PubMed]
- Khabele, D. The therapeutic potential of class I selective histone deacetylase inhibitors in ovarian cancer. Front. Oncol. 2014. [Google Scholar] [CrossRef]
- Ashburner, B.P.; Westerheide, S.D.; Baldwin, A.S., Jr. The p65 (RelA) subunit of NFκB interacts with the histone deacetylase (HDAC) corepressors HDAC1 and HDAC2 to negatively regulate gene expression. Mol. Cell Biol. 2001, 21, 7065–7077. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Gilmour, P.S.; Jimenez, L.A.; MacNee, W. Oxidative stress and TNFα induce histone acetylation and NFκB/AP-1 activation in alveolar epithelial cells: Potential mechanism in gene transcription in lung inflammation. Mol. Cell Biochem. 2002, 234–235, 239–248. [Google Scholar]
- Tomita, K.; Barnes, P.; Adcock, I. The effect of oxidative stress on histone acetylation and IL-8 release. Biochem. Biophys. Res. Commun. 2003, 301, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Mayo, M.W.; Denlinger, C.E.; Broad, R.M.; Yeung, F.; Reilly, E.T.; Shi, Y.; Jones, D.R. Ineffectiveness of histone deacetylase inhibitors to induce apoptosis involves the transcriptional activation of NFκB through the Akt pathway. J. Biol. Chem. 2003, 278, 18980–18989. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.R.; Chida, A.S.; Bauter, M.R.; Shafiq, N.; Seweryniak, K.; Maggirwar, S.B.; Kilty, I.; Rahman, I. Cigarette smoke induces proinflammatory cytokine release by activation of NFκB and posttranslational modifications of histone deacetylase in macrophages. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 291, L46–L57. [Google Scholar] [CrossRef]
- Ziesche, E.; Kettner-Buhrow, D.; Weber, A.; Wittwer, T.; Jurida, L.; Soelch, J.; Muller, H.; Newel, D.; Kronich, P.; Schneider, H.; et al. The coactivator role of histone deacetylase 3 in IL-1-signaling involves deacetylation of p65 NFκB. Nucl. Acids Res. 2013, 41, 90–109. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Sengupta, S.; Berk, M.; Kwak, Y.G.; Escobar, P.F.; Belinson, J.; Mok, S.C.; Xu, Y. Hypoxia enhances lysophosphatidic acid responsiveness in ovarian cancer cells and lysophosphatidic acid induces ovarian tumor metastasis in vivo. Cancer Res. 2006, 66, 7983–7990. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; de Marzo, A.M.; Laughner, E.; Lim, M.; Hilton, D.A.; Zagzag, D.; Buechler, P.; Isaacs, W.B.; Semenza, G.L.; Simons, J.W. Overexpression of hypoxia-inducible factor 1α in common human cancers and their metastases. Cancer Res. 1999, 59, 5830–5835. [Google Scholar] [PubMed]
- Birner, P.; Schindl, M.; Obermair, A.; Breitenecker, G.; Oberhuber, G. Expression of hypoxia-inducible factor 1α in epithelial ovarian tumors: Its impact on prognosis and on response to chemotherapy. Clin. Cancer Res. 2001, 7, 1661–1668. [Google Scholar] [PubMed]
- Braicu, E.I.; Luketina, H.; Richter, R.; Castillo-Tong, D.C.; Lambrechts, S.; Mahner, S.; Concin, N.; Mentze, M.; Zeillinger, R.; Vergote, I. HIF1α is an independent prognostic factor for overall survival in advanced primary epithelial ovarian cancer—A study of the OVCAD Consortium. Oncol. Targets Ther. 2014, 7, 1563–1569. [Google Scholar] [CrossRef]
- Kryczek, I.; Lange, A.; Mottram, P.; Alvarez, X.; Cheng, P.; Hogan, M.; Moons, L.; Wei, S.; Zou, L.; Machelon, V.; et al. CXCL12 and vascular endothelial growth factor synergistically induce neoangiogenesis in human ovarian cancers. Cancer Res. 2005, 65, 465–472. [Google Scholar] [PubMed]
- Koong, A.C.; Chen, E.Y.; Giaccia, A.J. Hypoxia causes the activation of NFκB through the phosphorylation of IκBα on tyrosine residues. Cancer Res. 1994, 54, 1425–1430. [Google Scholar] [PubMed]
- Culver, C.; Sundqvist, A.; Mudie, S.; Melvin, A.; Xirodimas, D.; Rocha, S. Mechanism of hypoxia-induced NFκB. Mol. Cell Biol. 2010, 30, 4901–4921. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Rajagopal, V.; Gonsalves, C.; Johnson, C.; Kalra, V.K. A novel role of hypoxia-inducible factor in cobalt chloride- and hypoxia-mediated expression of IL-8 chemokine in human endothelial cells. J. Immunol. 2006, 177, 7211–7224. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Pouyssegur, J. Tumor cell metabolism: Cancer’s Achilles’ heel. Cancer Cell 2008, 13, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.P.; Sabatini, D.M. Cancer cell metabolism: Warburg and beyond. Cell 2008, 134, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J. Clin. Investig. 2013, 123, 3664–3671. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Fidler, I.J. Acidic pH-induced elevation in interleukin 8 expression by human ovarian carcinoma cells. Cancer Res. 2000, 60, 4610–4616. [Google Scholar] [PubMed]
- Vegran, F.; Boidot, R.; Michiels, C.; Sonveaux, P.; Feron, O. Lactate influx through the endothelial cell monocarboxylate transporter MCT1 supports an NFκB/IL-8 pathway that drives tumor angiogenesis. Cancer Res. 2011, 71, 2550–2560. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.S.; Roberts, P.C.; Frisard, M.I.; McMillan, R.P.; Brown, T.J.; Lawless, M.H.; Hulver, M.W.; Schmelz, E.M. Metabolic changes during ovarian cancer progression as targets for sphingosine treatment. Exp. Cell Res. 2013, 319, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Caneba, C.; Yang, L.; Baddour, J.; Curtis, R.; Win, J.; Hartig, S.; Marini, J.; Nagrath, D. Nitric oxide is a positive regulator of the Warburg effect in ovarian cancer cells. Cell Death Dis. 2014, 5, e1302. [Google Scholar] [CrossRef]
- Kellenberger, L.D.; Bruin, J.E.; Greenaway, J.; Campbell, N.E.; Moorehead, R.A.; Holloway, A.C.; Petrik, J. The role of dysregulated glucose metabolism in epithelial ovarian cancer. J. Oncol. 2010. [Google Scholar] [CrossRef]
- Gallagher, E.J.; LeRoith, D. Diabetes, cancer, and metformin: Connections of metabolism and cell proliferation. Ann. NY Acad. Sci. 2011, 1243, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Hursting, S.D.; Dunlap, S.M.; Ford, N.A.; Hursting, M.J.; Lashinger, L.M. Calorie restriction and cancer prevention: A mechanistic perspective. Cancer Metable 2013. [Google Scholar] [CrossRef]
- Al-Wahab, Z.; Tebbe, C.; Chhina, J.; Dar, S.A.; Morris, R.T.; Ali-Fehmi, R.; Giri, S.; Munkarah, A.R.; Rattan, R. Dietary energy balance modulates ovarian cancer progression and metastasis. Oncotarget 2014, 5, 6063–6075. [Google Scholar] [PubMed]
- Shimazu, T.; Hirschey, M.D.; Newman, J.; He, W.; Shirakawa, K.; le Moan, N.; Grueter, C.A.; Lim, H.; Saunders, L.R.; Stevens, R.D.; et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science 2013, 339, 211–214. [Google Scholar] [CrossRef] [PubMed]
- De Visser, K.E.; Jonkers, J. Towards understanding the role of cancer-associated inflammation in chemoresistance. Curr. Pharm. Des. 2009, 15, 1844–1853. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.F.; Haskill, J.S.; Mukaida, N.; Matsushima, K.; Ting, J.P. Identification of tumor-specific paclitaxel (Taxol)-responsive regulatory elements in theinterleukin-8 promoter. Mol. Cell Biol. 1997, 17, 5097–5105. [Google Scholar] [PubMed]
- Aghajanian, C. Clinical update: Novel targets in gynecologic malignancies. Semin. Oncol. 2004, 31, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.G.; Alvero, A.B.; Chen, R.; Silasi, D.A.; Abrahams, V.M.; Chan, S.; Visintin, I.; Rutherford, T.; Mor, G. TLR-4 signaling promotes tumor growth and paclitaxel chemoresistance in ovarian cancer. Cancer Res. 2006, 66, 3859–3868. [Google Scholar] [CrossRef] [PubMed]
- Szajnik, M.; Szczepanski, M.J.; Czystowska, M.; Elishaev, E.; Mandapathil, M.; Nowak-Markwitz, E.; Spaczynski, M.; Whiteside, T.L. TLR4 signaling induced by lipopolysaccharide or paclitaxel regulates tumor survival and chemoresistance in ovarian cancer. Oncogene 2009, 28, 4353–4363. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.; Man, S.; Elliott, P.; Adams, J.; Kerbel, R.S. Lack of multicellular drug resistance observed in human ovarian and prostate carcinoma treated with the proteasome inhibitor PS-341. Clin. Cancer Res. 2000, 6, 3719–3728. [Google Scholar] [PubMed]
- Aghajanian, C.; Dizon, D.S.; Sabbatini, P.; Raizer, J.J.; Dupont, J.; Spriggs, D.R. Phase I trial of bortezomib and carboplatin in recurrent ovarian or primary peritoneal cancer. J. Clin. Oncol. 2005, 23, 5943–5949. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, P.T.; Landen, C.N., Jr.; Coleman, R.L.; Milam, M.R.; Levenback, C.; Johnston, T.A.; Gershenson, D.M. Phase I trial of the proteasome inhibitor bortezomib in combination with carboplatin in patients with platinum- and taxane-resistant ovarian cancer. Gynecol. Oncol. 2008, 108, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Aghajanian, C.; Blessing, J.A.; Darcy, K.M.; Reid, G.; deGeest, K.; Rubin, S.C.; Mannel, R.S.; Rotmensch, J.; Schilder, R.J.; Riordan, W. A phase II evaluation of bortezomib in the treatment of recurrent platinum-sensitive ovarian or primary peritoneal cancer: A Gynecologic Oncology Group study. Gynecol. Oncol. 2009, 115, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Jandial, D.D.; Farshchi-Heydari, S.; Larson, C.A.; Elliott, G.I.; Wrasidlo, W.J.; Howell, S.B. Enhanced delivery of cisplatin to intraperitoneal ovarian carcinomas mediated by the effects of bortezomib on the human copper transporter 1. Clin. Cancer Res. 2009, 15, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.B.; Safaei, R.; Larson, C.A.; Sailor, M.J. Copper transporters and the cellular pharmacology of the platinum-containing cancer drugs. Mol. Pharmacol. 2010, 77, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Messeguer, X.; Escudero, R.; Farre, D.; Nunez, O.; Martinez, J.; Alba, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Farre, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Rosello, L.; Alba, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef] [PubMed]
- Negus, R.P.; Turner, L.; Burke, F.; Balkwill, F.R. Hypoxia down-regulates MCP-1 expression: Implications for macrophage distribution in tumors. J. Leukoc. Biol. 1998, 63, 758–765. [Google Scholar] [PubMed]
- Sica, A.; Saccani, A.; Bottazzi, B.; Bernasconi, S.; Allavena, P.; Gaetano, B.; Fei, F.; LaRosa, G.; Scotton, C.; Balkwill, F.; et al. Defective expression of the monocyte chemotactic protein-1 receptor CCR2 in macrophages associated with human ovarian carcinoma. J. Immunol. 2000, 164, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Wojnarowicz, P.; Gambaro, K.; de Ladurantaye, M.; Quinn, M.C.; Provencher, D.; Mes-Masson, A.M.; Tonin, P.N. Overexpressing the CCL2 chemokine in an epithelial ovarian cancer cell line results in latency of in vivo tumourigenicity. Oncogenesis 2012, 1, e27. [Google Scholar] [CrossRef] [PubMed]
- Fader, A.N.; Rasool, N.; Vaziri, S.A.; Kozuki, T.; Faber, P.W.; Elson, P.; Biscotti, C.V.; Michener, C.M.; Rose, P.G.; Rojas-Espaillat, L.; et al. CCL2 expression in primary ovarian carcinoma is correlated with chemotherapy response and survival outcomes. Anticancer Res. 2010, 12, 4791–4798. [Google Scholar]
- Arnold, J.M.; Huggard, P.R.; Cummings, M.; Ramm, G.A.; Chenevix-Trench, G. Reduced expression of chemokine (C-C motif) ligand-2 (CCL2) in ovarian adenocarcinoma. Br. J. Cancer 2005, 92, 2024–2031. [Google Scholar] [CrossRef] [PubMed]
- Moisan, F.; Francisco, E.B.; Brozovic, A.; Duran, G.E.; Wang, Y.C.; Chaturvedi, S.; Seetharam, S.; Snyder, L.A.; Doshi, P.; Sikic, B.I. Enhancement of paclitaxel and carboplatin therapies by CCL2 blockade in ovarian cancers. Mol. Oncol. 2014, 8, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Soeda, S.; Kiko, Y.; Suzuki, O.; Hashimoto, Y.; Watanabe, T.; Nishiyama, H.; Tasaki, K.; Hojo, H.; Abe, M.; et al. MCP-1 promotes invasion and adhesion of human ovarian cancer cells. Anticancer Res. 2013, 33, 4785–4790. [Google Scholar] [PubMed]
- Szlosarek, P.W.; Grimshaw, M.J.; Kulbe, H.; Wilson, J.L.; Wilbanks, G.D.; Burke, F.; Balkwill, F.R. Expression and regulation of tumor necrosis factor alpha in normal and malignant ovarian epithelium. Mol. Cancer Ther. 2006, 5, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Silver, D.L.; Naora, H.; Liu, J.; Cheng, W.; Montell, D.J. Activated signal transducer and activator of transcription (STAT) 3: Localization in focal adhesions and function in ovarian cancer cell motility. Cancer Res. 2004, 64, 3550–3558. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guo, A.; Yu, J.; Possemato, A.; Chen, Y.; Zheng, W.; Polakiewicz, R.D.; Kinzler, K.W.; Vogelstein, B.; Velculescu, V.E.; et al. Identification of STAT3 as a substrate of receptor protein tyrosine phosphatase T. Proc. Natl. Acad. Sci. USA 2007, 104, 4060–4064. [Google Scholar] [CrossRef] [PubMed]
- Coward, J.; Kulbe, H.; Chakravarty, P.; Leader, D.; Vassileva, V.; Leinster, D.A.; Thompson, R.; Schioppa, T.; Nemeth, J.; Vermeulen, J.; et al. Interleukin-6 as a therapeutic target in human ovarian cancer. Clin. Cancer Res. 2011, 17, 6083–6096. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, K.; Komohara, Y.; Tashiro, H.; Ohtake, H.; Nakagawa, T.; Katabuchi, H.; Takeya, M. Involvement of M2-polarized macrophages in the ascites from advanced epithelial ovarian carcinoma in tumor progression via Stat3 activation. Cancer Sci. 2010, 101, 2128–2136. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, A.M.; Clarke, D.L.; Bradbury, D.A.; Corbett, L.M.; Patel, J.A.; Knox, A.J. Transcriptional regulation of monocyte chemotactic protein-1 release by endothelin-1 in human airway smooth muscle cells involves NFκB and AP-1. Br. J. Pharmacol. 2009, 157, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.Y.; Chang, S.C.; Wu, H.Y.; Yu, T.C.; Wei, W.C.; Lin, S.; Chien, C.L.; Chang, M.F. Upregulation of the chemokine (C-C motif) ligand 2 via a severe acute respiratory syndrome coronavirus spike-ACE2 signaling pathway. J. Virol. 2010, 84, 7703–7712. [Google Scholar] [CrossRef] [PubMed]
- Dragomir, E.; Manduteanu, I.; Calin, M.; Gan, A.M.; Stan, D.; Koenen, R.R.; Weber, C.; Simionescu, M. High glucose conditions induce upregulation of fractalkine and monocyte chemotactic protein-1 in human smooth muscle cells. Thromb. Haemost. 2008, 100, 1155–1165. [Google Scholar] [PubMed]
- Mojsilovic-Petrovic, J.; Callaghan, D.; Cui, H.; Dean, C.; Stanimirovic, D.B.; Zhang, W. Hypoxia-inducible factor-1 (HIF-1) is involved in the regulation of hypoxia-stimulated expression of monocyte chemoattractant protein-1 (MCP-1/CCL2) and MCP-5 (Ccl12) in astrocytes. J. Neuroinflam. 2007. [Google Scholar] [CrossRef]
- Elsharkawy, A.M.; Oakley, F.; Lin, F.; Packham, G.; Mann, D.A.; Mann, J. The NFκB p50:p50:HDAC-1 repressor complex orchestrates transcriptional inhibition of multiple pro-inflammatory genes. J. Hepatol. 2010, 53, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, S.; Yamada, Y.; Liu, F.; Murohara, T.; Itoh, K.; Yamamoto, M.; Ichihara, G. Ablation of the transcription factor Nrf2 promotes ischemia-induced neovascularization by enhancing the inflammatory response. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Toubal, A.; Clement, K.; Fan, R.; Ancel, P.; Pelloux, V.; Rouault, C.; Veyrie, N.; Hartemann, A.; Treuter, E.; Venteclef, N. SMRT-GPS2 corepressor pathway dysregulation coincides with obesity-linked adipocyte inflammation. J. Clin. Investig. 2013, 123, 362–379. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Kabir, S.M.; Lee, E.; Son, D. CXCR2-driven ovarian cancer progression involves upregulation of proinflammatory chemokines by potentiating NFκB Activation via EGFR-transactivated Akt signaling. PLOS ONE 2013, 8, e83789. [Google Scholar] [CrossRef]
- Son, D.S.; Kabir, S.M.; Dong, Y.; Lee, E.; Adunyah, S.E. Characteristics of chemokine signatures elicited by EGF and TNF in ovarian cancer cells. J. Inflamm. 2013. [Google Scholar] [CrossRef]
- Nirodi, C.; Hart, J.; Dhawan, P.; Moon, N.S.; Nepveu, A.; Richmond, A. The role of CDP in the negative regulation of CXCL1 gene expression. J. Biol. Chem. 2001, 276, 26122–26131. [Google Scholar] [CrossRef] [PubMed]
- Amiri, K.; Ha, H.; Smulson, M.; Richmond, A. Differential regulation of CXC ligand 1 transcription in melanoma cell lines by poly(ADP-ribose) polymerase-1. Oncogene 2006, 25, 7714–7722. [Google Scholar] [CrossRef] [PubMed]
- Son, D.; Kabir, S.M.; Dong, Y.; Lee, E.; Adunyah, S.E. Inhibitory effect of tumor suppressor p53 on proinflammatory chemokine expression in ovarian cancer cells by reducing proteasomal degradation of IκB. PLOS ONE 2012, 7, e51116. [Google Scholar] [CrossRef]
- Yoneda, J.; Kuniyasu, H.; Crispens, M.A.; Price, J.E.; Bucana, C.D.; Fidler, I.J. Expression of angiogenesis-related genes and progression of human ovarian carcinomas in nude mice. J. Natl. Cancer Inst. 1998, 90, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Merritt, W.M.; Lin, Y.G.; Spannuth, W.A.; Fletcher, M.S.; Kamat, A.A.; Han, L.Y.; Landen, C.N.; Jennings, N.; de Geest, K.; Langley, R.R.; et al. Effect of interleukin-8 gene silencing with liposome-encapsulated small interfering RNA on ovarian cancer cell growth. J. Natl. Cancer Inst. 2008, 100, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Pecot, C.V.; Rupaimoole, R.; Yang, D.; Akbani, R.; Ivan, C.; Lu, C.; Wu, S.; Han, H.; Shah, M.Y.; Rodriguez-Aguayo, C.; et al. Tumour angiogenesis regulation by the miR-200 family. Nat. Commun. 2013. [Google Scholar] [CrossRef]
- Mukaida, N.; Shiroo, M.; Matsushima, K. Genomic structure of the human monocyte-derived neutrophil chemotactic factor IL-8. J. Immunol. 1989, 143, 1366–1371. [Google Scholar] [PubMed]
- Mukaida, N.; Mahe, Y.; Matsushima, K. Cooperative interaction of nuclear factor-kappa B- and cis-regulatory enhancer binding protein-like factor binding elements in activating the interleukin-8 gene by pro-inflammatory cytokines. J. Biol. Chem. 1990, 265, 21128–21133. [Google Scholar] [PubMed]
- Kunsch, C.; Rosen, C.A. NFκB subunit-specific regulation of the interleukin-8 promoter. Mol. Cell Biol. 1993, 13, 6137–6146. [Google Scholar] [PubMed]
- Stein, B.; Baldwin, A.S., Jr. Distinct mechanisms for regulation of the interleukin-8 gene involve synergism and cooperativity between C/EBP and NFκB. Mol. Cell Biol. 1993, 13, 7191–7198. [Google Scholar] [PubMed]
- Matsusaka, T.; Fujikawa, K.; Nishio, Y.; Mukaida, N.; Matsushima, K.; Kishimoto, T.; Akira, S. Transcription factors NF-IL6 and NFκB synergistically activate transcription of the inflammatory cytokines, interleukin 6 and interleukin 8. Proc. Natl. Acad. Sci. USA 1993, 90, 10193–10197. [Google Scholar] [CrossRef] [PubMed]
- Kunsch, C.; Lang, R.K.; Rosen, C.A.; Shannon, M.F. Synergistic transcriptional activation of the IL-8 gene by NFκB p65 (RelA) and NF-IL-6. J. Immunol. 1994, 153, 153–164. [Google Scholar] [PubMed]
- Oliveira, I.C.; Mukaida, N.; Matsushima, K.; Vilcek, J. Transcriptional inhibition of the interleukin-8 gene by interferon is mediated by the NFκB site. Mol. Cell Biol. 1994, 14, 5300–5308. [Google Scholar] [PubMed]
- Yasumoto, K.; Okamoto, S.; Mukaida, N.; Murakami, S.; Mai, M.; Matsushima, K. Tumor necrosis factor alpha and interferon gamma synergistically induce interleukin 8 production in a human gastric cancer cell line through acting concurrently on AP-1 and NFκB-like binding sites of the interleukin 8 gene. J. Biol. Chem. 1992, 267, 22506–22511. [Google Scholar] [PubMed]
- Nourbakhsh, M.; Kalble, S.; Dorrie, A.; Hauser, H.; Resch, K.; Kracht, M. The NFκB repressing factor is involved in basal repression and interleukin (IL)-1-induced activation of IL-8 transcription by binding to a conserved NFκB-flanking sequence element. J. Biol. Chem. 2001, 276, 4501–4508. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, E.; Dittrich-Breiholz, O.; Holtmann, H.; Kracht, M. Multiple control of interleukin-8 gene expression. J. Leukoc. Biol. 2002, 72, 847–855. [Google Scholar] [PubMed]
- Sasaki, C.Y.; Barberi, T.J.; Ghosh, P.; Longo, D.L. Phosphorylation of RelA/p65 on serine 536 defines an IκBα -independent NFκB pathway. J. Biol. Chem. 2005, 280, 34538–34547. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, C.C.; Ramaswami, S.; Juvekar, A.; Vu, H.Y.; Galdieri, L.; Davidson, D.; Vancurova, I. Gene-specific repression of proinflammatory cytokines in stimulated human macrophages by nuclear IκBα. J. Immunol. 2010, 185, 3685–3693. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.; Singha, B.; Phyo, S.A.; Gatla, H.R.; Chang, T.P.; Sanacora, S.; Ramaswami, S.; Vancurova, I. Proteasome inhibition by bortezomib increases IL-8 expression in androgen-independent prostate cancer cells: The role of IKKα. J. Immunol. 2013, 191, 2837–2846. [Google Scholar] [CrossRef] [PubMed]
- Buss, H.; Dörrie, A.; Schmitz, M.L.; Hoffmann, E.; Resch, K.; Kracht, M. Constitutive and IL-1-inducible phosphorylation of p65 NFκB at serine 536 is mediated by multiple protein kinases including IκB kinase (IKK)-α, IKKβ, IKKε, TRAF family member-associated (TANK)-binding kinase 1 (TBK1), and an unknown kinase and couples p65 to TATA-binding protein associated factor II31-mediated IL-8 transcription. J. Biol. Chem. 2004, 279, 55633–55643. [Google Scholar] [CrossRef] [PubMed]
- Moreno, R.; Sobotzik, J.M.; Schultz, C.; Schmitz, M.L. Specification of the NFκB transcriptional response by p65 phosphorylation and TNF-induced nuclear translocation of IKK epsilon. Nucleic Acids Res. 2010, 38, 6029–6044. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Pathak, P.S.; Fukumura, D. Hypoxia-induced activation of p38 mitogen-activated protein kinase and phosphatidylinositol 3'-kinase signaling pathways contributes to expression of interleukin 8 in human ovarian carcinoma cells. Clin. Cancer Res. 2004, 10, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Yu, S.; Bast, R.C.; Liu, S.; Xu, H.J.; Hu, S.X.; LaPushin, R.; Claret, F.X.; Aggarwal, B.B.; Lu, Y.; et al. Mechanisms for lysophosphatidic acid-induced cytokine production in ovarian cancer cells. J. Biol. Chem. 2004, 279, 9653–9661. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, M.M.; Arevalo, J.M.; Armaiz-Pena, G.N.; Lu, C.; Stone, R.L.; Moreno-Smith, M.; Nishimura, M.; Lee, J.W.; Jennings, N.B.; Bottsford-Miller, J.; et al. Stress effects on FosB- and IL8-driven ovarian cancer growth and metastasis. J. Biol. Chem. 2010, 285, 35462–35470. [Google Scholar] [CrossRef] [PubMed]
- Bartels, M.; Schweda, A.T.; Dreikhausen, U.; Frank, R.; Resch, K.; Beil, W.; Nourbakhsh, M. Peptide-mediated disruption of NFκB/NRF interaction inhibits IL-8 gene activation by IL-1 or Helicobacter pylori. J. Immunol. 2007, 179, 7605–7613. [Google Scholar] [CrossRef] [PubMed]
- Hoberg, J.E.; Yeung, F.; Mayo, M.W. SMRT derepression by the IκB kinase alpha: A prerequisite to NFκB transcription and survival. Mol. Cell 2004, 16, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Nozell, S.; Laver, T.; Patel, K.; Benveniste, E.N. Mechanism of IFN-beta-mediated inhibition of IL-8 gene expression in astroglioma cells. J. Immunol. 2006, 177, 822–830. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singha, B.; Gatla, H.R.; Vancurova, I. Transcriptional Regulation of Chemokine Expression in Ovarian Cancer. Biomolecules 2015, 5, 223-243. https://doi.org/10.3390/biom5010223
Singha B, Gatla HR, Vancurova I. Transcriptional Regulation of Chemokine Expression in Ovarian Cancer. Biomolecules. 2015; 5(1):223-243. https://doi.org/10.3390/biom5010223
Chicago/Turabian StyleSingha, Bipradeb, Himavanth R. Gatla, and Ivana Vancurova. 2015. "Transcriptional Regulation of Chemokine Expression in Ovarian Cancer" Biomolecules 5, no. 1: 223-243. https://doi.org/10.3390/biom5010223