

Zinc Finger and BTB Domain-Containing 20: A Newly Emerging Player in Pathogenesis and Development of Human Cancers

Abstract
:1. Introduction
2. Functional Domains of ZBTB Proteins
2.1. The BTB Domain
2.2. The C2H2-Type ZF Domain
2.3. Other Known Domains
2.4. The Linker Region
2.5. ZBTB Family Members
3. Physiological Roles of ZBTB20
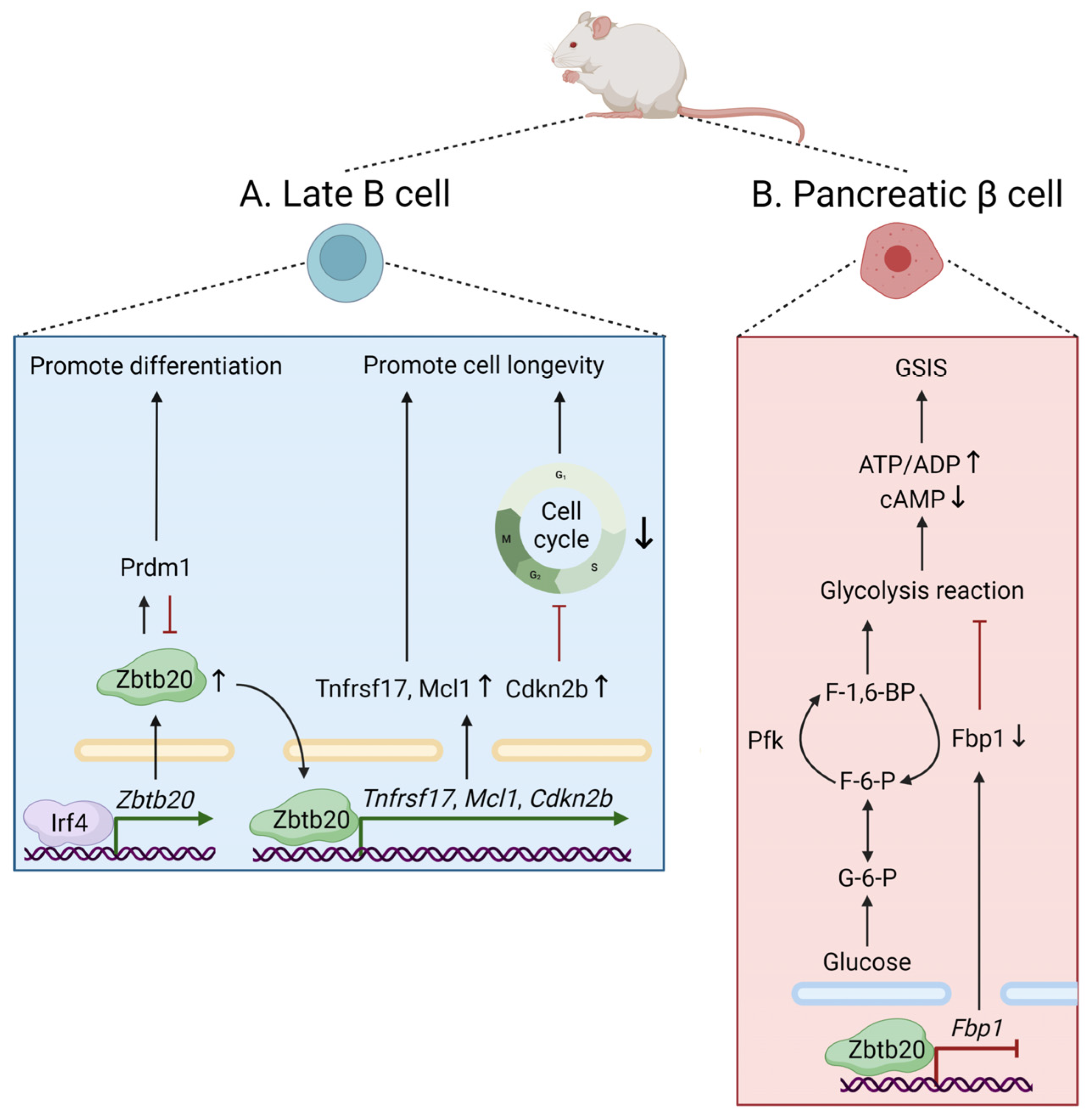
3.1. Lymphoid Development and Differentiation
3.2. Cellular Metabolism
3.3. Neurodevelopment
3.4. Immune Response and Inflammation
4. ZBTB20 in Cancers
4.1. Hepatocellular Carcinoma
4.2. Gastric Cancer
4.3. CNS Cancer
4.4. Blood Cancer
4.5. Other Cancers

{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Potential Function/Mechanism | Ref. | ||
|---|---|---|---|---|
| HCC | Mice | Transcriptional repression of the Afp gene | [78,79] | |
| (1) Promotion of liver regeneration (2) Regulation of the hepatic expression of Egfr | [80,117] | |||
| One of nineteen highly significant candidate locus implicated in mouse HCC | [82] | |||
| Humans | Reactivation of AFP via the miR-122-mediated regulation | [83] | ||
| An independent marker for poor prognosis in human HCC | [84,85] | |||
| Promotion of HCC by suppressing FOXO1 | [85] | |||
| Promotion of HCC by correlation with SETD7 | [86] | |||
| Suppression of HCV infection | [75] | |||
| Association with HBV integration frequency | [87] | |||
| GC | Humans | Identification of rs9841504 as a new susceptibility locus for non-cardia GC in the Chinese population | [88] | |
| Association of rs9841504 with severe intestinal metaplasia/atypical hyperplasia in the Chinese population | [89] | |||
| Identification of rs758277701 in the MSI subtype of GC in the Korean population | [92] | |||
| Identification of rs9288999 as a protective factor for reducing GC risk in the Chinese Han population | [93] | |||
| Promotion of GC via the NFKBIA/NF-κB signaling pathway | [94] | |||
| CNS cancer | Humans | Glioma | A mutation hotspot | [95,96] |
| Neuronal and mixed neuronal-glial tumors | Implication of miRNAs that target ZBTB20 in the classification of pediatric cases | [97] | ||
| Low-grade glioma | Identification of ZBTB20-AS4 as a critical IncRNA for predicting the prognosis | [98,99] | ||
| Shh-MB | Identification of the fusion transcripts of ZBTB20 as recurrent fusions | [100] | ||
| Glioblastoma | Promotion of glioblastoma through the miR-758-5p/ZBTB20 axis or by ZBTB20 | [101] | ||
| Blood cancer | Humans | B-CLL | The top differentially expressed gene in terms of VH mutation status | [102] |
| AML | Promotion of cell growth and migration via the LINC00641/miR-378a/ZBTB20 axis | [103] | ||
| Promotion of malignant phenotypes via the circ-SFMBT2/miR-582-3p/ZBTB20 axis | [104] | |||
| Promotion of leukemia development via the circ-0001602/miR-192-5p/ZBTB20 axis | [105] | |||
| MCL | (1) A novel downstream target repressed by BACH1 (2) Involvement in the BACH1-mediated regulation of tumor immune microenvironment | [106] | ||
| Others | Humans | BC | Downregulation in ERα+ BC biopsies upon treatment of aromatase inhibitors | [109] |
| Upregulation in ERα+ BC cell lines upon anacardic acid treatment | [110] | |||
| Promotion of cell migration and invasion via the SNHG8/miR-634/ZBTB20 axis | [111] | |||
| Promotion of cell proliferation, migration, and invasion via the circ-0104345/miR-876-3p/ZBTB20 axis | [112] | |||
| Colorectal cancer | Identification of rs10511330 and rs16822593 as two of the top 10 SNPs in patients | [113] | ||
| Cervical cancer | One of ten potential driver genes | [114] | ||
| NSCLC | (1) Upregulation in NSCLC tissues (2) Promotion of cell proliferation by repressing FOXO1 | [115] | ||
| Ovarian cancer | Increase in cells that migrate in omentum tissue pretreated with extracellular vesicles isolated from ascitic supernatant of high-grade patients | [116] | ||
5. Summary and Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Maeda, T. Regulation of hematopoietic development by ZBTB transcription factors. Int. J. Hematol. 2016, 104, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Mi, J.; Li, N.; Sui, L.; Wan, T.; Zhang, J.; Chen, T.; Cao, X. Identification and characterization of DPZF, a novel human BTB/POZ zinc finger protein sharing homology to BCL-6. Biochem. Biophys. Res. Commun. 2001, 282, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, A.P.; Zhang, H.; Zhang, Y.; Michaud, M.; Xie, Z.; Patti, M.E.; Grusby, M.J.; Zhang, W.J. Zinc finger protein Zbtb20 is essential for postnatal survival and glucose homeostasis. Mol. Cell. Biol. 2009, 29, 2804–2815. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, E.H.; Tonchev, A.B.; Stoykova, A.; Chowdhury, K. Regulation of archicortical arealization by the transcription factor Zbtb20. Hippocampus 2012, 22, 2144–2156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xie, Z.; Zhou, L.; Li, L.; Zhang, H.; Zhou, G.; Ma, X.; Herrera, P.L.; Liu, Z.; Grusby, M.J.; et al. The zinc finger protein ZBTB20 regulates transcription of fructose-1,6-bisphosphatase 1 and beta cell function in mice. Gastroenterology 2012, 142, 1571–1580.e6. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, P.; Bao, Y.; Han, Y.; Wang, Y.; Zhang, Q.; Zhan, Z.; Meng, J.; Li, Y.; Li, N.; et al. Zinc finger protein ZBTB20 promotes Toll-like receptor-triggered innate immune responses by repressing IκBα gene transcription. Proc. Natl. Acad. Sci. USA 2013, 110, 11097–11102. [Google Scholar] [CrossRef]
- Chevrier, S.; Emslie, D.; Shi, W.; Kratina, T.; Wellard, C.; Karnowski, A.; Erikci, E.; Smyth, G.K.; Chowdhury, K.; Tarlinton, D.; et al. The BTB-ZF transcription factor Zbtb20 is driven by Irf4 to promote plasma cell differentiation and longevity. J. Exp. Med. 2014, 211, 827–840. [Google Scholar] [CrossRef]
- Wang, Y.; Bhattacharya, D. Adjuvant-specific regulation of long-term antibody responses by ZBTB20. J. Exp. Med. 2014, 211, 841–856. [Google Scholar] [CrossRef]
- Sun, Y.; Preiss, N.K.; Valenteros, K.B.; Kamal, Y.; Usherwood, Y.K.; Frost, H.R.; Usherwood, E.J. Zbtb20 Restrains CD8 T Cell Immunometabolism and Restricts Memory Differentiation and Antitumor Immunity. J. Immunol. 2020, 205, 2649–2666. [Google Scholar] [CrossRef]
- Consortium, T.U. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2022, 51, D523–D531. [Google Scholar] [CrossRef]
- Bardwell, V.J.; Treisman, R. The POZ domain: A conserved protein-protein interaction motif. Genes Dev. 1994, 8, 1664–1677. [Google Scholar] [CrossRef]
- Stogios, P.J.; Downs, G.S.; Jauhal, J.J.; Nandra, S.K.; Prive, G.G. Sequence and structural analysis of BTB domain proteins. Genome Biol. 2005, 6, R82. [Google Scholar] [CrossRef]
- Perez-Torrado, R.; Yamada, D.; Defossez, P.A. Born to bind: The BTB protein-protein interaction domain. Bioessays 2006, 28, 1194–1202. [Google Scholar] [CrossRef]
- Bonchuk, A.; Denisov, S.; Georgiev, P.; Maksimenko, O. Drosophila BTB/POZ domains of “ttk group” can form multimers and selectively interact with each other. J. Mol. Biol. 2011, 412, 423–436. [Google Scholar] [CrossRef]
- Hong, S.H.; David, G.; Wong, C.W.; Dejean, A.; Privalsky, M.L. SMRT corepressor interacts with PLZF and with the PML-retinoic acid receptor α (RARα) and PLZF-RARα oncoproteins associated with acute promyelocytic leukemia. Proc. Natl. Acad. Sci. USA 1997, 94, 9028–9033. [Google Scholar] [CrossRef]
- Ahmad, K.F.; Melnick, A.; Lax, S.; Bouchard, D.; Liu, J.; Kiang, C.L.; Mayer, S.; Takahashi, S.; Licht, J.D.; Prive, G.G. Mechanism of SMRT corepressor recruitment by the BCL6 BTB domain. Mol. Cell 2003, 12, 1551–1564. [Google Scholar] [CrossRef]
- Seyfert, V.L.; Allman, D.; He, Y.; Staudt, L.M. Transcriptional repression by the proto-oncogene BCL-6. Oncogene 1996, 12, 2331–2342. [Google Scholar]
- Chang, C.C.; Ye, B.H.; Chaganti, R.S.; Dalla-Favera, R. BCL-6, a POZ/zinc-finger protein, is a sequence-specific transcriptional repressor. Proc. Natl. Acad. Sci. USA 1996, 93, 6947–6952. [Google Scholar] [CrossRef]
- Melnick, A.; Ahmad, K.F.; Arai, S.; Polinger, A.; Ball, H.; Borden, K.L.; Carlile, G.W.; Prive, G.G.; Licht, J.D. In-depth mutational analysis of the promyelocytic leukemia zinc finger BTB/POZ domain reveals motifs and residues required for biological and transcriptional functions. Mol. Cell. Biol. 2000, 20, 6550–6567. [Google Scholar] [CrossRef]
- Kang, M.I.; Kobayashi, A.; Wakabayashi, N.; Kim, S.G.; Yamamoto, M. Scaffolding of Keap1 to the actin cytoskeleton controls the function of Nrf2 as key regulator of cytoprotective phase 2 genes. Proc. Natl. Acad. Sci. USA 2004, 101, 2046–2051. [Google Scholar] [CrossRef]
- Pintard, L.; Willis, J.H.; Willems, A.; Johnson, J.L.; Srayko, M.; Kurz, T.; Glaser, S.; Mains, P.E.; Tyers, M.; Bowerman, B.; et al. The BTB protein MEL-26 is a substrate-specific adaptor of the CUL-3 ubiquitin-ligase. Nature 2003, 425, 311–316. [Google Scholar] [CrossRef]
- Xu, L.; Wei, Y.; Reboul, J.; Vaglio, P.; Shin, T.H.; Vidal, M.; Elledge, S.J.; Harper, J.W. BTB proteins are substrate-specific adaptors in an SCF-like modular ubiquitin ligase containing CUL-3. Nature 2003, 425, 316–321. [Google Scholar] [CrossRef]
- Geyer, R.; Wee, S.; Anderson, S.; Yates, J.; Wolf, D.A. BTB/POZ domain proteins are putative substrate adaptors for cullin 3 ubiquitin ligases. Mol. Cell 2003, 12, 783–790. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kang, M.I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Mol. Cell. Biol. 2004, 24, 7130–7139. [Google Scholar] [CrossRef]
- Chaharbakhshi, E.; Jemc, J.C. Broad-complex, tramtrack, and bric-a-brac (BTB) proteins: Critical regulators of development. Genesis 2016, 54, 505–518. [Google Scholar] [CrossRef]
- Cerchietti, L.C.; Ghetu, A.F.; Zhu, X.; Da Silva, G.F.; Zhong, S.; Matthews, M.; Bunting, K.L.; Polo, J.M.; Fares, C.; Arrowsmith, C.H.; et al. A small-molecule inhibitor of BCL6 kills DLBCL cells in vitro and in vivo. Cancer Cell 2010, 17, 400–411. [Google Scholar] [CrossRef]
- Sakamoto, K.; Sogabe, S.; Kamada, Y.; Sakai, N.; Asano, K.; Yoshimatsu, M.; Ida, K.; Imaeda, Y.; Sakamoto, J.I. Discovery of high-affinity BCL6-binding peptide and its structure-activity relationship. Biochem. Biophys. Res. Commun. 2017, 482, 310–316. [Google Scholar] [CrossRef]
- McCoull, W.; Cheung, T.; Anderson, E.; Barton, P.; Burgess, J.; Byth, K.; Cao, Q.; Castaldi, M.P.; Chen, H.; Chiarparin, E.; et al. Development of a Novel B-Cell Lymphoma 6 (BCL6) PROTAC To Provide Insight into Small Molecule Targeting of BCL6. ACS Chem. Biol. 2018, 13, 3131–3141. [Google Scholar] [CrossRef]
- Slabicki, M.; Yoon, H.; Koeppel, J.; Nitsch, L.; Roy Burman, S.S.; Di Genua, C.; Donovan, K.A.; Sperling, A.S.; Hunkeler, M.; Tsai, J.M.; et al. Small-molecule-induced polymerization triggers degradation of BCL6. Nature 2020, 588, 164–168. [Google Scholar] [CrossRef]
- Ai, Y.; Hwang, L.; MacKerell, A.D., Jr.; Melnick, A.; Xue, F. Progress toward B-Cell Lymphoma 6 BTB Domain Inhibitors for the Treatment of Diffuse Large B-Cell Lymphoma and Beyond. J. Med. Chem. 2021, 64, 4333–4358. [Google Scholar] [CrossRef]
- Zacharchenko, T.; Kalverda, A.P.; Wright, S.C. Structural basis of Apt48 inhibition of the BCL6 BTB domain. Structure 2022, 30, 396–407.e3. [Google Scholar] [CrossRef]
- Fedotova, A.A.; Bonchuk, A.N.; Mogila, V.A.; Georgiev, P.G. C2H2 Zinc Finger Proteins: The Largest but Poorly Explored Family of Higher Eukaryotic Transcription Factors. Acta Nat. 2017, 9, 47–58. [Google Scholar] [CrossRef]
- Schmitges, F.W.; Radovani, E.; Najafabadi, H.S.; Barazandeh, M.; Campitelli, L.F.; Yin, Y.; Jolma, A.; Zhong, G.; Guo, H.; Kanagalingam, T.; et al. Multiparameter functional diversity of human C2H2 zinc finger proteins. Genome Res. 2016, 26, 1742–1752. [Google Scholar] [CrossRef]
- Wolfe, S.A.; Nekludova, L.; Pabo, C.O. DNA recognition by Cys2His2 zinc finger proteins. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 183–212. [Google Scholar] [CrossRef]
- Tsuzuki, S.; Enver, T. Interactions of GATA-2 with the promyelocytic leukemia zinc finger (PLZF) protein, its homologue FAZF, and the t(11;17)-generated PLZF-retinoic acid receptor α oncoprotein. Blood 2002, 99, 3404–3410. [Google Scholar] [CrossRef]
- Guidez, F.; Howell, L.; Isalan, M.; Cebrat, M.; Alani, R.M.; Ivins, S.; Hormaeche, I.; McConnell, M.J.; Pierce, S.; Cole, P.A.; et al. Histone acetyltransferase activity of p300 is required for transcriptional repression by the promyelocytic leukemia zinc finger protein. Mol. Cell. Biol. 2005, 25, 5552–5566. [Google Scholar] [CrossRef]
- Donaldson, N.S.; Daniel, Y.; Kelly, K.F.; Graham, M.; Daniel, J.M. Nuclear trafficking of the POZ-ZF protein Znf131. Biochim. Biophys. Acta 2007, 1773, 546–555. [Google Scholar] [CrossRef]
- Reeves, R.; Nissen, M.S. The A.T-DNA-binding domain of mammalian high mobility group I chromosomal proteins. A novel peptide motif for recognizing DNA structure. J. Biol. Chem. 1990, 265, 8573–8582. [Google Scholar] [CrossRef]
- Huth, J.R.; Bewley, C.A.; Nissen, M.S.; Evans, J.N.; Reeves, R.; Gronenborn, A.M.; Clore, G.M. The solution structure of an HMG-I(Y)-DNA complex defines a new architectural minor groove binding motif. Nat. Struct. Biol. 1997, 4, 657–665. [Google Scholar] [CrossRef]
- Aktar, S.; Sasaki, H.; Unoki, M. Identification of ZBTB24 protein domains and motifs for heterochromatin localization and transcriptional activation. Genes Cells 2019, 24, 746–755. [Google Scholar] [CrossRef]
- Fedele, M.; Benvenuto, G.; Pero, R.; Majello, B.; Battista, S.; Lembo, F.; Vollono, E.; Day, P.M.; Santoro, M.; Lania, L.; et al. A novel member of the BTB/POZ family, PATZ, associates with the RNF4 RING finger protein and acts as a transcriptional repressor. J. Biol. Chem. 2000, 275, 7894–7901. [Google Scholar] [CrossRef]
- Dent, A.L.; Shaffer, A.L.; Yu, X.; Allman, D.; Staudt, L.M. Control of inflammation, cytokine expression, and germinal center formation by BCL-6. Science 1997, 276, 589–592. [Google Scholar] [CrossRef]
- Ye, B.H.; Cattoretti, G.; Shen, Q.; Zhang, J.; Hawe, N.; de Waard, R.; Leung, C.; Nouri-Shirazi, M.; Orazi, A.; Chaganti, R.S.; et al. The BCL-6 proto-oncogene controls germinal-centre formation and Th2-type inflammation. Nat. Genet. 1997, 16, 161–170. [Google Scholar] [CrossRef]
- Fukuda, T.; Yoshida, T.; Okada, S.; Hatano, M.; Miki, T.; Ishibashi, K.; Okabe, S.; Koseki, H.; Hirosawa, S.; Taniguchi, M.; et al. Disruption of the Bcl6 gene results in an impaired germinal center formation. J. Exp. Med. 1997, 186, 439–448. [Google Scholar] [CrossRef]
- Johnston, R.J.; Poholek, A.C.; DiToro, D.; Yusuf, I.; Eto, D.; Barnett, B.; Dent, A.L.; Craft, J.; Crotty, S. Bcl6 and Blimp-1 are reciprocal and antagonistic regulators of T follicular helper cell differentiation. Science 2009, 325, 1006–1010. [Google Scholar] [CrossRef]
- Nurieva, R.I.; Chung, Y.; Martinez, G.J.; Yang, X.O.; Tanaka, S.; Matskevitch, T.D.; Wang, Y.H.; Dong, C. Bcl6 mediates the development of T follicular helper cells. Science 2009, 325, 1001–1005. [Google Scholar] [CrossRef]
- Kovalovsky, D.; Uche, O.U.; Eladad, S.; Hobbs, R.M.; Yi, W.; Alonzo, E.; Chua, K.; Eidson, M.; Kim, H.J.; Im, J.S.; et al. The BTB-zinc finger transcriptional regulator PLZF controls the development of invariant natural killer T cell effector functions. Nat. Immunol. 2008, 9, 1055–1064. [Google Scholar] [CrossRef]
- Savage, A.K.; Constantinides, M.G.; Han, J.; Picard, D.; Martin, E.; Li, B.; Lantz, O.; Bendelac, A. The transcription factor PLZF directs the effector program of the NKT cell lineage. Immunity 2008, 29, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Jash, A.; Wang, Y.; Weisel, F.J.; Scharer, C.D.; Boss, J.M.; Shlomchik, M.J.; Bhattacharya, D. ZBTB32 Restricts the Duration of Memory B Cell Recall Responses. J. Immunol. 2016, 197, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, A.M.; Zawislak, C.L.; Nakayama, T.; Sun, J.C. The transcription factor Zbtb32 controls the proliferative burst of virus-specific natural killer cells responding to infection. Nat. Immunol. 2014, 15, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Licht, J.D.; Chomienne, C.; Goy, A.; Chen, A.; Scott, A.A.; Head, D.R.; Michaux, J.L.; Wu, Y.; DeBlasio, A.; Miller, W.H., Jr.; et al. Clinical and molecular characterization of a rare syndrome of acute promyelocytic leukemia associated with translocation (11;17). Blood 1995, 85, 1083–1094. [Google Scholar] [CrossRef]
- Cattoretti, G.; Pasqualucci, L.; Ballon, G.; Tam, W.; Nandula, S.V.; Shen, Q.; Mo, T.; Murty, V.V.; Dalla-Favera, R. Deregulated BCL6 expression recapitulates the pathogenesis of human diffuse large B cell lymphomas in mice. Cancer Cell 2005, 7, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Duy, C.; Hurtz, C.; Shojaee, S.; Cerchietti, L.; Geng, H.; Swaminathan, S.; Klemm, L.; Kweon, S.M.; Nahar, R.; Braig, M.; et al. BCL6 enables Ph+ acute lymphoblastic leukaemia cells to survive BCR-ABL1 kinase inhibition. Nature 2011, 473, 384–388. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, T.; Matthews, W.C.; Jackson, E.R.; Staudt, D.E.; Douglas, A.M.; Findlay, I.J.; Persson, M.L.; Duchatel, R.J.; Mannan, A.; Germon, Z.P.; et al. B-cell Lymphoma 6 (BCL6): From Master Regulator of Humoral Immunity to Oncogenic Driver in Pediatric Cancers. Mol. Cancer Res. 2022, 20, 1711–1723. [Google Scholar] [CrossRef] [PubMed]
- Barna, M.; Pandolfi, P.P.; Niswander, L. Gli3 and Plzf cooperate in proximal limb patterning at early stages of limb development. Nature 2005, 436, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Kohlhase, J.; Bohm, D.; Schweiger, B.; Hoffmann, D.; Heitmann, M.; Horsthemke, B.; Wieczorek, D. Biallelic loss of function of the promyelocytic leukaemia zinc finger (PLZF) gene causes severe skeletal defects and genital hypoplasia. J. Med. Genet. 2008, 45, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhou, M.; He, Q.; He, Z. Zbtb40 Deficiency Leads to Morphological and Phenotypic Abnormalities of Spermatocytes and Spermatozoa and Causes Male Infertility. Cells 2023, 12, 1264. [Google Scholar] [CrossRef] [PubMed]
- Buaas, F.W.; Kirsh, A.L.; Sharma, M.; McLean, D.J.; Morris, J.L.; Griswold, M.D.; de Rooij, D.G.; Braun, R.E. Plzf is required in adult male germ cells for stem cell self-renewal. Nat. Genet. 2004, 36, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, R.M.; Seandel, M.; Falciatori, I.; Rafii, S.; Pandolfi, P.P. Plzf regulates germline progenitor self-renewal by opposing mTORC1. Cell 2010, 142, 468–479. [Google Scholar] [CrossRef]
- Hu, S.; Fambrough, D.; Atashi, J.R.; Goodman, C.S.; Crews, S.T. The Drosophila abrupt gene encodes a BTB-zinc finger regulatory protein that controls the specificity of neuromuscular connections. Genes Dev. 1995, 9, 2936–2948. [Google Scholar] [CrossRef]
- Zhu, S.; Lin, S.; Kao, C.F.; Awasaki, T.; Chiang, A.S.; Lee, T. Gradients of the Drosophila Chinmo BTB-zinc finger protein govern neuronal temporal identity. Cell 2006, 127, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Ma, X.; Ji, W.; Zhou, G.; Lu, Y.; Xiang, Z.; Wang, Y.X.; Zhang, L.; Hu, Y.; Ding, Y.Q.; et al. Zbtb20 is essential for the specification of CA1 field identity in the developing hippocampus. Proc. Natl. Acad. Sci. USA 2010, 107, 6510–6515. [Google Scholar] [CrossRef] [PubMed]
- Mitchelmore, C.; Kjaerulff, K.M.; Pedersen, H.C.; Nielsen, J.V.; Rasmussen, T.E.; Fisker, M.F.; Finsen, B.; Pedersen, K.M.; Jensen, N.A. Characterization of two novel nuclear BTB/POZ domain zinc finger isoforms. Association with differentiation of hippocampal neurons, cerebellar granule cells, and macroglia. J. Biol. Chem. 2002, 277, 7598–7609. [Google Scholar] [CrossRef] [PubMed]
- Preiss, N.K.; Kamal, Y.; Wilkins, O.M.; Li, C.; Kolling, F.W.t.; Trask, H.W.; Usherwood, Y.K.; Cheng, C.; Frost, H.R.; Usherwood, E.J. Characterizing control of memory CD8 T cell differentiation by BTB-ZF transcription factor Zbtb20. Life Sci. Alliance 2023, 6, e202201683. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Long, J.; Huang, M.X.; Luo, P.Y.; Bian, Z.H.; Xu, Y.F.; Wang, C.B.; Yang, S.H.; Li, L.; Selmi, C.; et al. Characterization of Organ-Specific Regulatory B Cells Using Single-Cell RNA Sequencing. Front. Immunol. 2021, 12, 711980. [Google Scholar] [CrossRef] [PubMed]
- Krzyzanowska, A.K.; Haynes Ii, R.A.H.; Kovalovsky, D.; Lin, H.C.; Osorio, L.; Edelblum, K.L.; Corcoran, L.M.; Rabson, A.B.; Denzin, L.K.; Sant’Angelo, D.B. Zbtb20 identifies and controls a thymus-derived population of regulatory T cells that play a role in intestinal homeostasis. Sci. Immunol. 2022, 7, eabf3717. [Google Scholar] [CrossRef]
- Li, H.; Liu, G.; Wan, X.; Zhou, L.; Qin, Z.B.; Ma, X.H.; Su, K.; Liu, Y.J.; Yuan, J.; Wei, C.C.; et al. The zinc finger and BTB domain containing protein ZBTB20 regulates plasma triglyceride metabolism by repressing lipoprotein lipase gene transcription in hepatocytes. Hepatology 2022, 75, 1169–1180. [Google Scholar] [CrossRef]
- Cordeddu, V.; Redeker, B.; Stellacci, E.; Jongejan, A.; Fragale, A.; Bradley, T.E.; Anselmi, M.; Ciolfi, A.; Cecchetti, S.; Muto, V.; et al. Mutations in ZBTB20 cause Primrose syndrome. Nat. Genet. 2014, 46, 815–817. [Google Scholar] [CrossRef]
- Melis, D.; Carvalho, D.; Barbaro-Dieber, T.; Espay, A.J.; Gambello, M.J.; Gener, B.; Gerkes, E.; Hitzert, M.M.; Hove, H.B.; Jansen, S.; et al. Primrose syndrome: Characterization of the phenotype in 42 patients. Clin. Genet. 2020, 97, 890–901. [Google Scholar] [CrossRef]
- Nielsen, J.V.; Nielsen, F.H.; Ismail, R.; Noraberg, J.; Jensen, N.A. Hippocampus-like corticoneurogenesis induced by two isoforms of the BTB-zinc finger gene Zbtb20 in mice. Development 2007, 134, 1133–1140. [Google Scholar] [CrossRef]
- Tonchev, A.B.; Tuoc, T.C.; Rosenthal, E.H.; Studer, M.; Stoykova, A. Zbtb20 modulates the sequential generation of neuronal layers in developing cortex. Mol. Brain 2016, 9, 65. [Google Scholar] [CrossRef]
- Ren, A.; Zhang, H.; Xie, Z.; Ma, X.; Ji, W.; He, D.Z.; Yuan, W.; Ding, Y.Q.; Zhang, X.H.; Zhang, W.J. Regulation of hippocampus-dependent memory by the zinc finger protein Zbtb20 in mature CA1 neurons. J. Physiol. 2012, 590, 4917–4932. [Google Scholar] [CrossRef]
- Cao, D.; Ma, X.; Cai, J.; Luan, J.; Liu, A.J.; Yang, R.; Cao, Y.; Zhu, X.; Zhang, H.; Chen, Y.X.; et al. ZBTB20 is required for anterior pituitary development and lactotrope specification. Nat. Commun. 2016, 7, 11121. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Yan, X.; Ye, Y.; Han, L.; Ma, X.; Wang, T.; Cao, D.; Zhang, W.J. ZBTB20 Regulates Prolactin Expression and Lactotrope Function in Adult Mice. Endocrinology 2022, 163, bqac181. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pittman, K.J.; Barker, J.R.; Salinas, R.E.; Stanaway, I.B.; Williams, G.D.; Carroll, R.J.; Balmat, T.; Ingham, A.; Gopalakrishnan, A.M.; et al. An Atlas of Genetic Variation Linking Pathogen-Induced Cellular Traits to Human Disease. Cell Host Microbe 2018, 24, 308–323.e306. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Shi, M.; Qiu, J.; Shi, Z.; Wang, C.; Fu, Y.; Lin, C.; Zhang, L.; Tao, J.; Liu, C.; et al. ZBTB20 regulates cardiac allograft rejection through NFsmall ka, CyrillicB-mediated inflammation in mouse heart transplantation. Transpl. Immunol. 2022, 74, 101676. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, N.; Zhu, X.; Yang, L.; Ye, B.; Li, H.; Zhu, P.; Lu, T.; Tian, Y.; Fan, Z. Circular RNA circZbtb20 maintains ILC3 homeostasis and function via Alkbh5-dependent m(6)A demethylation of Nr4a1 mRNA. Cell Mol. Immunol. 2021, 18, 1412–1424. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Zhang, H.; Tsai, W.; Zhang, Y.; Du, Y.; Zhong, J.; Szpirer, C.; Zhu, M.; Cao, X.; Barton, M.C.; et al. Zinc finger protein ZBTB20 is a key repressor of α-fetoprotein gene transcription in liver. Proc. Natl. Acad. Sci. USA 2008, 105, 10859–10864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cao, D.; Zhou, L.; Zhang, Y.; Guo, X.; Li, H.; Chen, Y.; Spear, B.T.; Wu, J.W.; Xie, Z.; et al. ZBTB20 is a sequence-specific transcriptional repressor of α-fetoprotein gene. Sci. Rep. 2015, 5, 11979. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shi, J.H.; Jiang, H.; Wang, K.; Lu, J.Y.; Jiang, X.; Ma, X.; Chen, Y.X.; Ren, A.J.; Zheng, J.; et al. ZBTB20 regulates EGFR expression and hepatocyte proliferation in mouse liver regeneration. Cell Death Dis. 2018, 9, 462. [Google Scholar] [CrossRef]
- Natarajan, A.; Wagner, B.; Sibilia, M. The EGF receptor is required for efficient liver regeneration. Proc. Natl. Acad. Sci. USA 2007, 104, 17081–17086. [Google Scholar] [CrossRef]
- Keng, V.W.; Villanueva, A.; Chiang, D.Y.; Dupuy, A.J.; Ryan, B.J.; Matise, I.; Silverstein, K.A.; Sarver, A.; Starr, T.K.; Akagi, K.; et al. A conditional transposon-based insertional mutagenesis screen for genes associated with mouse hepatocellular carcinoma. Nat. Biotechnol. 2009, 27, 264–274. [Google Scholar] [CrossRef]
- Kojima, K.; Takata, A.; Vadnais, C.; Otsuka, M.; Yoshikawa, T.; Akanuma, M.; Kondo, Y.; Kang, Y.J.; Kishikawa, T.; Kato, N.; et al. MicroRNA122 is a key regulator of α-fetoprotein expression and influences the aggressiveness of hepatocellular carcinoma. Nat. Commun. 2011, 2, 338. [Google Scholar] [CrossRef]
- Wang, Q.; Tan, Y.X.; Ren, Y.B.; Dong, L.W.; Xie, Z.F.; Tang, L.; Cao, D.; Zhang, W.P.; Hu, H.P.; Wang, H.Y. Zinc finger protein ZBTB20 expression is increased in hepatocellular carcinoma and associated with poor prognosis. BMC Cancer 2011, 11, 271. [Google Scholar] [CrossRef]
- Kan, H.; Huang, Y.; Li, X.; Liu, D.; Chen, J.; Shu, M. Zinc finger protein ZBTB20 is an independent prognostic marker and promotes tumor growth of human hepatocellular carcinoma by repressing FoxO1. Oncotarget 2016, 7, 14336–14349. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, S.; Hu, J.; Yu, C.; He, M.; Cai, Z. Increased Expression of SETD7 Promotes Cell Proliferation by Regulating Cell Cycle and Indicates Poor Prognosis in Hepatocellular Carcinoma. PLoS ONE 2016, 11, e0154939. [Google Scholar] [CrossRef]
- He, Z.; Zhu, J.; Mo, J.; Zhao, H.; Chen, Q. HBV DNA integrates into upregulated ZBTB20 in patients with hepatocellular carcinoma. Mol. Med. Rep. 2020, 22, 380–386. [Google Scholar] [CrossRef]
- Shi, Y.; Hu, Z.; Wu, C.; Dai, J.; Li, H.; Dong, J.; Wang, M.; Miao, X.; Zhou, Y.; Lu, F.; et al. A genome-wide association study identifies new susceptibility loci for non-cardia gastric cancer at 3q13.31 and 5p13.1. Nat. Genet. 2011, 43, 1215–1218. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Dai, S.; Chen, W.; Xia, C.; Lu, L.; Dai, S.; Qi, J.; Wang, M.; Wang, M.; Zhou, L.; et al. Environmental factors, seven GWAS-identified susceptibility loci, and risk of gastric cancer and its precursors in a Chinese population. Cancer Med. 2017, 6, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Song, H.R.; Kim, H.N.; Kweon, S.S.; Choi, J.S.; Shim, H.J.; Cho, S.H.; Chung, I.J.; Park, Y.K.; Kim, S.H.; Choi, Y.D.; et al. Genetic variations in the PRKAA1 and ZBTB20 genes and gastric cancer susceptibility in a Korean population. Mol. Carcinog. 2013, 52 (Suppl. 1), E155–E160. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Gu, J.; Ajani, J.A.; Chang, D.W.; Wu, X.; Stroehlein, J.R. Genetic and intermediate phenotypic susceptibility markers of gastric cancer in Hispanic Americans: A case-control study. Cancer 2014, 120, 3040–3048. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Cho, M.Y.; Kim, J.; Kim, S.N.; Oh, S.C.; Lee, K.A. Profiling cancer-associated genetic alterations and molecular classification of cancer in Korean gastric cancer patients. Oncotarget 2017, 8, 69888–69905. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Xiao, K. Prediction of gastric cancer risk: Association between ZBTB20 genetic variance and gastric cancer risk in Chinese Han population. Biosci. Rep. 2020, 40, BSR20202102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, X.; Zhang, M.; Cheng, L.; Zhang, Y.; Wang, X. ZBTB20 promotes cell migration and invasion of gastric cancer by inhibiting IkappaBα to induce NF-kappaB activation. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3862–3872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cheng, W.; Ren, X.; Wang, Z.; Liu, X.; Li, G.; Han, S.; Jiang, T.; Wu, A. Tumor Purity as an Underlying Key Factor in Glioma. Clin. Cancer Res. 2017, 23, 6279–6291. [Google Scholar] [CrossRef]
- Huang, Y.; Gao, X.; Yang, E.; Yue, K.; Cao, Y.; Zhao, B.; Zhang, H.; Dai, S.; Zhang, L.; Luo, P.; et al. Top-down stepwise refinement identifies coding and noncoding RNA-associated epigenetic regulatory maps in malignant glioma. J. Cell. Mol. Med. 2022, 26, 2230–2250. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewska, M.; Gruszka, R.; Stawiski, K.; Fendler, W.; Kordacka, J.; Grajkowska, W.; Daszkiewicz, P.; Liberski, P.P.; Zakrzewski, K. Expression-based decision tree model reveals distinct microRNA expression pattern in pediatric neuronal and mixed neuronal-glial tumors. BMC Cancer 2019, 19, 544. [Google Scholar] [CrossRef]
- Xiang, Z.; Chen, X.; Lv, Q.; Peng, X. A Novel Inflammatory lncRNAs Prognostic Signature for Predicting the Prognosis of Low-Grade Glioma Patients. Front. Genet. 2021, 12, 697819. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, N.; Wu, J.; Gao, X.; Zhao, H.; Liu, Z.; Yan, X.; Dong, J.; Wang, F.; Ba, Y.; et al. 5-Methylcytosine Related LncRNAs Reveal Immune Characteristics, Predict Prognosis and Oncology Treatment Outcome in Lower-Grade Gliomas. Front. Immunol. 2022, 13, 844778. [Google Scholar] [CrossRef]
- Skowron, P.; Farooq, H.; Cavalli, F.M.G.; Morrissy, A.S.; Ly, M.; Hendrikse, L.D.; Wang, E.Y.; Djambazian, H.; Zhu, H.; Mungall, K.L.; et al. The transcriptional landscape of Shh medulloblastoma. Nat. Commun. 2021, 12, 1749. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, J.; Hui, X.; Wang, W.; Fang, D.; Ding, L. Mir-758-5p Suppresses Glioblastoma Proliferation, Migration and Invasion by Targeting ZBTB20. Cell. Physiol. Biochem. 2018, 48, 2074–2083. [Google Scholar] [CrossRef]
- Haslinger, C.; Schweifer, N.; Stilgenbauer, S.; Dohner, H.; Lichter, P.; Kraut, N.; Stratowa, C.; Abseher, R. Microarray gene expression profiling of B-cell chronic lymphocytic leukemia subgroups defined by genomic aberrations and VH mutation status. J. Clin. Oncol. 2004, 22, 3937–3949. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.H.; Yu, L.J. Long non-coding RNA LINC00641 promotes cell growth and migration through modulating miR-378a/ZBTB20 axis in acute myeloid leukemia. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7498–7509. [Google Scholar] [CrossRef]
- Chang, W.; Shang, Z.; Ming, X.; Wu, J.; Xiao, Y. Circ-SFMBT2 facilitates the malignant growth of acute myeloid leukemia cells by modulating miR-582-3p/ZBTB20 pathway. Histol. Histopathol. 2022, 37, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Deng, J.; Chen, C.; Ma, X.; Yu, L.; Chen, L. Circ_0001602 aggravates the progression of acute myeloid leukemia by regulating the miR-192-5p/ZBTB20 axis. Hematology 2023, 28, 2240133. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Feng, M.; Zhang, Z.; Liu, J.; Zhang, H. BACH1 loss exerts antitumor effects on mantle cell lymphoma cells via inducing a tumor-intrinsic innate immune response and cell cycle arrest. Mol. Cancer Res. 2023, 21, 1274–1287. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.J.; Millenson, M.M.; Padavic-Shaller, K.; Wang, H.; Rogatko, A.; Clyde, J.; Boyd, R.L.; Yeslow, G.; Halbherr, T.; Schilder, R.J.; et al. Phase II study of fludarabine and α-interferon in patients with low-grade non-Hodgkin’s lymphoma. Haematologica 2004, 89, 1484–1491. [Google Scholar] [PubMed]
- Dal Col, J.; Mastorci, K.; Fae, D.A.; Muraro, E.; Martorelli, D.; Inghirami, G.; Dolcetti, R. Retinoic acid/α-interferon combination inhibits growth and promotes apoptosis in mantle cell lymphoma through Akt-dependent modulation of critical targets. Cancer Res. 2012, 72, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Mackay, A.; Urruticoechea, A.; Dixon, J.M.; Dexter, T.; Fenwick, K.; Ashworth, A.; Drury, S.; Larionov, A.; Young, O.; White, S.; et al. Molecular response to aromatase inhibitor treatment in primary breast cancer. Breast Cancer Res. 2007, 9, R37. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.J.; Krishna, A.; Vittitow, S.L.; Alizadeh-Rad, N.; Muluhngwi, P.; Rouchka, E.C.; Klinge, C.M. Transcriptomic response of breast cancer cells to anacardic acid. Sci. Rep. 2018, 8, 8063. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Qiu, B.; Yang, X.J.; Tang, H.L.; Peng, S.J.; Yang, P.; Dong, Y.M.; Yang, L.; Bao, G.Q.; Zhao, H.D. LncRNA SNHG8 promotes cell migration and invasion in breast cancer cell through miR-634/ZBTB20 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11639–11649. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, A.; Wang, L.; Shi, F.; Lin, F.; Cui, H. A Novel circ_0104345/miR-876-3p/ZBTB20 Axis Regulates the Proliferation, Migration, Invasion, and Apoptosis of Breast Cancer Cells. Biochem. Genet. 2023, 61, 2548–2565. [Google Scholar] [CrossRef]
- Penney, M.E.; Parfrey, P.S.; Savas, S.; Yilmaz, Y.E. Associations of single nucleotide polymorphisms with mucinous colorectal cancer: Genome-wide common variant and gene-based rare variant analyses. Biomark. Res. 2018, 6, 17. [Google Scholar] [CrossRef]
- Xu, Y.; Luo, H.; Hu, Q.; Zhu, H. Identification of Potential Driver Genes Based on Multi-Genomic Data in Cervical Cancer. Front. Genet. 2021, 12, 598304. [Google Scholar] [CrossRef]
- Zhao, J.G.; Ren, K.M.; Tang, J. Zinc finger protein ZBTB20 promotes cell proliferation in non-small cell lung cancer through repression of FoxO1. FEBS Lett. 2014, 588, 4536–4542. [Google Scholar] [CrossRef]
- Mitra, A.; Yoshida-Court, K.; Solley, T.N.; Mikkelson, M.; Yeung, C.L.A.; Nick, A.; Lu, K.; Klopp, A.H. Extracellular vesicles derived from ascitic fluid enhance growth and migration of ovarian cancer cells. Sci. Rep. 2021, 11, 9149. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.Z.; Zhuang, P.Y.; Hei, Z.Y.; Lin, P.Y.; Chen, Z.S.; Liu, Y.B.; Quan, Z.W.; Tang, Z.H. ZBTB20 is involved in liver regeneration after partial hepatectomy in mouse. Hepatobiliary Pancreat. Dis. Int. 2014, 13, 48–54. [Google Scholar] [CrossRef] [PubMed]



| Physiological Role | Tissue/Cell Type | Potential Mechanism | Ref. | |
|---|---|---|---|---|
| Lymphoid development and differentiation | Mice | B1, GC B, mature BMPC | Promotion of plasma cell differentiation | [7] |
| B cell and plasma cell | Regulation of long-term antibody production and plasma cell persistence after alum-adjuvanted immunization | [8] | ||
| CD8+ T cell | (1) Regulation of mitochondrial metabolism and glycolysis (2) Restriction of memory CD8+ T cell differentiation (3) Restriction of anti-tumor immunity | [9] | ||
| Transcriptional and epigenetic regulation of memory CD8+ T cell differentiation | [64] | |||
| Breg cell | A potential marker gene for Breg cells | [65] | ||
| Intestinal Treg cell | Modulation of intestinal homeostasis | [66] | ||
| Humans | BM, LN, thymus, PBC, fetal liver | A certain role in hematopoiesis, immune responses, and oncogenesis due to its high homology to BCL6 | [2] | |
| Cellular metabolism | Mice | Liver | Regulation of transcriptional profiling of genes in glucose metabolism | [3] |
| Pancreatic β cell | Regulation of glucose sensing and insulin secretion by repressing Fbp1 | [5] | ||
| Hepatocytes | Regulation of plasma triglyceride metabolism by repressing transcription of lipoprotein lipase | [67] | ||
| Humans | Multisystem | Regulation of lipid and glucose metabolism as well as mitochondrial function | [68,69] | |
| Neurodevelopment | Mice | Hippocampal neuron, cerebellar granule neuron, macroglia | Characterization of two isoforms, Zbtb20(S) and Zbtb20(L), in CNS | [63] |
| Immature cortical neuron | Regulation of Hi-like cortical neurogenesis | [70] | ||
| Whole brain | Modulation of the sequential neurogenesis in developing cortex | [71] | ||
| Hippocampal neuron | Specification of CA1 field identity in the developing hippocampus | [62] | ||
| Regulation of Hi-dependent long-term memory in mature CA1 neurons | [72] | |||
| Anterior pituitary | Regulation of anterior pituitary development and lactotrope specification | [73] | ||
| Regulation of prolactin expression and lactotrope function | [74] | |||
| Immune response and inflammation | Mice | Macrophage | Full activation of TLR signaling and TLR-triggered innate immune response by suppressing the Nfkbia gene transcription | [6] |
| Spleen cell | Regulation of T cells involved in acute heart allograft rejection through the activation of NF-κB pathway | [76] | ||
| Lymphocyte | Identification of circZbtb20 as a regulator of ILC3 homeostasis and function | [77] | ||
| Humans | 528 lymphoblastoid cell lines | Regulation of multiple infection-related phenotypes | [75] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, H. Zinc Finger and BTB Domain-Containing 20: A Newly Emerging Player in Pathogenesis and Development of Human Cancers. Biomolecules 2024, 14, 192. https://doi.org/10.3390/biom14020192
Liu J, Zhang H. Zinc Finger and BTB Domain-Containing 20: A Newly Emerging Player in Pathogenesis and Development of Human Cancers. Biomolecules. 2024; 14(2):192. https://doi.org/10.3390/biom14020192
Chicago/Turabian StyleLiu, Jiangyuan, and Han Zhang. 2024. "Zinc Finger and BTB Domain-Containing 20: A Newly Emerging Player in Pathogenesis and Development of Human Cancers" Biomolecules 14, no. 2: 192. https://doi.org/10.3390/biom14020192