
The Role of Gli1+ Mesenchymal Stem Cells in Osteogenesis of Craniofacial Bone
 and
and
Abstract
:1. Introduction
1.1. Characteristics of Gli1+ MSCs
1.2. Mechnisms of Osteogenesis of Craniofacial Bone
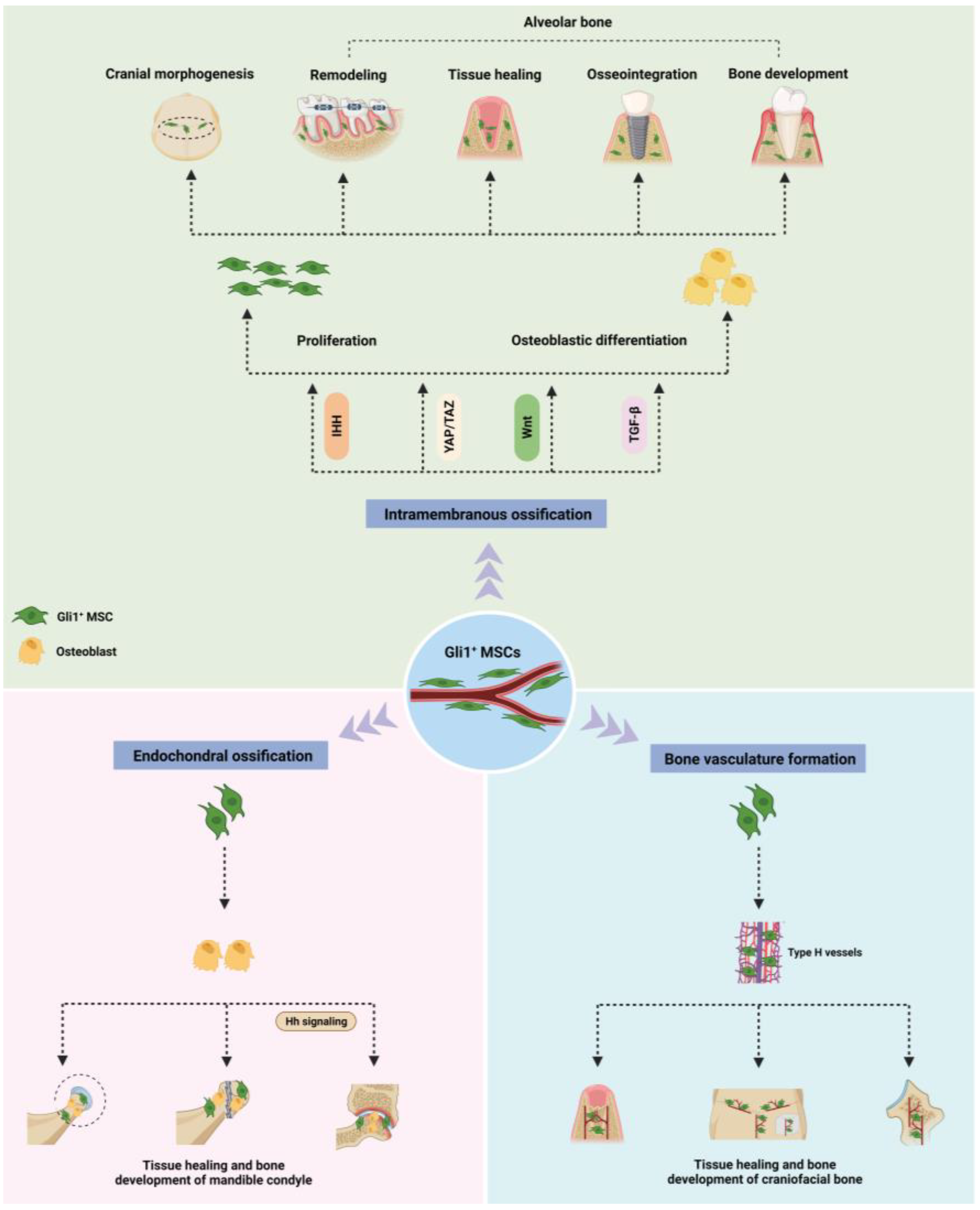
2. Role of Gli1+ MSC in Osteogenesis of Craniofacial Bones Formed through Intramembrane Ossification
2.1. Skull
2.2. Alveolar Bone
3. Roles of Gli1+ MSCs in Endochondral Ossification of Craniofacial Bones
4. Role of Gli1+ MSC in the Formation of Bone Vasculature
5. Age-Related Distribution in Gli1+ MSCs
6. The Plasticity of Gli1+ MSCs and the Strategies on Navigating Their Differentiation into the Osteogenic Lineage
6.1. The Overview of Gli1+ MSCs Plasticity
6.2. Strategies to Guide the Osteogenic Lineage Differentiation of Gli1+ MSCs
6.2.1. The Application of Osteo-Inductive Medium
6.2.2. Tissue Engineering
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kfoury, Y.; Scadden, D.T. Mesenchymal cell contributions to the stem cell niche. Cell Stem Cell 2015, 16, 239–253. [Google Scholar] [CrossRef]
- Peng, L.; Ye, L.; Zhou, X.D. Mesenchymal stem cells and tooth engineering. Int. J. Oral. Sci. 2009, 1, 6–12. [Google Scholar] [CrossRef]
- Asatrian, G.; Pham, D.; Hardy, W.R.; James, A.W.; Peault, B. Stem cell technology for bone regeneration: Current status and potential applications. Stem Cells Cloning 2015, 8, 39–48. [Google Scholar]
- Huang, Z.; Su, X.; Julaiti, M.; Chen, X.; Luan, Q. The role of PRX1-expressing cells in periodontal regeneration and wound healing. Front. Physiol. 2023, 14, 978640. [Google Scholar] [CrossRef]
- Alman, B.A. The role of hedgehog signalling in skeletal health and disease. Nat. Rev. Rheumatol. 2015, 11, 552–560. [Google Scholar] [CrossRef]
- Sabol, M.; Trnski, D.; Musani, V.; Ozretić, P.; Levanat, S. Role of GLI Transcription Factors in Pathogenesis and Their Potential as New Therapeutic Targets. Int. J. Mol. Sci. 2018, 19, 2562. [Google Scholar] [CrossRef] [PubMed]
- Hojo, H.; Ohba, S.; Yano, F.; Saito, T.; Ikeda, T.; Nakajima, K.; Komiyama, Y.; Nakagata, N.; Suzuki, K.; Takato, T.; et al. Gli1 protein participates in Hedgehog-mediated specification of osteoblast lineage during endochondral ossification. J. Biol. Chem. 2012, 287, 17860–17869. [Google Scholar] [CrossRef] [PubMed]
- Kesper, D.A.; Didt-Koziel, L.; Vortkamp, A. Gli2 activator function in preosteoblasts is sufficient to mediate Ihh-dependent osteoblast differentiation, whereas the repressor function of Gli2 is dispensable for endochondral ossification. Dev. Dyn. 2010, 239, 1818–1826. [Google Scholar] [CrossRef] [PubMed]
- Drakopoulou, E.; Outram, S.V.; Rowbotham, N.J.; Ross, S.E.; Furmanski, A.L.; Saldana, J.I.; Hager-Theodorides, A.L.; Crompton, T. Non-redundant role for the transcription factor Gli1 at multiple stages of thymocyte development. Cell Cycle 2010, 9, 4144–4152. [Google Scholar] [CrossRef]
- Park, H.L.; Bai, C.; Platt, K.A.; Matise, M.P.; Beeghly, A.; Hui, C.C.; Nakashima, M.; Joyner, A.L. Mouse Gli1 mutants are viable but have defects in SHH signaling in combination with a Gli2 mutation. Development 2000, 127, 1593–1605. [Google Scholar] [CrossRef]
- Huang, M.; Qing, Y.; Shi, Q.; Cao, Y.; Song, K. miR-342-3p elevates osteogenic differentiation of umbilical cord mesenchymal stem cells via inhibiting Sufu in vitro. Biochem. Biophys. Res. Commun. 2017, 491, 571–577. [Google Scholar] [CrossRef]
- Qing, Y.; Huang, M.; Cao, Y.; Du, T.; Song, K. Effects of miRNA-342-3p in modulating Hedgehog signaling pathway of human umbilical cord mesenchymal stem cells by down-regulating Sufu. Oral. Dis. 2019, 25, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Tian, Y.; Boogerd, C.J.; Lu, M.M.; Kadzik, R.S.; Stewart, K.M.; Evans, S.M.; Morrisey, E.E. Coordination of heart and lung co-development by a multipotent cardiopulmonary progenitor. Nature 2013, 500, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Degirmenci, B.; Valenta, T.; Dimitrieva, S.; Hausmann, G.; Basler, K. GLI1-expressing mesenchymal cells form the essential Wnt-secreting niche for colon stem cells. Nature 2018, 558, 449–453. [Google Scholar] [CrossRef]
- Dörner, J.; Martinez Rodriguez, V.; Ziegler, R.; Röhrig, T.; Cochran, R.S.; Götz, R.M.; Levin, M.D.; Pihlajoki, M.; Heikinheimo, M.; Wilson, D.B. GLI1(+) progenitor cells in the adrenal capsule of the adult mouse give rise to heterotopic gonadal-like tissue. Mol. Cell. Endocrinol. 2017, 441, 164–175. [Google Scholar] [CrossRef]
- Zhao, H.; Feng, J.; Ho, T.V.; Grimes, W.; Urata, M.; Chai, Y. The suture provides a niche for mesenchymal stem cells of craniofacial bones. Nat. Cell Biol. 2015, 17, 386–396. [Google Scholar] [CrossRef]
- Shi, Y.; He, G.; Lee, W.C.; McKenzie, J.A.; Silva, M.J.; Long, F. Gli1 identifies osteogenic progenitors for bone formation and fracture repair. Nat. Commun. 2017, 8, 2043. [Google Scholar] [CrossRef]
- Song, K.; Qing, Y.; Guo, Q.; Peden, E.K.; Chen, C.; Mitch, W.E.; Truong, L.; Cheng, J. PDGFRA in vascular adventitial MSCs promotes neointima formation in arteriovenous fistula in chronic kidney disease. JCI Insight 2020, 5, e137298. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, A.; Shalehin, N.; Takebe, H.; Shimo, T.; Irie, K. Sonic Hedgehog Signaling and Tooth Development. Int. J. Mol. Sci. 2020, 21, 1587. [Google Scholar] [CrossRef]
- Kramann, R.; Goettsch, C.; Wongboonsin, J.; Iwata, H.; Schneider, R.K.; Kuppe, C.; Kaesler, N.; Chang-Panesso, M.; Machado, F.G.; Gratwohl, S.; et al. Adventitial MSC-like Cells Are Progenitors of Vascular Smooth Muscle Cells and Drive Vascular Calcification in Chronic Kidney Disease. Cell Stem Cell 2016, 19, 628–642. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, X.; Yuan, W.; Liu, J.; Yang, W.; Zhu, Y.; Ye, C.; Xiong, X.; Zhang, Q.; Liu, J.; et al. Gli1+ Mesenchymal Stem Cells in Bone and Teeth. Curr. Stem Cell Res. Ther. 2022, 17, 494–502. [Google Scholar] [CrossRef]
- Bernhard, J.C.; Marolt Presen, D.; Li, M.; Monforte, X.; Ferguson, J.; Leinfellner, G.; Heimel, P.; Betti, S.L.; Shu, S.; Teuschl-Woller, A.H.; et al. Effects of Endochondral and Intramembranous Ossification Pathways on Bone Tissue Formation and Vascularization in Human Tissue-Engineered Grafts. Cells 2022, 11, 3070. [Google Scholar] [CrossRef]
- Yuan, Y.; Chai, Y. Regulatory mechanisms of jaw bone and tooth development. Curr. Top. Dev. Biol. 2019, 133, 91–118. [Google Scholar]
- Parada, C.; Chai, Y. Mandible and Tongue Development. Curr. Top. Dev. Biol. 2015, 115, 31–58. [Google Scholar] [PubMed]
- Maruyama, T.; Stevens, R.; Boka, A.; DiRienzo, L.; Chang, C.; Yu, H.I.; Nishimori, K.; Morrison, C.; Hsu, W. BMPR1A maintains skeletal stem cell properties in craniofacial development and craniosynostosis. Sci. Transl. Med. 2021, 13, eabb4416. [Google Scholar] [CrossRef]
- Tevlin, R.; McArdle, A.; Atashroo, D.; Walmsley, G.G.; Senarath-Yapa, K.; Zielins, E.R.; Paik, K.J.; Longaker, M.T.; Wan, D.C. Biomaterials for craniofacial bone engineering. J. Dent. Res. 2014, 93, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Aghali, A. Craniofacial Bone Tissue Engineering: Current Approaches and Potential Therapy. Cells 2021, 10, 2993. [Google Scholar] [CrossRef]
- Li, Y.; Fraser, D.; Mereness, J.; Van Hove, A.; Basu, S.; Newman, M.; Benoit, D.S.W. Tissue Engineered Neurovascularization Strategies for Craniofacial Tissue Regeneration. ACS Appl. Bio Mater. 2022, 5, 20–39. [Google Scholar] [CrossRef]
- Kusumbe, A.P.; Ramasamy, S.K.; Adams, R.H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 2014, 507, 323–328. [Google Scholar] [CrossRef]
- Filipowska, J.; Tomaszewski, K.A.; Niedźwiedzki, Ł.; Walocha, J.A.; Niedźwiedzki, T. The role of vasculature in bone development, regeneration and proper systemic functioning. Angiogenesis 2017, 20, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Ge, Q.; Fang, L.; Yu, H.; Zou, Z.; Zhang, P.; Lv, S.; Tong, P.; Xiao, L.; Chen, D.; et al. TGF-β/Smad2 signalling regulates enchondral bone formation of Gli1(+) periosteal cells during fracture healing. Cell Prolif. 2020, 53, e12904. [Google Scholar] [CrossRef]
- Jeffery, E.C.; Mann, T.L.A.; Pool, J.A.; Zhao, Z.; Morrison, S.J. Bone marrow and periosteal skeletal stem/progenitor cells make distinct contributions to bone maintenance and repair. Cell Stem Cell 2022, 29, 1547–1561.e6. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Stenberg, W.; Luo, W.; Feng, J.Q.; Zhao, H. Alveolar Bone Marrow Gli1+ Stem Cells Support Implant Osseointegration. J. Dent. Res. 2022, 101, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Feng, J.; Seidel, K.; Shi, S.; Klein, O.; Sharpe, P.; Chai, Y. Secretion of shh by a neurovascular bundle niche supports mesenchymal stem cell homeostasis in the adult mouse incisor. Cell Stem Cell 2014, 14, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Zhao, H.; Urata, M.; Chai, Y. Sutures Possess Strong Regenerative Capacity for Calvarial Bone Injury. Stem Cells Dev. 2016, 25, 1801–1807. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, Z.; Liu, P.; Wu, M.; Liu, A.Q.; Hu, C.; Liu, X.; Guo, H.; Yang, X.; Guo, X.; et al. Gli1(+) Cells Residing in Bone Sutures Respond to Mechanical Force via IP(3)R to Mediate Osteogenesis. Stem Cells Int. 2021, 2021, 8138374. [Google Scholar] [PubMed]
- Jing, D.; Chen, Z.; Men, Y.; Yi, Y.; Wang, Y.; Wang, J.; Yi, J.; Wan, L.; Shen, B.; Feng, J.Q.; et al. Response of Gli1(+) Suture Stem Cells to Mechanical Force Upon Suture Expansion. J. Bone Miner. Res. 2022, 37, 1307–1320. [Google Scholar] [CrossRef]
- Guo, Y.; Yuan, Y.; Wu, L.; Ho, T.V.; Jing, J.; Sugii, H.; Li, J.; Han, X.; Feng, J.; Guo, C.; et al. BMP-IHH-mediated interplay between mesenchymal stem cells and osteoclasts supports calvarial bone homeostasis and repair. Bone Res. 2018, 6, 30. [Google Scholar] [CrossRef]
- Yu, M.; Ma, L.; Yuan, Y.; Ye, X.; Montagne, A.; He, J.; Ho, T.V.; Wu, Y.; Zhao, Z.; Sta Maria, N.; et al. Cranial Suture Regeneration Mitigates Skull and Neurocognitive Defects in Craniosynostosis. Cell 2021, 184, 243–256.e18. [Google Scholar] [CrossRef]
- Xu, C.; Xie, X.; Zhao, H.; Wu, Y.; Wang, J.; Feng, J.Q. TGF-Beta Receptor II Is Critical for Osteogenic Progenitor Cell Proliferation and Differentiation During Postnatal Alveolar Bone Formation. Front. Physiol. 2021, 12, 721775. [Google Scholar] [CrossRef] [PubMed]
- Men, Y.; Wang, Y.; Yi, Y.; Jing, D.; Luo, W.; Shen, B.; Stenberg, W.; Chai, Y.; Ge, W.P.; Feng, J.Q.; et al. Gli1+ Periodontium Stem Cells Are Regulated by Osteocytes and Occlusal Force. Dev. Cell 2020, 54, 639–654.e6. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Takebe, H.; Mizoguchi, T.; Nakamura, H.; Iijima, M.; Irie, K.; Hosoya, A. Differentiation ability of Gli1(+) cells during orthodontic tooth movement. Bone 2023, 166, 116609. [Google Scholar] [CrossRef] [PubMed]
- Shalehin, N.; Seki, Y.; Takebe, H.; Fujii, S.; Mizoguchi, T.; Nakamura, H.; Yoshiba, N.; Yoshiba, K.; Iijima, M.; Shimo, T.; et al. Gli1(+)-PDL Cells Contribute to Alveolar Bone Homeostasis and Regeneration. J. Dent. Res. 2022, 101, 1537–1543. [Google Scholar] [CrossRef]
- Liu, A.Q.; Zhang, L.S.; Chen, J.; Sui, B.D.; Liu, J.; Zhai, Q.M.; Li, Y.J.; Bai, M.; Chen, K.; Jin, Y.; et al. Mechanosensing by Gli1(+) cells contributes to the orthodontic force-induced bone remodelling. Cell Prolif. 2020, 53, e12810. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lan, L.; Lei, J.; He, Y.; Zhang, Y. Gli1+ Osteogenic Progenitors Contribute to Condylar Development and Fracture Repair. Front. Cell Dev. Biol. 2022, 10, 819689. [Google Scholar] [CrossRef]
- Lei, J.; Chen, S.; Jing, J.; Guo, T.; Feng, J.; Ho, T.V.; Chai, Y. Inhibiting Hh Signaling in Gli1(+) Osteogenic Progenitors Alleviates TMJOA. J. Dent. Res. 2022, 101, 664–674. [Google Scholar] [CrossRef]
- Chen, J.; Li, M.; Liu, A.Q.; Zheng, C.X.; Bao, L.H.; Chen, K.; Xu, X.L.; Guan, J.T.; Bai, M.; Zhou, T.; et al. Gli1(+) Cells Couple with Type H Vessels and Are Required for Type H Vessel Formation. Stem Cell Rep. 2020, 15, 110–124. [Google Scholar] [CrossRef]
- Stanton, E.; Urata, M.; Chen, J.F.; Chai, Y. The clinical manifestations, molecular mechanisms and treatment of craniosynostosis. Dis. Model. Mech. 2022, 15, dmm049390. [Google Scholar] [CrossRef]
- Di Pietro, L.; Barba, M.; Prampolini, C.; Ceccariglia, S.; Frassanito, P.; Vita, A.; Guadagni, E.; Bonvissuto, D.; Massimi, L.; Tamburrini, G.; et al. GLI1 and AXIN2 Are Distinctive Markers of Human Calvarial Mesenchymal Stromal Cells in Nonsyndromic Craniosynostosis. Int. J. Mol. Sci. 2020, 21, 4356. [Google Scholar] [CrossRef]
- Li, B.; Wang, Y.; Fan, Y.; Ouchi, T.; Zhao, Z.; Li, L. Cranial Suture Mesenchymal Stem Cells: Insights and Advances. Biomolecules 2021, 11, 8. [Google Scholar] [CrossRef]
- Jiang, N.; Guo, W.; Chen, M.; Zheng, Y.; Zhou, J.; Kim, S.G.; Embree, M.C.; Songhee Song, K.; Marao, H.F.; Mao, J.J. Periodontal Ligament and Alveolar Bone in Health and Adaptation: Tooth Movement. Front. Oral Biol. 2016, 18, 1–8. [Google Scholar] [PubMed]
- Hollý, D.; Klein, M.; Mazreku, M.; Zamborský, R.; Polák, Š.; Danišovič, Ľ.; Csöbönyeiová, M. Stem Cells and Their Derivatives-Implications for Alveolar Bone Regeneration: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 11746. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ruan, J.; Weir, M.D.; Ren, K.; Schneider, A.; Wang, P.; Oates, T.W.; Chang, X.; Xu, H.H.K. Periodontal Bone-Ligament-Cementum Regeneration via Scaffolds and Stem Cells. Cells 2019, 8, 537. [Google Scholar] [CrossRef]
- Tavelli, L.; Barootchi, S.; Rasperini, G.; Giannobile, W.V. Clinical and patient-reported outcomes of tissue engineering strategies for periodontal and peri-implant reconstruction. Periodontology 2000 2022, 91, 217–269. [Google Scholar] [CrossRef]
- Gao, Z.H.; Hu, L.; Liu, G.L.; Wei, F.L.; Liu, Y.; Liu, Z.H.; Fan, Z.P.; Zhang, C.M.; Wang, J.S.; Wang, S.L. Bio-Root and Implant-Based Restoration as a Tooth Replacement Alternative. J. Dent. Res. 2016, 95, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Totaro, A.; Panciera, T.; Piccolo, S. YAP/TAZ upstream signals and downstream responses. Nat. Cell Biol. 2018, 20, 888–899. [Google Scholar] [CrossRef]
- Acri, T.M.; Shin, K.; Seol, D.; Laird, N.Z.; Song, I.; Geary, S.M.; Chakka, J.L.; Martin, J.A.; Salem, A.K. Tissue Engineering for the Temporomandibular Joint. Adv. Healthc. Mater. 2019, 8, e1801236. [Google Scholar] [CrossRef]
- de Souza, R.F.; Lovato da Silva, C.H.; Nasser, M.; Fedorowicz, Z.; Al-Muharraqi, M.A. Interventions for the management of temporomandibular joint osteoarthritis. Cochrane Database Syst. Rev. 2012, 2012, Cd007261. [Google Scholar]
- Song, K.; Rao, N.J.; Chen, M.L.; Huang, Z.J.; Cao, Y.G. Enhanced bone regeneration with sequential delivery of basic fibroblast growth factor and sonic hedgehog. Injury 2011, 42, 796–802. [Google Scholar] [CrossRef]
- Luo, W.; Yi, Y.; Jing, D.; Zhang, S.; Men, Y.; Ge, W.P.; Zhao, H. Investigation of Postnatal Craniofacial Bone Development with Tissue Clearing-Based Three-Dimensional Imaging. Stem Cells Dev. 2019, 28, 1310–1321. [Google Scholar] [CrossRef]
- Corsi, K.A.; Pollett, J.B.; Phillippi, J.A.; Usas, A.; Li, G.; Huard, J. Osteogenic potential of postnatal skeletal muscle-derived stem cells is influenced by donor sex. J. Bone Miner. Res. 2007, 22, 1592–1602. [Google Scholar] [CrossRef]
- Kretlow, J.D.; Jin, Y.Q.; Liu, W.; Zhang, W.J.; Hong, T.H.; Zhou, G.; Baggett, L.S.; Mikos, A.G.; Cao, Y. Donor age and cell passage affects differentiation potential of murine bone marrow-derived stem cells. BMC Cell Biol. 2008, 9, 60. [Google Scholar] [CrossRef]
- Rando, T.A. Stem cells, ageing and the quest for immortality. Nature 2006, 441, 1080–1086. [Google Scholar] [CrossRef]
- Zhang, H.; Menzies, K.J.; Auwerx, J. The role of mitochondria in stem cell fate and aging. Development 2018, 145, dev143420. [Google Scholar] [CrossRef]
- Ruzankina, Y.; Brown, E.J. Relationships between stem cell exhaustion, tumour suppression and ageing. Br. J. Cancer 2007, 97, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Xu, C.; Zhao, H.; Wang, J.; Feng, J.Q. A Biphasic Feature of Gli1(+)-Mesenchymal Progenitors during Cementogenesis That Is Positively Controlled by Wnt/β-Catenin Signaling. J. Dent. Res. 2021, 100, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.H.; Péault, B. A Gli(1)ttering Role for Perivascular Stem Cells in Blood Vessel Remodeling. Cell Stem Cell 2016, 19, 563–565. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.A.; Armstrong, R.C. Postnatal Sonic hedgehog (Shh) responsive cells give rise to oligodendrocyte lineage cells during myelination and in adulthood contribute to remyelination. Exp. Neurol. 2018, 299 Pt A, 122–136. [Google Scholar] [CrossRef]
- Zhang, Z. Bone regeneration by stem cell and tissue engineering in oral and maxillofacial region. Front. Med. 2011, 5, 401–413. [Google Scholar] [CrossRef]
- Bulgin, D.; Hodzic, E.; Komljenovic-Blitva, D. Advanced and prospective technologies for potential use in craniofacial tissues regeneration by stem cells and growth factors. J. Craniofac. Surg. 2011, 22, 342–348. [Google Scholar] [CrossRef]
- Behr, B.; Tang, C.; Germann, G.; Longaker, M.T.; Quarto, N. Locally applied vascular endothelial growth factor A increases the osteogenic healing capacity of human adipose-derived stem cells by promoting osteogenic and endothelial differentiation. Stem Cells 2011, 29, 286–296. [Google Scholar] [CrossRef]
- Andreas, K.; Sittinger, M.; Ringe, J. Toward in situ tissue engineering: Chemokine-guided stem cell recruitment. Trends Biotechnol. 2014, 32, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, X.T.; Yin, Y.; Wu, R.X.; Tian, B.M.; Chen, F.M. Administration of signalling molecules dictates stem cell homing for in situ regeneration. J. Cell. Mol. Med. 2017, 21, 3162–3177. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, C.; Farlie, P.; Penington, A.J. Bone regeneration in a rabbit critical-sized skull defect using autologous adipose-derived cells. Tissue Eng. Part A 2008, 14, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.X.; Yin, Y.; He, X.T.; Li, X.; Chen, F.M. Engineering a Cell Home for Stem Cell Homing and Accommodation. Adv. Biosyst. 2017, 1, e1700004. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Li, X.; He, X.T.; Wu, R.X.; Sun, H.H.; Chen, F.M. Leveraging Stem Cell Homing for Therapeutic Regeneration. J. Dent. Res. 2017, 96, 601–609. [Google Scholar] [CrossRef]
- Heng, B.C.; Cao, T.; Stanton, L.W.; Robson, P.; Olsen, B. Strategies for directing the differentiation of stem cells into the osteogenic lineage in vitro. J. Bone Miner. Res. 2004, 19, 1379–1394. [Google Scholar] [CrossRef]
- Sun, H.; Wu, C.; Dai, K.; Chang, J.; Tang, T. Proliferation and osteoblastic differentiation of human bone marrow-derived stromal cells on akermanite-bioactive ceramics. Biomaterials 2006, 27, 5651–5657. [Google Scholar] [CrossRef]
- Binulal, N.S.; Deepthy, M.; Selvamurugan, N.; Shalumon, K.T.; Suja, S.; Mony, U.; Jayakumar, R.; Nair, S.V. Role of nanofibrous poly(caprolactone) scaffolds in human mesenchymal stem cell attachment and spreading for in vitro bone tissue engineering—Response to osteogenic regulators. Tissue Eng. Part A 2010, 16, 393–404. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Osteogenic Patterns | Anatomical Position | Transgenic Mice Model | Intervention | Related Signaling | Results | References |
|---|---|---|---|---|---|---|
| Intramembranous ossification | Skull | Gli1-LacZ Gli1-CreERT2; tdTomato | Suture and calvarial bone injury | / | 1. Gli1+ MSCs are the cellular sources for injury repair and bone regeneration 2. The healing rate of calvarial bone is inversely proportional to the distance between the suture and injury site | [35] |
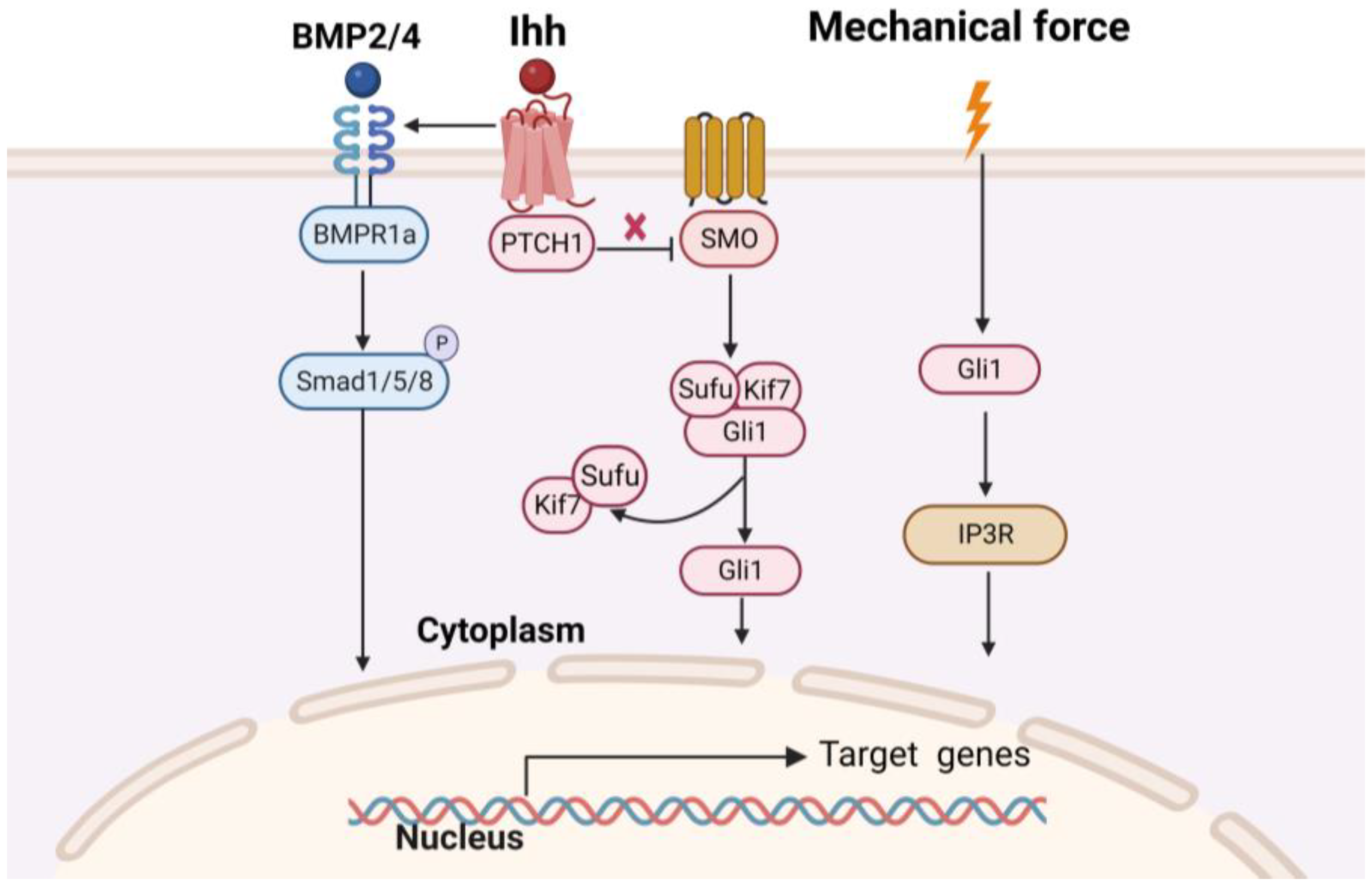
| Gli1-LacZG li1-CreERT2; mT/mG | Midpalatal suture expansion | IP3R | Gli1+ MSCs participated in mechanical force-induced osteogenesis by regulating IP3R-mediated intracellular calcium concentration | [36] | ||
| Gli1-LacZ Gli1-CreERT2; Ai14 Gli1-CreERT2; Ctnnb1f/f; Ai14 | Calvarial suture expansion | Wnt | 1. Gli1+ MSCs actively contributes to bone remodeling in response to tensile force 2. Conditional knockout of Ctnnb1 impeded the activation of Gli1+ MSCs, subsequently inhibiting the bone restoration under mechanical expansion | [37] | ||
| Gli1-LacZ Gli1-CreERT2; tdTomato Gli1-CreERT2; DTAf/f Gli1-CreERT2; Smoothenedf/f | Suture and calvarial bone injury | / | 1. Gli1+ MSCs in the suture mesenchyme give rise to osteogenic fronts, periosteum, and dura 2. Ablation of Gli1+ MSCs leads to craniosynostosis, skull growth arrest, and compromised injury repair. 3. Craniofacial bones exhibited severe osteoporosis and reduced bone volume following the blockage of the Hh pathway | [16] | ||
| Gli1-LacZ Gli1-CreERT2; tdTomato Gli1-CreERT2; Bmpr1af/f | Calvarial bone defect; addition of exogenous IHH | IHH/BMP | 1. Gli1+ MSCs give rise to osteoprogenitors that display active BMP signaling activity within the cranial suture 2. Loss of BMPR1a in Gli1+ MSCs disrupts osteoclastogenic activity by decreasing RANKL/OPG ratio and IHH activity 3. Upregulation of Hh signaling helped maintain the balance between osteoclastogenesis and osteogenesis in cranial sutures and partially restored the calvarial bone-healing process in BMPR1a mutant mice | [38] | ||
| Twist1+/− mice Gli1-CreERT2; Twist1f/f | Craniosynostosis model; Calvarial bone defect | / | 1. The regenerated suture creates a niche into which endogenous Gli1+ MSCs migrated, sustaining calvarial bone homeostasis and repair. 2. Using a biodegradable material combined with Gli1+ MSCs can successfully regenerate a functional cranial suture in Twist1+/− mice that corrects skull deformity, normalizes intracranial pressure, and rescues neurocognitive behavior deficits. | [39] | ||
| Alveolar bone | Gli1-CreERT2; TGFβR2f/f; tdTomato | / | TGF-β | Disrupting TGF-β signaling in Gli1+ MSCs leads to a reduction in OSX+ alveolar bone cell numbers, disturbance of periodontal homeostasis, and early postnatal alveolar bone loss | [40] | |
| Gli1-LacZ Gli1-CreERT2; Ai14 Gli1-CreERT2; Ctnnb1f/f; Ai14 | Physiological occlusal force | Wnt | 1. Gli1+ MSCs as multipotential stem cells contribute to the periodontium tissue turnover by migrating out of the NVB niche 2. Knockout of β-catenin results in a significant reduction of alveolar bone height and density, and over half of the molar root surface was exposed 3. Extracting one side molar arrested Gli1+ MSCs activation in opposing molars, resulting in PDL tissue loss and reduced Wnt activity. | [41] | ||
| Gli1-CreERT2; tdTomato | Orthodontic force | / | Gli1+ MSCs in the PDL, as a source of osteoblasts on the tension side of the alveolar bone, can proliferate and differentiate into osteoblasts and fibroblasts during orthodontic tooth movement | [42] | ||
| Gli1-CreERT2; Ai14 Gli1CreERT2; Ai14; eGFP-DTAGli1-creERT2; β-cateninf/f; Ai14 | Tooth extraction; Implant placement | Wnt | 1. Gli1+ MSCs were activated and proliferated along blood vessels after tooth extraction, and their progeny contributed to new bone formation 2. The bone volume and density reduction surrounding the implant in the β-catenin ablation group compromised the healing and osseointegration processes. | [33] | ||
| Gli1-CreERT2; tdTomato | Tooth transplantation | / | Gli1+ MSCs are localized within the mature PDL exhibited stem cell properties and could differentiate into osteoblasts and osteocytes during alveolar bone regeneration. | [43] | ||
| Gli1-LacZ Gli1-CreERT2; eGFP-DTA Gli1-CreERT2; YAPf/f | Orthodontic force | YAP/TAZ | 1. MSCs expressing Gli1 can respond to orthodontic force by supplying Runx2+ cells for alveolar bone remodeling 2. Conditional ablation of the Yap gene in Gli1+ MSCs can suppress osteogenic differentiation and defective bone formation | [44] | ||
| Endochondral ossification | Mandibular condyle | Gli1-CreERT2; tdTomato | Condyle fracture and Sham surgery | / | 1. Gli1+ MSCs are spatially located at the superficial layers of the cartilage and chondro-osseous junction, and contribute to osteoblasts in the subchondral bone during condyle postnatal development. 2. Gli1+ MSCs could differentiate into osteoblasts and chondrocytes during condylar fracture healing. 3. Wnt/β-catenin signaling mediates the proliferation and osteogenic differentiation of Gli1+ MSCs in vitro. | [45] |
| Gli1-CreERT2; tdTomato Gli1-CreERT2;Smofl/fl | TMJOA | Hh | 1. Gli1+ MSCs are osteogenic progenitors contributing to subchondral bone formation and homeostasis in the mandibular condyle of the temporomandibular joint in vivo. 2. Uneven distribution of osteogenic differentiation of Gli1+ MSCs in the subchondral bone leads to abnormal subchondral bone remodeling via Hh signaling activation and to the development of TMJOA. 3. The selective pharmacological inhibition and specific genetic inhibition of Hh signaling in Gli1+ MSCs results in improved subchondral bone microstructure, attenuated local immune inflammatory response in the subchondral bone, and reduced degeneration of the articular cartilage. | [46] | ||
| Bone vasculature formation. | Vessel | Gli1-LacZ Gli1- creERT2;iDTA | / | / | 1. Type H vessels are identified with distinct functional properties to couple angiogenesis to osteogenesis and mediate bone generation. 2. Type H vessels are the preferable vascular subtype in which Gli1+ MSCs are adjacently localized, and mediate developmental and regenerative angiogenesis in bone. 3. Gli1+ MSC ablation inhibits type H vessel formation associated with suppressed bone generation and regeneration. | [47] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.; Liu, Z.; Xiao, L.; Ai, M.; Cao, Y.; Mao, J.; Song, K. The Role of Gli1+ Mesenchymal Stem Cells in Osteogenesis of Craniofacial Bone. Biomolecules 2023, 13, 1351. https://doi.org/10.3390/biom13091351
Wu L, Liu Z, Xiao L, Ai M, Cao Y, Mao J, Song K. The Role of Gli1+ Mesenchymal Stem Cells in Osteogenesis of Craniofacial Bone. Biomolecules. 2023; 13(9):1351. https://doi.org/10.3390/biom13091351
Chicago/Turabian StyleWu, Laidi, Zhixin Liu, Li Xiao, Mi Ai, Yingguang Cao, Jing Mao, and Ke Song. 2023. "The Role of Gli1+ Mesenchymal Stem Cells in Osteogenesis of Craniofacial Bone" Biomolecules 13, no. 9: 1351. https://doi.org/10.3390/biom13091351