
Assigning the Absolute Configuration of Inositol Poly- and Pyrophosphates by NMR Using a Single Chiral Solvating Agent

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. NMR Measurements
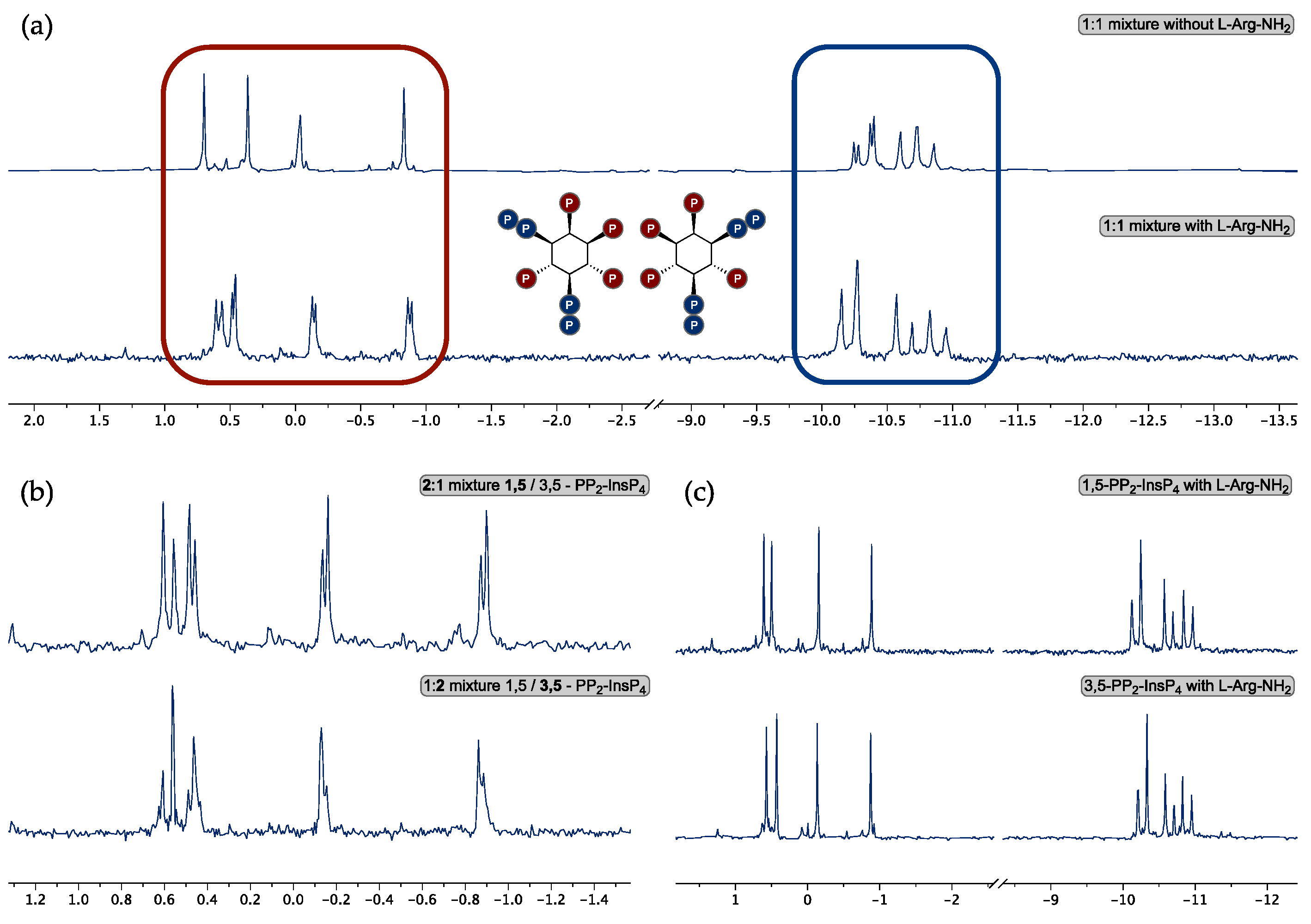
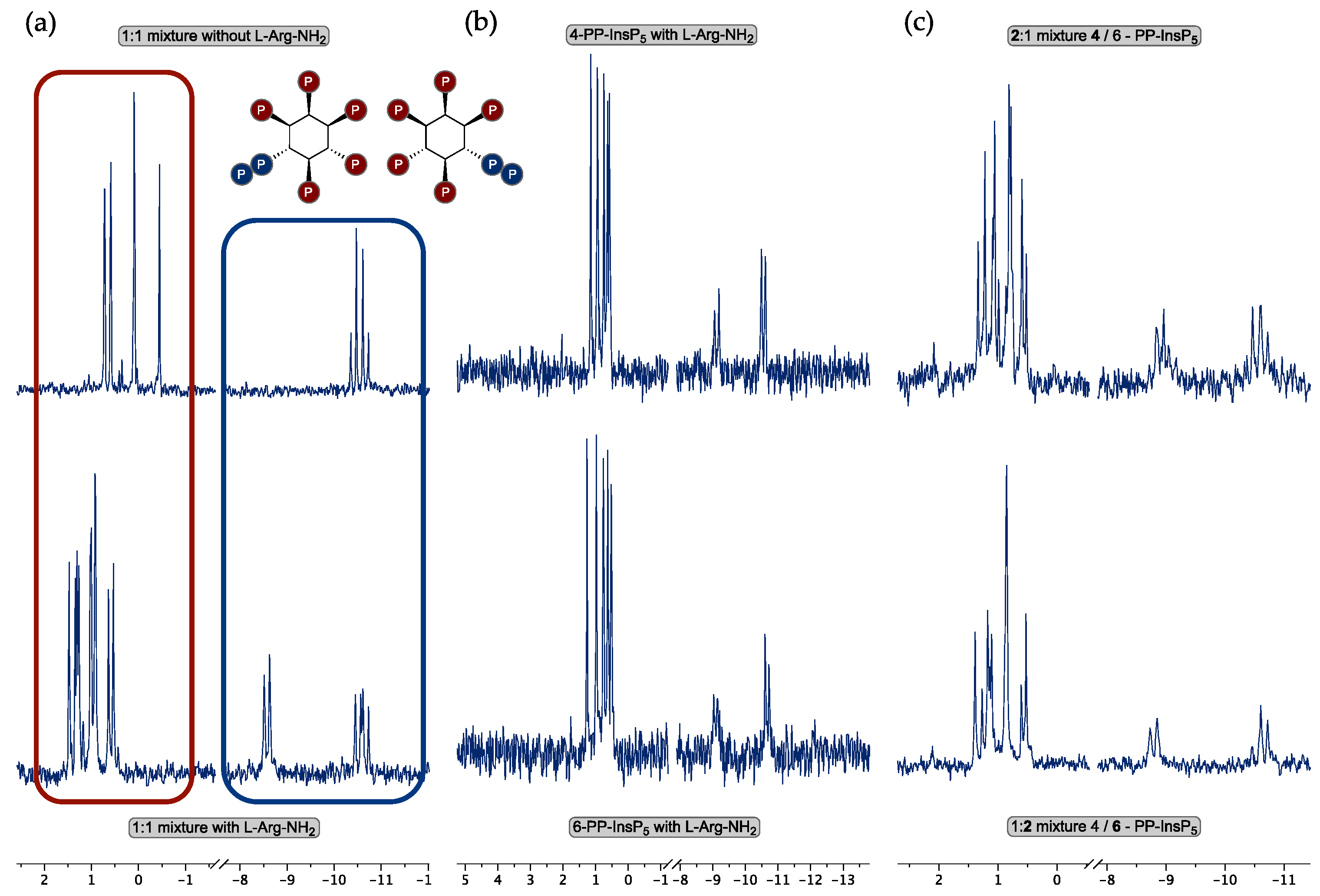
3. Results
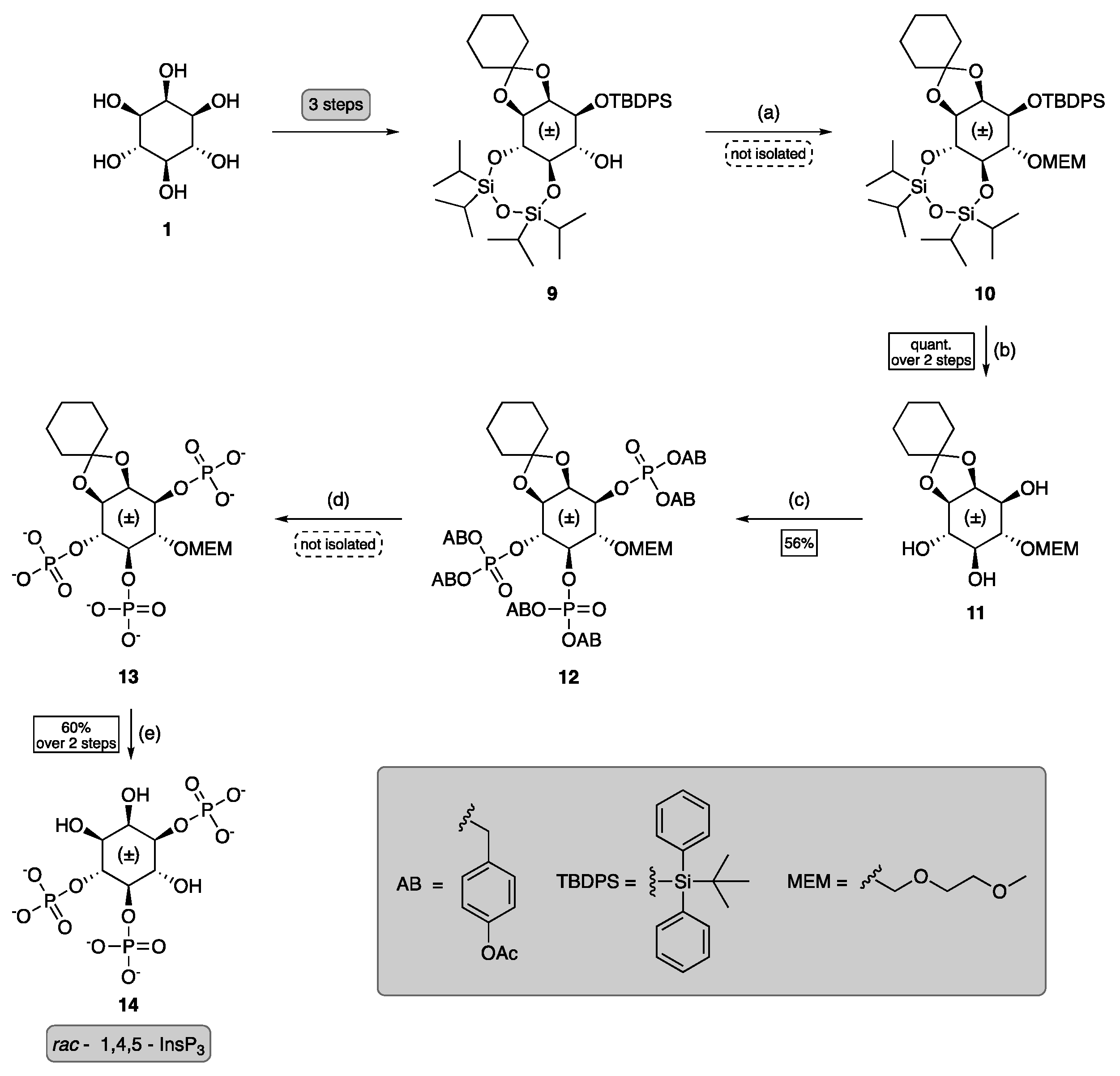
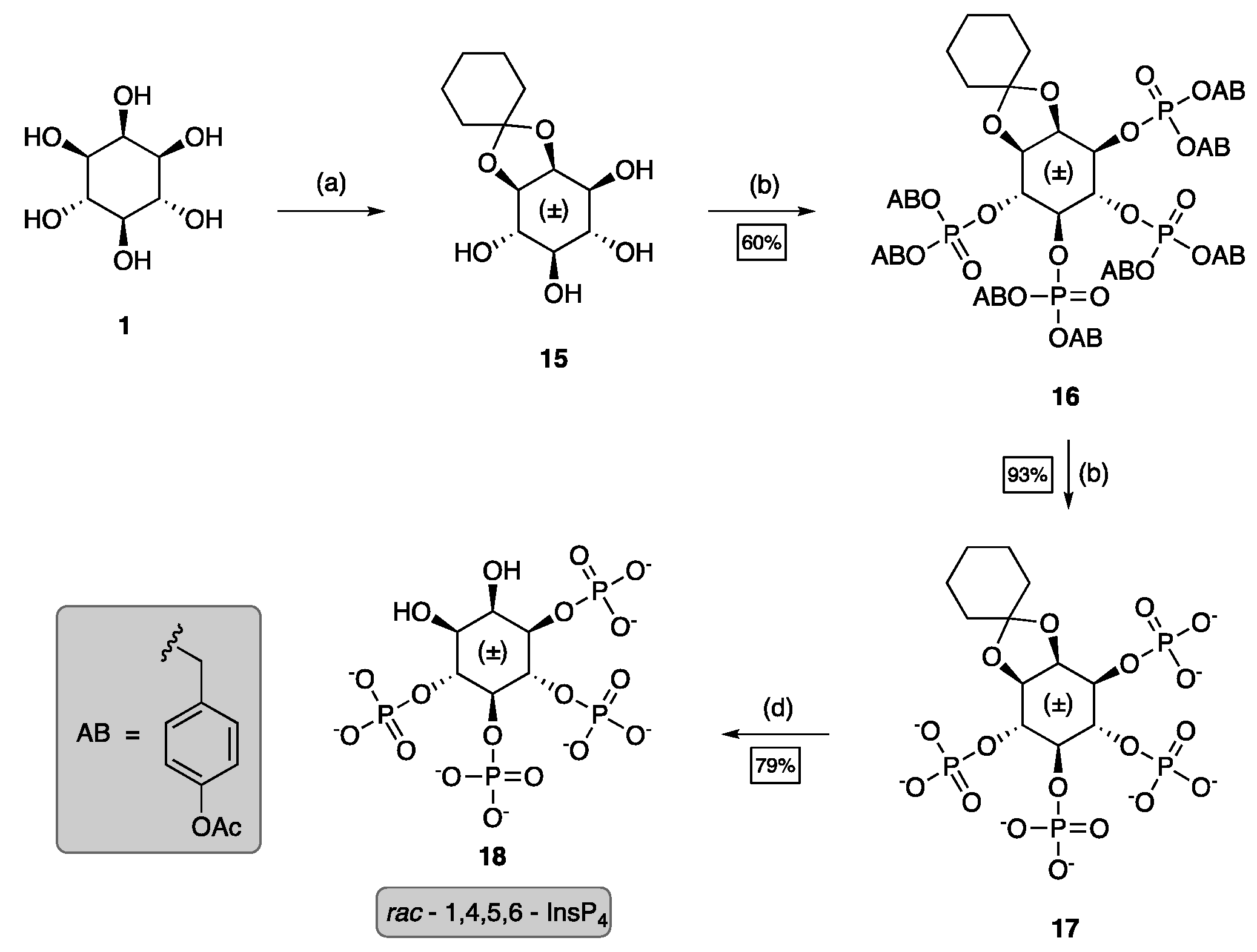
3.1. Chemical Synthesis of InsP Model Compounds
3.2. Enantiomer Assignment by 31P-NMR Using a Chiral Solvating Agent
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bittner, T.; Wittwer, C.; Hauke, S.; Wohlwend, D.; Mundinger, S.; Dutta, A.K.; Bezold, D.; Dürr, T.; Friedrich, T.; Schultz, C.; et al. Photolysis of Caged Inositol Pyrophosphate InsP8 Directly Modulates Intracellular Ca2+Oscillations and Controls C2AB Domain Localization. J. Am. Chem. Soc. 2020, 142, 10606–10611. [Google Scholar] [CrossRef] [PubMed]
- Thota, S.G.; Bhandari, R. The Emerging Roles of Inositol Pyrophosphates in Eukaryotic Cell Physiology. J. Biosci. 2015, 40, 593–605. [Google Scholar] [CrossRef]
- Chakraborty, A.; Koldobskiy, M.A.; Bello, N.T.; Maxwell, M.; Potter, J.J.; Juluri, K.R.; Maag, D.; Kim, S.; Huang, A.S.; Dailey, M.J.; et al. Inositol Pyrophosphates Inhibit Akt Signaling, Thereby Regulating Insulin Sensitivity and Weight Gain. Cell 2010, 143, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Illies, C.; Gromada, J.; Fiume, R.; Leibiger, B.; Yu, J.; Juhl, K.; Yang, S.-N.; Barma, D.K.; Falck, J.R.; Saiardi, A.; et al. Requirement of Inositol Pyrophosphates for Full Exocytotic Capacity in Pancreatic β Cells. Science 2007, 318, 1299–1302. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-S.; Lee, J.-Y.; Kyung, J.W.; Yang, Y.; Park, S.J.; Lee, S.; Pavlovic, I.; Kong, B.; Jho, Y.S.; Jessen, H.J.; et al. Inositol Pyrophosphates Inhibit Synaptotagmin-Dependent Exocytosis. Proc. Natl. Acad. Sci. USA 2016, 113, 8314–8319. [Google Scholar] [CrossRef] [PubMed]
- Hauke, S.; Dutta, A.K.; Eisenbeis, V.B.; Bezold, D.; Bittner, T.; Wittwer, C.; Thakor, D.; Pavlovic, I.; Schultz, C.; Jessen, H.J. Photolysis of Cell-Permeant Caged Inositol Pyrophosphates Controls Oscillations of Cytosolic Calcium in a β-Cell Line. Chem. Sci. 2019, 10, 2687–2692. [Google Scholar] [CrossRef] [PubMed]
- Szijgyarto, Z.; Garedew, A.; Azevedo, C.; Saiardi, A. Influence of Inositol Pyrophosphates on Cellular Energy Dynamics. Science 2011, 334, 802–805. [Google Scholar] [CrossRef]
- Irvine, R.F.; Schell, M.J. Back in the Water: The Return of the Inositol Phosphates. Nat. Rev. Mol. Cell. Biol. 2001, 2, 327–338. [Google Scholar] [CrossRef]
- Qiu, D.; Gu, C.; Liu, G.; Ritter, K.; Eisenbeis, V.B.; Bittner, T.; Gruzdev, A.; Seidel, L.; Bengsch, B.; Shears, S.B.; et al. Capillary Electrophoresis Mass Spectrometry Identifies New Isomers of Inositol Pyrophosphates in Mammalian Tissues. Chem. Sci. 2022, 14, 658–667. [Google Scholar] [CrossRef]
- Riemer, E.; Qiu, D.; Laha, D.; Harmel, R.K.; Gaugler, P.; Gaugler, V.; Frei, M.; Hajirezaei, M.R.; Laha, N.P.; Krusenbaum, L.; et al. ITPK1 Is an InsP6/ADP Phosphotransferase That Controls Phosphate Signaling in Arabidopsis. Mol. Plant 2021, 14, 1864–1880. [Google Scholar] [CrossRef]
- Wang, H.; Nair, V.S.; Holland, A.A.; Capolicchio, S.; Jessen, H.J.; Johnson, M.K.; Shears, S.B. Asp1 from Schizosaccharomyces Pombe Binds a [2Fe-2S]2+ Cluster Which Inhibits Inositol Pyrophosphate 1-Phosphatase Activity. Biochemistry 2015, 54, 6462–6474. [Google Scholar] [CrossRef]
- Mulugu, S.; Bai, W.; Fridy, P.C.; Bastidas, R.J.; Otto, J.C.; Dollins, D.E.; Haystead, T.A.; Ribeiro, A.A.; York, J.D. A Conserved Family of Enzymes That Phosphorylate Inositol Hexakisphosphate. Science 2007, 316, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Laussmann, T.; Reddy, K.M.; Reddy, K.K.; Falck, J.R.; Vogel, G. Diphospho-Myo-Inositol Phosphates from Dictyostelium Identified as d-6-Diphospho-Myo-Inositol Pentakisphosphate and d-5,6-Bisdiphospho-Myo-Inositol Tetrakisphosphate. Biochem. J. 1997, 322, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Falck, J.R.; Hall, T.M.T.; Shears, S.B. Structural Basis for an Inositol Pyrophosphate Kinase Surmounting Phosphate Crowding. Nat. Chem. Biol. 2012, 8, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Fridy, P.C.; Ribeiro, A.A.; Choi, J.H.; Barma, D.K.; Vogel, G.; Falck, J.R.; Shears, S.B.; York, J.D.; Mayr, G.W. Structural Analysis and Detection of Biological Inositol Pyrophosphates Reveal That the Family of VIP/Diphosphoinositol Pentakisphosphate Kinases Are1/3-Kinases. J. Biol. Chem. 2009, 284, 1863–1872. [Google Scholar] [CrossRef]
- Losito, O.; Szijgyarto, Z.; Resnick, A.C.; Saiardi, A. Inositol Pyrophosphates and Their Unique Metabolic Complexity: Analysis by Gel Electrophoresis. PLoS ONE 2009, 4, e5580. [Google Scholar] [CrossRef]
- Qiu, D.; Wilson, M.S.; Eisenbeis, V.B.; Harmel, R.K.; Riemer, E.; Haas, T.M.; Wittwer, C.; Jork, N.; Gu, C.; Shears, S.B.; et al. Analysis of Inositol Phosphate Metabolism by Capillary Electrophoresis Electrospray Ionization Mass Spectrometry. Nat. Commun. 2020, 11, 6035. [Google Scholar] [CrossRef]
- Laussmann, T.; Eujen, R.; Weisshuhn, C.M.; Thiel, U.; Vogel, G. Structures of Diphospho-Myo-Inositol Pentakisphosphate and Bisdiphospho-Myo-Inositol Tetrakisphosphate from Dictyostelium Resolved by NMR Analysis. Biochem. J. 1996, 315, 715–720. [Google Scholar] [CrossRef]
- Laha, D.; Parvin, N.; Hofer, A.; Giehl, R.F.H.; Fernandez-Rebollo, N.; Von Wirén, N.; Saiardi, A.; Jessen, H.J.; Schaaf, G. Arabidopsis ITPK1 and ITPK2 Have an Evolutionarily Conserved Phytic Acid Kinase Activity. ACS Chem. Biol. 2019, 14, 2127–2133. [Google Scholar] [CrossRef]
- Draškovič, P.; Saiardi, A.; Bhandari, R.; Burton, A.; Ilc, G.; Kovačevič, M.; Snyder, S.H.; Podobnik, M. Inositol Hexakisphosphate Kinase Products Contain Diphosphate and Triphosphate Groups. Chem. Biol. 2008, 15, 274–286. [Google Scholar] [CrossRef]
- Capolicchio, S.; Wang, H.; Thakor, D.T.; Shears, S.B.; Jessen, H.J. Synthesis of Densely Phosphorylated Bis-1,5-Diphospho-Myo-Inositol Tetrakisphosphate and Its Enantiomer by Bidirectional P-Anhydride Formation. Angew. Chem. Int. Ed. 2014, 53, 9508–9511. [Google Scholar] [CrossRef] [PubMed]
- Capolicchio, S.; Thakor, D.T.; Linden, A.; Jessen, H.J. Synthesis of Unsymmetric Diphospho-Inositol Polyphosphates. Angew. Chem. Int. Ed. 2013, 52, 6912–6916. [Google Scholar] [CrossRef]
- Nguyen Trung, M.; Kieninger, S.; Fandi, Z.; Qiu, D.; Liu, G.; Mehendale, N.K.; Saiardi, A.; Jessen, H.; Keller, B.; Fiedler, D. Stable Isotopomers of Myo-Inositol Uncover a Complex MINPP1-Dependent Inositol Phosphate Network. ACS Cent. Sci. 2022, 8, 1683–1694. [Google Scholar] [CrossRef] [PubMed]
- Blüher, D.; Laha, D.; Thieme, S.; Hofer, A.; Eschen-Lippold, L.; Masch, A.; Balcke, G.; Pavlovic, I.; Nagel, O.; Schonsky, A.; et al. A 1-Phytase Type III Effector Interferes with Plant Hormone Signaling. Nat. Commun. 2017, 8, 2159. [Google Scholar] [CrossRef]
- Wu, M.; Chong, L.S.; Capolicchio, S.; Jessen, H.J.; Resnick, A.C.; Fiedler, D. Elucidating Diphosphoinositol Polyphosphate Function with Nonhydrolyzable Analogues. Angew. Chem. Int. Ed. 2014, 53, 7192–7197. [Google Scholar] [CrossRef]
- Hostachy, S.; Utesch, T.; Franke, K.; Dornan, G.L.; Furkert, D.; Türkaydin, B.; Haucke, V.; Sun, H.; Fiedler, D. Dissecting the Activation of Insulin Degrading Enzyme by Inositol Pyrophosphates and Their Bisphosphonate Analogs. Chem. Sci. 2021, 12, 10696–10702. [Google Scholar] [CrossRef]
- Haas, T.M.; Mundinger, S.; Qiu, D.; Jork, N.; Ritter, K.; Dürr-Mayer, T.; Ripp, A.; Saiardi, A.; Schaaf, G.; Jessen, H.J. Stable Isotope Phosphate Labelling of Diverse Metabolites Is Enabled by a Family of 18O-Phosphoramidites**. Angew. Chem. Int. Ed. 2022, 61, e202112457. [Google Scholar] [CrossRef]
- Kiely, D.E.; Abruscato, G.J.; Baburao, V. A Synthesis of (±)Myo-Inositol 1-Phosphate. Carbohydr. Res. 1974, 34, 307–313. [Google Scholar] [CrossRef]
- Rukavishnikov, A.V.; Ryan, M.; Griffith, O.H.; Keana, J.F.W. A Chromogenic Substrate for the Continuous Assay of Mammalian Phosphoinositide-Specific Phospholipase C. Bioorg. Med. Chem. Lett. 1997, 7, 1239–1242. [Google Scholar] [CrossRef]
- Subramanian, D.; Laketa, V.; Müller, R.; Tischer, C.; Zarbakhsh, S.; Pepperkok, R.; Schultz, C. Activation of Membrane-Permeant Caged PtdIns(3)P Induces Endosomal Fusion in Cells. Nat. Chem. Biol. 2010, 6, 324–326. [Google Scholar] [CrossRef]
- Jessen, H.J.; Schulz, T.; Balzarini, J.; Meier, C. Bioreversible Protection of Nucleoside Diphosphates. Angew. Chem. Int. Ed. 2008, 47, 8719–8722. [Google Scholar] [CrossRef] [PubMed]
- Furkert, D.; Hostachy, S.; Nadler-Holly, M.; Fiedler, D. Triplexed Affinity Reagents to Sample the Mammalian Inositol Pyrophosphate Interactome. Cell Chem. Biol. 2020, 27, 1097–1108.e4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Thompson, J.; Prestwich, G.D. A Scalable Synthesis of the IP7 Isomer, 5-PP-Ins(1,2,3,4,6)P5. Org. Lett. 2009, 11, 1551–1554. [Google Scholar] [CrossRef]
- Desfougères, Y.; Wilson, M.S.C.; Laha, D.; Miller, G.J.; Saiardi, A. ITPK1 Mediates the Lipid-Independent Synthesis of Inositol Phosphates Controlled by Metabolism. Proc. Natl. Acad. Sci. USA 2019, 116, 24551–24561. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Parekh, N.; Mondal, B.; Gupta, S.S. Controlled Synthesis of End-Functionalized Mannose-6-Phosphate Glycopolypeptides for Lysosome Targeting. ACS Macro. Lett. 2016, 5, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Kurz, L.; Schmieder, P.; Veiga, N.; Fiedler, D. One Scaffold, Two Conformations: The Ring-Flip of the Messenger InsP8 Occurs under Cytosolic Conditions. Biomolecules 2023, 13, 645. [Google Scholar] [CrossRef]
- Liu, G.; Riemer, E.; Schneider, R.; Cabuzu, D.; Bonny, O.; Wagner, C.A.; Qiu, D.; Saiardi, A.; Strauss, A.; Lahaye, T.; et al. The Phytase RipBL1 Enables the Assignment of a Specific Inositol Phosphate Isomer as a Structural Component of Human Kidney Stones. RSC Chem. Biol. 2023, 4, 300–309. [Google Scholar] [CrossRef]
- Qiu, D.; Eisenbeis, V.B.; Saiardi, A.; Jessen, H.J. Absolute Quantitation of Inositol Pyrophosphates by Capillary Electrophoresis Electrospray Ionization Mass Spectrometry. J. Vis. Exp. 2021, 174, e62847. [Google Scholar] [CrossRef]
- Wilson, M.; Saiardi, A. Inositol Phosphates Purification Using Titanium Dioxide Beads. Bio Protoc. 2018, 8, e2959. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ritter, K.; Jork, N.; Unmüßig, A.-S.; Köhn, M.; Jessen, H.J. Assigning the Absolute Configuration of Inositol Poly- and Pyrophosphates by NMR Using a Single Chiral Solvating Agent. Biomolecules 2023, 13, 1150. https://doi.org/10.3390/biom13071150
Ritter K, Jork N, Unmüßig A-S, Köhn M, Jessen HJ. Assigning the Absolute Configuration of Inositol Poly- and Pyrophosphates by NMR Using a Single Chiral Solvating Agent. Biomolecules. 2023; 13(7):1150. https://doi.org/10.3390/biom13071150
Chicago/Turabian StyleRitter, Kevin, Nikolaus Jork, Anne-Sophie Unmüßig, Maja Köhn, and Henning J. Jessen. 2023. "Assigning the Absolute Configuration of Inositol Poly- and Pyrophosphates by NMR Using a Single Chiral Solvating Agent" Biomolecules 13, no. 7: 1150. https://doi.org/10.3390/biom13071150