

From Environmental Epigenetics to the Inheritance of Acquired Traits: A Historian and Molecular Perspective on an Unnecessary Lamarckian Explanation

{kind=link}
{kind=link}
Abstract
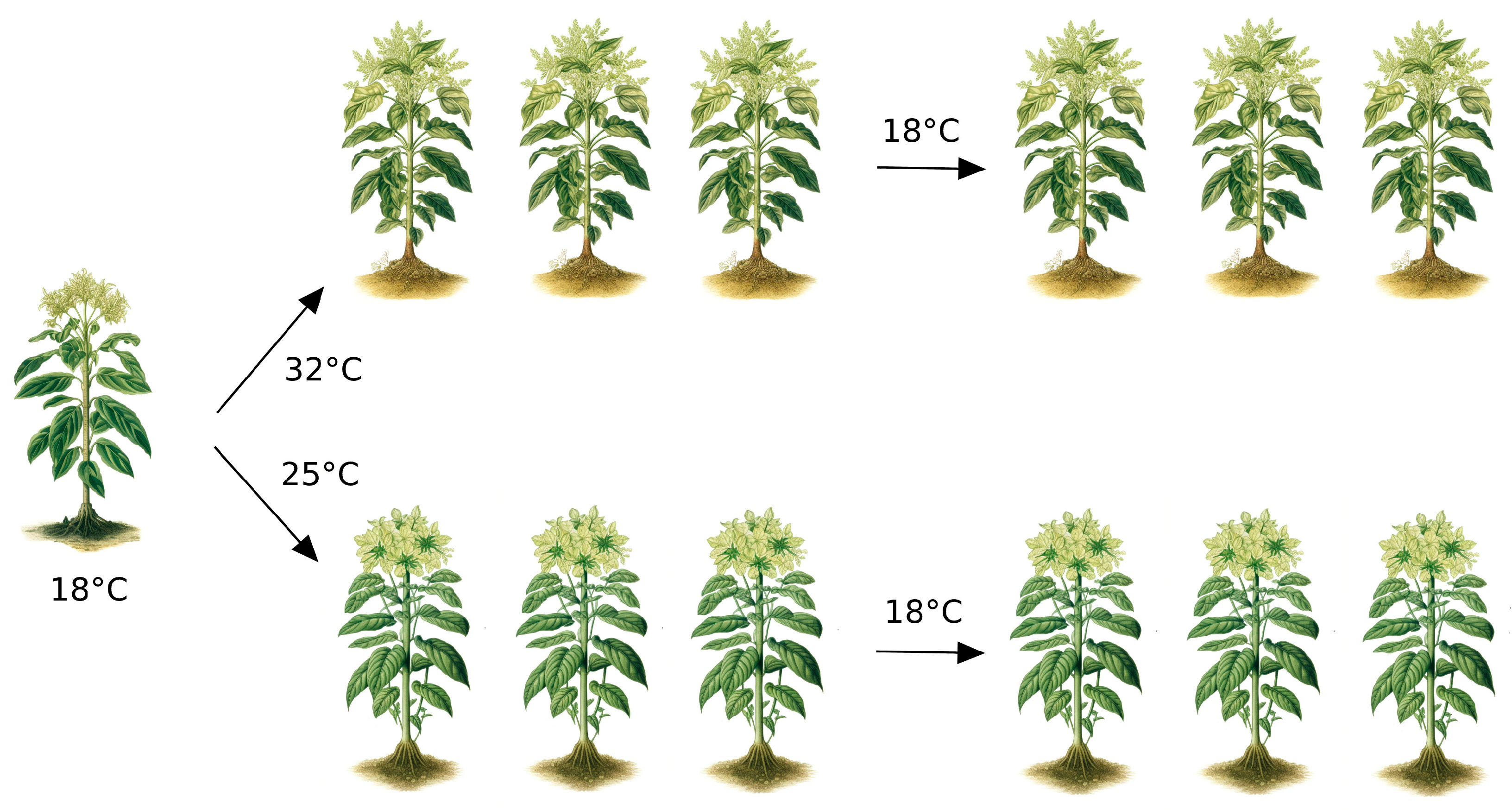
:1. Introduction
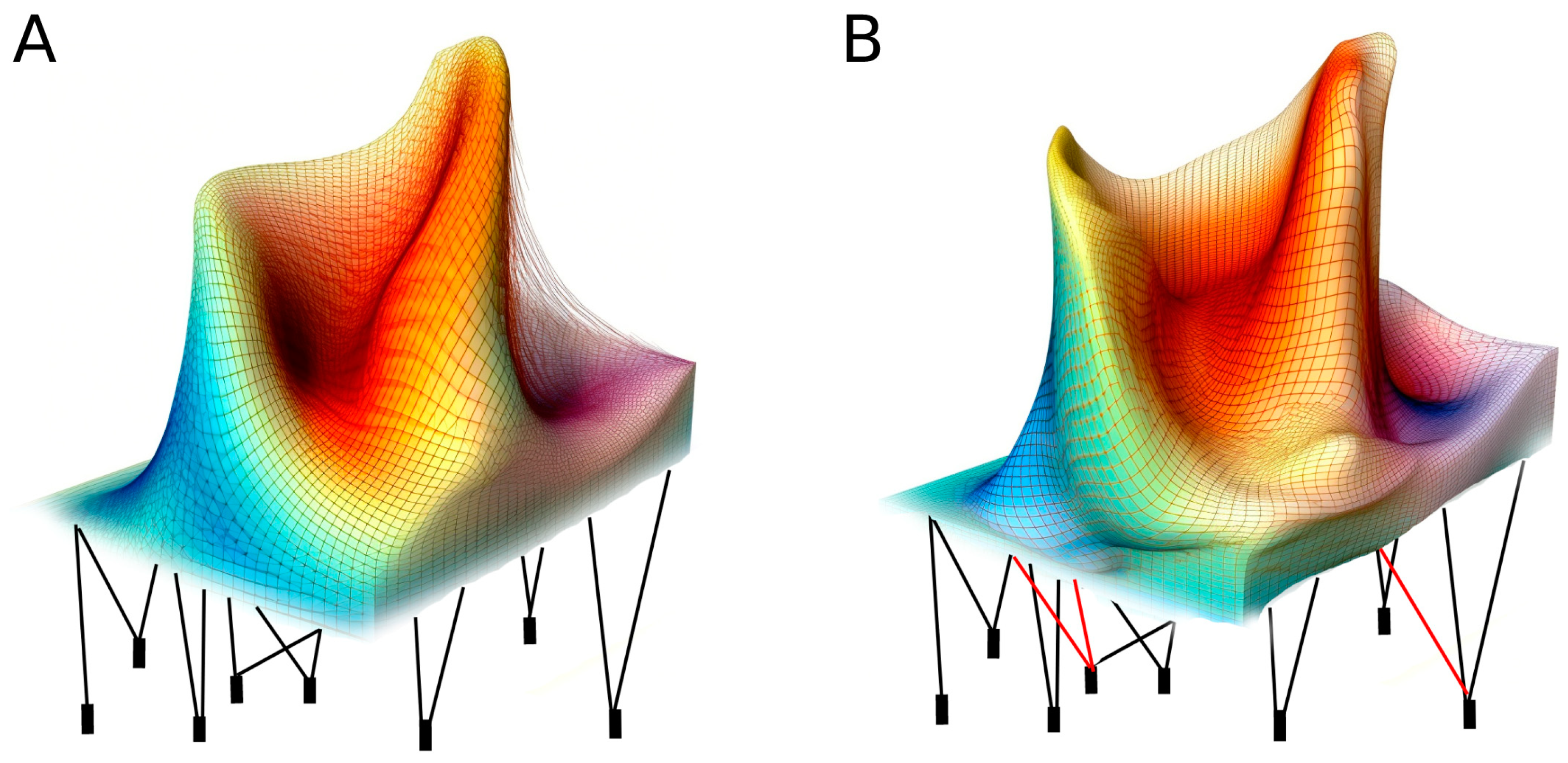
2. A Discussion about Terms
3. Toward a New Neo-Lamarckism?
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waddington, C.H. Organisers and Genes; Cambridge University Press: Cambridge, UK, 1940. [Google Scholar]
- Dupont, C.; Armant, D.R.; Brenner, C.A. Epigenetics: Definition, Mechanisms and Clinical Perspective. Semin. Reprod. Med. 2009, 27, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Kan, R.L.; Chen, J.; Sallam, T. Crosstalk between epitranscriptomic and epigenetic mechanisms in gene regulation. Trends Genet. 2021, 38, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Deans, C.; Maggert, K.A. What do you mean, “epigenetic”? Genetics 2015, 199, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandrioli, M.; Borsatti, F. Analysis of heterochromatic epigenetic markers in the holocentric chromosomes of the aphid Acyrthosiphon pisum. Chrom. Res. 2007, 15, 1015–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandrioli, M. A new synthesis in epigenetics: Towards a unified function of DNA methylation from invertebrates to vertebrates. Cell. Mol. Life Sci. 2007, 64, 2522–2524. [Google Scholar] [CrossRef] [Green Version]
- Tammen, S.A.; Friso, S.; Choi, S.W. Epigenetics: The link between nature and nurture. Mol. Asp. Med. 2013, 34, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Bannister, A.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Z.; Jia, J.; Du, T.; Zhang, N.; Tang, Y.; Fang, Y.; Fang, D. Overview of histone modification. Adv. Exp. Med. Biol. 2021, 1283, 1–16. [Google Scholar]
- Wei, J.W.; Huang, K.; Yang, C.; Kang, C.S. Non-coding RNAs as regulators in epigenetics. Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Kilvitis, H.J.; Alvarez, M.; Foust, C.M.; Schrey, A.W.; Robertson, M.; Richards, C.L. Ecological epigenetics. Adv. Exp. Med. Biol. 2014, 781, 191–210. [Google Scholar]
- Guillette, L.J.; Parrott, B.B.; Nilsson, E.; Haque, M.M.; Skinner, M.K. Epigenetic programming alterations in alligators from environmentally contaminated lakes. Gen. Comp. Endocrinol. 2016, 238, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrizio, P.; Garvis, S.; Palladino, F. Histone methylation and memory of environmental stress. Cells 2019, 8, 339. [Google Scholar] [CrossRef] [Green Version]
- Weaver, I.C.G.; Champagne, F.A.; Brown, S.E.; Dymov, S.; Sharma, S.; Meaney, M.J.; Szyf, M. Reversal of maternal programming of stress responses in adult offspring through methyl supplementation: Altering epigenetic marking later in life. J. Neurosci. 2005, 25, 11045–11054. [Google Scholar] [CrossRef] [Green Version]
- Ledón-Rettig, C.C.; Richards, C.L.; Martin, L.B. Epigenetics for behavioral ecologists. Behav. Ecol. 2012, 24, 311–324. [Google Scholar] [CrossRef] [Green Version]
- Ashe, A.; Colot, V.; Oldroyd, B.P. How does epigenetics influence the course of evolution? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, e20200111. [Google Scholar] [CrossRef] [PubMed]
- Endler, J.A. Natural Selection in the Wild; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Herrera, C.M.; Bazaga, P. Epigenetic differentiation and relationship to adaptive genetic divergence in discrete populations of the violet Viola cazorlensis. New Phytol. 2010, 187, 867–876. [Google Scholar] [CrossRef]
- Lamarck, J.B. Discours d’ouverture des Cours de Zoologie donnés dans le Museum d’Histoire naturelle (1800–1806). Bull. Sci. Fr. Belg. 1907, 40, 453–595. [Google Scholar]
- Lamarck, J.B. Philosophie Zoologique, ou Exposition des Considérations Relatives à L’histoire Naturelle des Animaux; Librairie F. Savy: Paris, France, 1809. [Google Scholar]
- Hala, D.; Huggett, D.B.; Burggren, W.W. Environmental stressors and the epigenome. Drug Discov. Today Technol. 2014, 12, e3–e8. [Google Scholar] [CrossRef]
- Lillycrop, K.A.; Burdge, G.C. Environmental challenge, epigenetic plasticity and the induction of altered phenotypes in mammals. Epigenomics 2014, 6, 623–636. [Google Scholar] [CrossRef]
- Koonin, E.V. Calorie restriction à Lamarck. Cell 2014, 158, 237–238. [Google Scholar] [CrossRef] [Green Version]
- Joo, E.; Dowty, J.G.; Milne, R.L.; Wong, E.M.; Dugué, P.A.; English, D.; Hopper, J.L.; Goldgar, D.E.; Giles, G.G.; Southey, M.C. Heritable DNA methylation marks associated with susceptibility to breast cancer. Nat. Commun. 2018, 9, 867. [Google Scholar] [CrossRef] [Green Version]
- Bošković, A.; Rando, O.J. Transgenerational epigenetic inheritance. Annu. Rev. Genet. 2018, 52, 21–41. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Y.; Liu, T.; Chang, H.; Yang, X. The regulation mechanisms and the Lamarckian inheritance property of DNA methylation in animals. Mamm. Genome 2021, 32, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Stajic, D.; Jansen, L.E.T. Empirical evidence for epigenetic inheritance driving evolutionary adaptation. Phil. Trans. R. Soc. B 2021, 376, e20200121. [Google Scholar] [CrossRef] [PubMed]
- Harvey, Z.H.; Chen, Y.; Jarosz, D.F. Protein-based inheritance: Epigenetics beyond the chromosome. Mol. Cell 2018, 69, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Nicoglou, A.; Merlin, F. Epigenetics: A way to bridge the gap between biological fields. Stud. Hist. Philos. Biol. Biomed. Sci. 2017, 66, 73–82. [Google Scholar] [CrossRef]
- Jablonka, E.; Lamb, M.J. The inheritance of acquired epigenetic variations. J. Theor. Biol. 1989, 139, 69–83. [Google Scholar] [CrossRef]
- Jablonka, E.; Raz, G. Transgenerational epigenetic inheritance: Prevalence, mechanisms, and implications for the study of heredity and evolution. Q. Rev. Biol. 2009, 84, 131–176. [Google Scholar] [CrossRef] [Green Version]
- Pigliucci, M.; Müller, G.B. Elements of an extended evolutionary synthesis. In Evolution, the Extended Synthesis; Pigliucci, M., Müller, G.B., Eds.; MIT Press: Cambridge, MA, USA, 2010; pp. 3–17. [Google Scholar]
- Jablonka, E.; Lamb, M.J. Inheritance Systems and the Extended Evolutionary Synthesis; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Shapiro, J.A. What we have learned about evolutionary genome change in the past 7 decades. Biosystems 2022, 215–216, e104669. [Google Scholar] [CrossRef]
- Shapiro, J.A. Why the third way of evolution is necessary. Theor. Biol. Forum. 2021, 114, 13–26. [Google Scholar]
- Martin, C.; Zhang, Y. Mechanisms of epigenetic inheritance. Curr. Opin. Cell Biol. 2007, 19, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Youngston, N.A.; Whitelaw, E. Transgenerational epigenetic effects. Annu. Rev. Genom. Hum. Genet. 2008, 9, 233–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heard, E.; Martienssen, R.A. Transgenerational epigenetic inheritance: Myths and mechanisms. Cell 2014, 157, 95–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart-Morgan, K.R.; Petryk, N.; Groth, A. Chromatin replication and epigenetic cell memory. Nat. Cell Biol. 2020, 22, 361–371. [Google Scholar] [CrossRef]
- Skinner, M.K. What is an epigenetic transgenerational phenotype? F3 or F2. Reprod. Toxicol. 2008, 5, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, A.B.; Morgan, C.P.; Leu, N.A.; Bale, T.L. Transgenerational epigenetic programming via sperm microRNA recapitulates effects of paternal stress. Proc. Natl. Acad. Sci. USA 2015, 112, 13699–13704. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; Sun, Z. Lamarck rises from his grave: Parental environment-induced epigenetic inheritance in model organisms and humans. Biol. Rev. 2017, 92, 2084–2111. [Google Scholar] [CrossRef] [Green Version]
- Perez, M.F.; Lehner, B. Intergenerational and transgenerational epigenetic inheritance in animals. Nat. Cell Biol. 2019, 21, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Mounger, J.; Ainouche, M.L.; Bossdorf, O.; Cavé-Radet, A.; Li, B.; Parepa, M.; Salmon, A.; Yang, J.; Richards, C.L. Epigenetics and the success of invasive plants. Phil. Trans. R. Soc. B 2021, 376, e20200117. [Google Scholar] [CrossRef]
- McGuigan, K.; Hoffmann, A.A.; Sgrò, C.M. How is epigenetics predicted to contribute to climate change adaptation? What evidence do we need? Phil. Trans. R. Soc. B 2021, 376, e20200119. [Google Scholar] [CrossRef]
- Mandrioli, M.; Manicardi, G.C. Cytosine methylation in insects: New routes for the comprehension of insect complexity. AIMS Biophys. 2015, 2, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Mayr, E. Lamarck revisited. J. Hist. Biol. 1972, 5, 55–94. [Google Scholar] [CrossRef]
- Osborn, H.F. From the Greeks to Darwin: An Outline of the Development of the Evolution Idea; Macmillan and Co.: New York, NY, USA, 1894; p. 284. [Google Scholar]
- Zirkle, C. The early history of the idea of the inheritance of acquired characters and of pangenesis. Trans. Am. Phil. Soc. 1946, 35, 91–151. [Google Scholar] [CrossRef]
- Portera, M.; Mandrioli, M. Who’s afraid of epigenetics? Habits, instincts, and Charles Darwin’s evolutionary theory. Hist. Philos. Life Sci. 2021, 43, 20. [Google Scholar] [CrossRef] [PubMed]
- Barsanti, G. Lamarck and the birth of Biology (1740–1810). In Romanticism in Science. Science in Europe, 1790–1840. Boston Studies in the Philosophy and History of Science; Poggi, S., Bossi, M., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 47–74. [Google Scholar]
- Loison, L. Epigenetic inheritance and evolution: A historian’s perspective. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, e20200120. [Google Scholar] [CrossRef]
- La Vergata, A. L’evoluzione Biologica: Da Linneo a Darwin. Loescher Editore; UTET: Torino, Italy, 1979; pp. 12–13. [Google Scholar]
- Burkhardt, R.W. The Spirit of System: Lamarck and Evolutionary Biology; Harvard University Press: Cambridge, UK, 1977. [Google Scholar]
- Mandrioli, M. At the origin of the inheritance of acquired characters: From Erasmus Darwin to Jean Baptiste de Lamarck to remember their true legacy. Sci. Filos. 2022, 28, 26–38. [Google Scholar]
- Rabaud, E. La Tératogenèse: Étude des Variations de L’organisme; Octave Doin: Paris, France, 1914. [Google Scholar]
- Cairns, J.; Overbaugh, J.; Miller, S. The origin of mutants. Nature 1988, 335, 142–145. [Google Scholar] [CrossRef]
- Gillis, A.M. Can organisms direct their evolution? BioScience 1991, 41, 202–205. [Google Scholar] [CrossRef]
- Lenski, R.E.; Mittler, J.E. The directed mutation controversy and neo-Darwinism. Science 1993, 259, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Sniegowski, P.D.; Lenski, R.E. Mutation and adaptation: The directed mutation controversy in evolutionary perspective. Ann. Rev. Ecol. Syst. 1995, 26, 553–578. [Google Scholar] [CrossRef]
- Foster, P.L. Adaptive mutation: Implications for evolution. Bioessays 2000, 22, 1067–1074. [Google Scholar] [CrossRef] [PubMed]
- Laland, K.; Uller, T.; Feldman, M.; Sterelny, K.; Müller, G.B.; Moczek, A.; Jablonka, E.; Odling-Smee, J.; Wray, G.A.; Hoekstra, H.E.; et al. Does evolutionary theory need a rethink? Nature 2014, 514, 161–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, M.K. Environmental epigenetics and a unified theory of the molecular aspects of evolution: A neo-Lamarckian concept that facilitates neo-Darwinian evolution. Genome Biol. Evol. 2015, 7, 1296–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigliucci, M. Evolution of phenotypic plasticity: Where are we going now? Trends Ecol. Evol. 2005, 20, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Huneman, P. Revisiting the modern synthesis. J. Hist. Biol. 2019, 52, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Camacho, M.P. What’s all the fuss about? The inheritance of acquired traits is compatible with the Central Dogma. Hist. Phil. Life Sci. 2020, 42, 32. [Google Scholar] [CrossRef]
- Skinner, M.K.; Nilsson, E.E. Role of environmentally induced epigenetic transgenerational inheritance in evolutionary biology: Unified Evolution Theory. Environ. Epigenet. 2021, 7, dvab012. [Google Scholar] [CrossRef]
- Darwin, C. The Variation of Animals and Plants under Domestication; John Murray: London, UK, 1868. [Google Scholar]
- Lakatos, I.; Musgrave, A. Criticism and the Growth of Knowledge; Cambridge University Press: Cambridge, UK, 1970. [Google Scholar]
- Brooks, D.R. The Mastodon in the room: How Darwinian is neo-Darwinism? Stud. Hist. Philos. Biol. Biomed. Sci. 2011, 42, 82–88. [Google Scholar] [CrossRef]
- Hogeweg, P. Toward a theory of multilevel evolution: Long-term information integration shapes the mutational landscape and enhances evolvability. Adv. Exp. Med. Biol. 2012, 751, 195–224. [Google Scholar]
- Pievani, T. Individuals and groups in evolution: Darwinian pluralism and the multilevel selection debate. J. Biosci. 2014, 39, 319–325. [Google Scholar] [CrossRef]
- Brown, O.R.; Hullender, D.A. Neo-Darwinism must mutate to survive. Prog. Biophys. Mol. Biol. 2022, 172, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Darwinian evolution includes Lamarckian inheritance of acquired characters. Int. J. Epidemiol. 2016, 45, 2206–2207. [Google Scholar]
- Penny, D. Epigenetics, Darwin, and Lamarck. Genome Biol. Evol. 2015, 7, 1758–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henikoff, S. Darwin meets Waddington. Curr. Biol. 2018, 28, R679–R694. [Google Scholar] [CrossRef] [Green Version]
- Fanti, L.; Piacentini, L.; Cappucci, U.; Casale, A.M.; Pimpinelli, S. Canalization by selection of de novo induced mutations. Genetics 2017, 206, 1995–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klosin, A.; Lehner, B. Mechanisms, timescales and principles of trans-generational epigenetic inheritance in animals. Curr. Opin. Genet. Dev. 2016, 36, 41–49. [Google Scholar] [CrossRef]
- Di Croce, L.; Shilatifard, A. Editorial overview: Cancer genomics: Darwin meets Waddington: The interplay between cancer genomes and epigenomes. Curr. Opin. Genet. Dev. 2016, 36, iv–vi. [Google Scholar] [CrossRef]
- Waddington, C.H. The Strategy of the Genes; Routledge: London, UK, 1957; p. 274. [Google Scholar]
- Rajagopal, J.; Stanger, B.Z. Plasticity in the adult: How should the Waddington diagram be applied to regenerating tissues? Dev. Cell 2016, 36, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Holmes, S.G.; Broach, J.R. Silencers are required for inheritance of the repressed state in yeast. Genes Dev. 1996, 10, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.H.; Chang, C.R.; Joy, P.; Yablok, S.; Gartenberg, M.R. Controlling gene expression in yeast by inducible site-specific recombination. Nucleic Acids Res. 2000, 28, e108. [Google Scholar] [CrossRef] [Green Version]
- Moazed, D. Mechanisms for the inheritance of chromatin states. Cell 2011, 146, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Joh, R.I.; Palmieri, C.M.; Hill, I.T.; Motamedi, M. Regulation of histone methylation by noncoding RNAs. Biochim. Biophys. Acta 2014, 1839, 1385–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busturia, A.; Wightman, C.D.; Sakonju, S. A silencer is required for maintenance of transcriptional repression throughout Drosophila development. Development 1997, 124, 4343–4350. [Google Scholar] [CrossRef] [PubMed]
- Jablonka, E.; Lamb, M.J. Evolution in Four Dimensions; MIT Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Geoghegen, J.L.; Spencer, H.G. Population-epigenetic models of selection. Theor. Popul. Biol. 2012, 81, 232–242. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandrioli, M. From Environmental Epigenetics to the Inheritance of Acquired Traits: A Historian and Molecular Perspective on an Unnecessary Lamarckian Explanation. Biomolecules 2023, 13, 1077. https://doi.org/10.3390/biom13071077
Mandrioli M. From Environmental Epigenetics to the Inheritance of Acquired Traits: A Historian and Molecular Perspective on an Unnecessary Lamarckian Explanation. Biomolecules. 2023; 13(7):1077. https://doi.org/10.3390/biom13071077
Chicago/Turabian StyleMandrioli, Mauro. 2023. "From Environmental Epigenetics to the Inheritance of Acquired Traits: A Historian and Molecular Perspective on an Unnecessary Lamarckian Explanation" Biomolecules 13, no. 7: 1077. https://doi.org/10.3390/biom13071077