
Novel Peptides with Dual Properties for Treating Pseudomonas aeruginosa Keratitis: Antibacterial and Corneal Wound Healing
 , , , ,
, , , ,  , ,
, ,  ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Peptides
2.2. Bacterial Strains and Antimicrobial Activity
2.3. In Vivo Infection
2.4. In Vivo Corneal Debridement Wound
2.5. Human Corneal Epithelial Cell Culture Conditions
2.6. In Vitro Cell Scratch Assay
2.7. Proliferation Assay
2.8. Cell Viability Assay
2.9. Enzyme-Linked Immunosorbent Assays (ELISA)
2.10. Statistical Analyses
3. Results
3.1. Antipseudomonal Activity of Esc Peptides In Vitro and in a Mouse Model of Keratitis
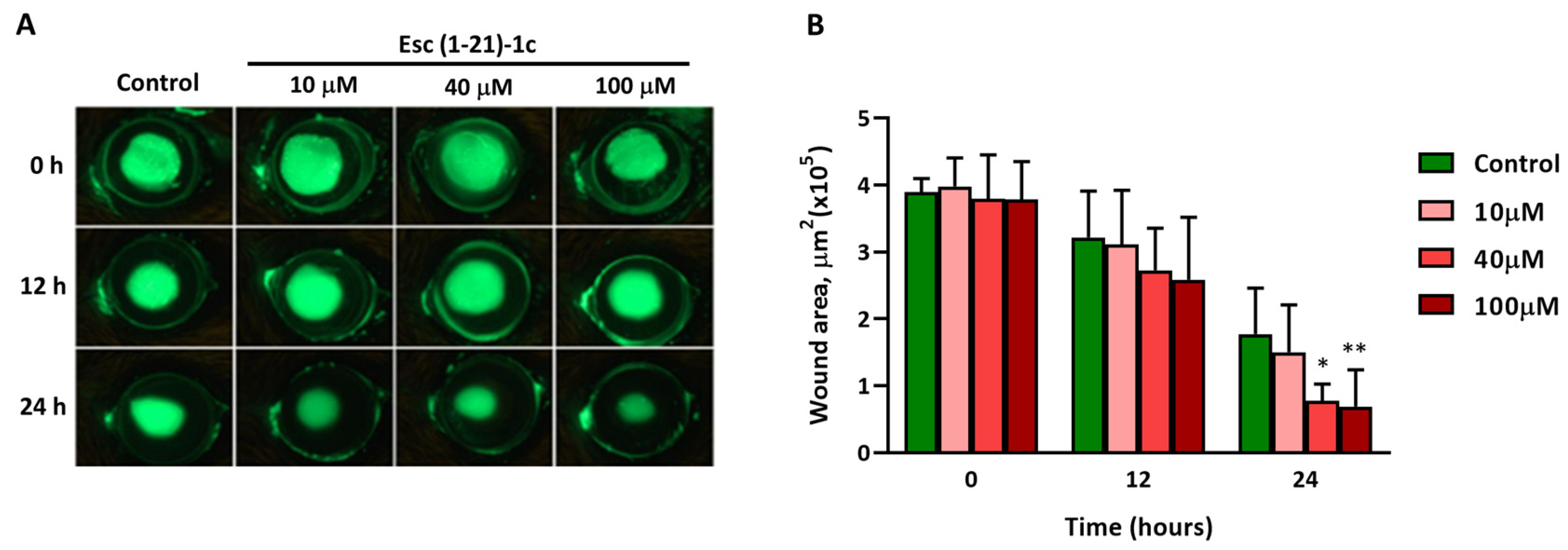
3.2. In Vivo Corneal Wound Healing
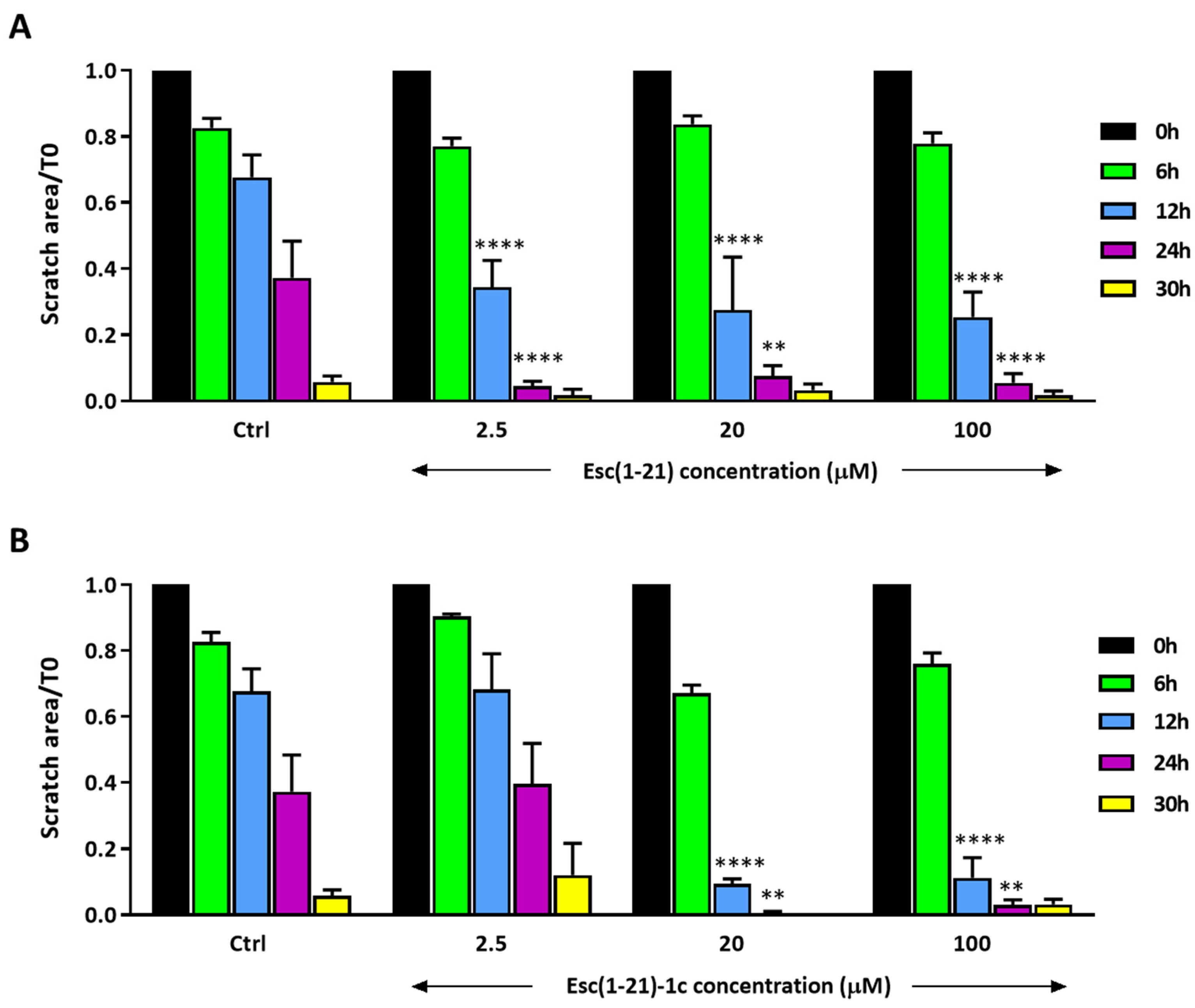
3.3. In Vitro Wound Healing on Human Corneal Epithelial Cells
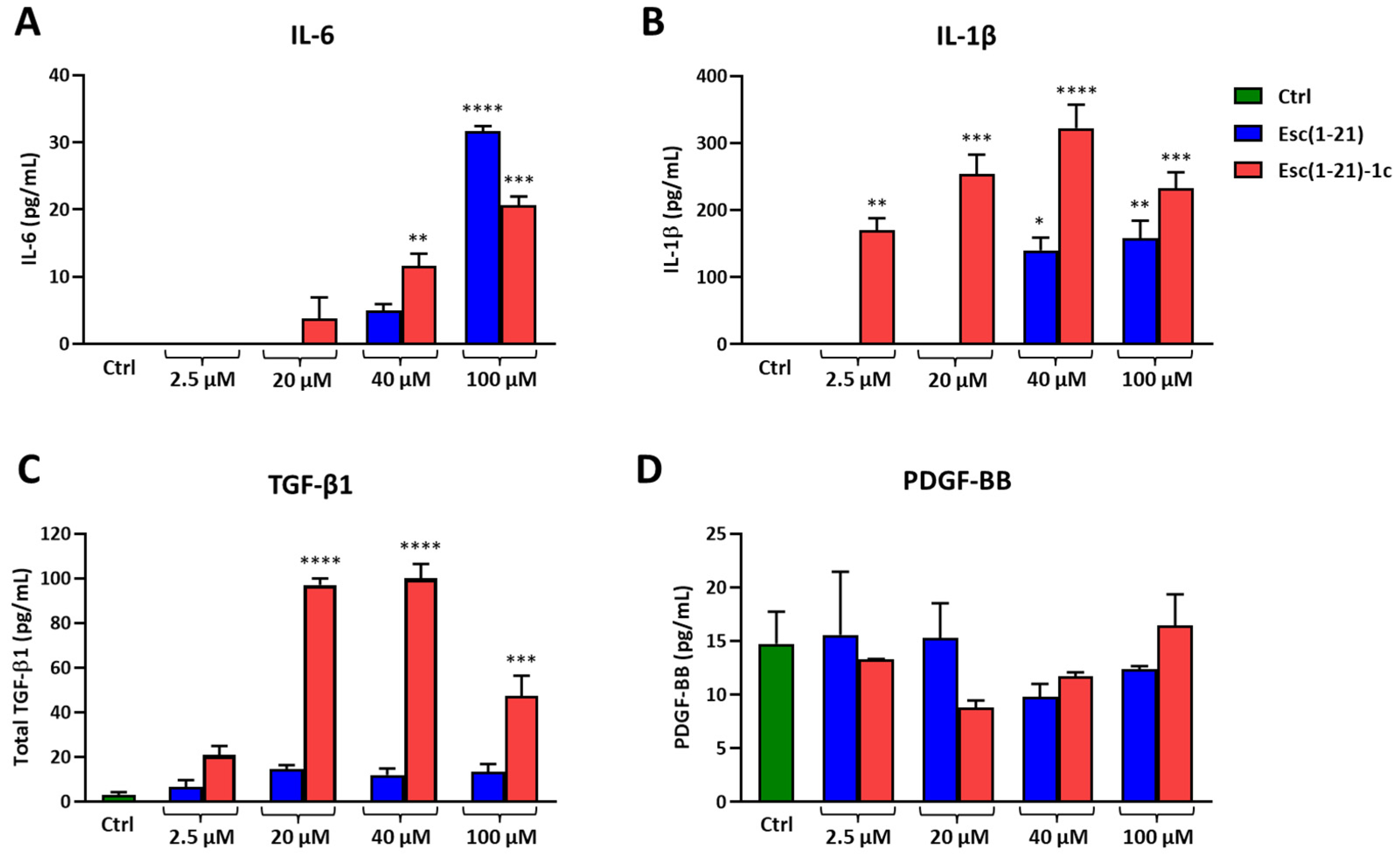
3.4. Cytokine Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leong, Y.Y.; Tong, L. Barrier function in the ocular surface: From conventional paradigms to new opportunities. Ocul. Surf. 2015, 13, 103–109. [Google Scholar] [CrossRef]
- Fleiszig, S.M.J.; Kroken, A.R.; Nieto, V.; Grosser, M.R.; Wan, S.J.; Metruccio, M.M.E.; Evans, D.J. Contact lens-related corneal infection: Intrinsic resistance and its compromise. Prog. Retin. Eye Res. 2020, 76, 100804. [Google Scholar] [CrossRef]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef]
- Green, M.; Apel, A.; Stapleton, F. Risk factors and causative organisms in microbial keratitis. Cornea 2008, 27, 22–27. [Google Scholar] [CrossRef]
- Schein, O.D.; McNally, J.J.; Katz, J.; Chalmers, R.L.; Tielsch, J.M.; Alfonso, E.; Bullimore, M.; O’Day, D.; Shovlin, J. The incidence of microbial keratitis among wearers of a 30-day silicone hydrogel extended-wear contact lens. Ophthalmology 2005, 112, 2172–2179. [Google Scholar] [CrossRef]
- Ganz, T. Antimicrobial polypeptides. J. Leukoc. Biol. 2003, 75, 34–38. [Google Scholar] [CrossRef]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef]
- Mangoni, M.L. Temporins, anti-infective peptides with expanding properties. Cell. Mol. Life Sci. 2006, 63, 1060–1069. [Google Scholar] [CrossRef]
- Conlon, J.M.; Iwamuro, S.; King, J.D. Dermal cytolytic peptides and the system of innate immunity in anurans. Ann. N. Y. Acad. Sci. 2009, 1163, 75–82. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Barra, D. Antimicrobial peptides from amphibian skin: What do they tell us? Biopolymers 1998, 47, 435–450. [Google Scholar] [CrossRef]
- Islas-Rodriguez, A.E.; Marcellini, L.; Orioni, B.; Barra, D.; Stella, L.; Mangoni, M.L. Esculentin 1–21: A linear antimicrobial peptide from frog skin with inhibitory effect on bovine mastitis-causing bacteria. J. Pept. Sci. 2009, 15, 607–614. [Google Scholar] [CrossRef]
- Luca, V.; Stringaro, A.; Colone, M.; Pini, A.; Mangoni, M.L. Esculentin(1–21), an amphibian skin membrane-active peptide with potent activity on both planktonic and biofilm cells of the bacterial pathogen Pseudomonas aeruginosa. Cell. Mol. Life Sci. 2013, 70, 2773–2786. [Google Scholar] [CrossRef]
- Casciaro, B.; Loffredo, M.R.; Luca, V.; Verrusio, W.; Cacciafesta, M.; Mangoni, M.L. Esculentin-1a Derived Antipseudomonal Peptides: Limited Induction of Resistance and Synergy with Aztreonam. Protein Pept. Lett. 2018, 25, 1155–1162. [Google Scholar] [CrossRef]
- Kolar, S.S.N.; Luca, V.; Baidouri, H.; Mannino, G.; McDermott, A.M.; Mangoni, M.L. Esculentin-1a(1-21)NH2: A frog skin-derived peptide for microbial keratitis. Cell. Mol. Life Sci. 2015, 72, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Mangoni, M.L.; Di, Y.P. In vivo therapeutic efficacy of frog skin-derived peptides against Pseudomonas aeruginosa-induced pulmonary infection. Sci. Rep. 2017, 7, 8548. [Google Scholar] [CrossRef] [Green Version]
- Di Grazia, A.; Cappiello, F.; Cohen, H.; Casciaro, B.; Luca, V.; Pini, A.; Di, Y.P.; Shai, Y.; Mangoni, M.L. D-Amino acids incorporation in the frog skin-derived peptide esculentin-1a(1-21)NH2 is beneficial for its multiple functions. Amino Acids 2015, 47, 2505–2519. [Google Scholar] [CrossRef]
- Casciaro, B.; Dutta, D.; Loffredo, M.R.; Marcheggiani, S.; McDermott, A.M.; Willcox, M.D.; Mangoni, M.L. Esculentin-1a derived peptides kill Pseudomonas aeruginosa biofilm on soft contact lenses and retain antibacterial activity upon immobilization to the lens surface. Biopolymers 2018, 110, e23074. [Google Scholar] [CrossRef]
- Cappiello, F.; Casciaro, B.; Loffredo, M.R.; Puglisi, E.; Lin, Q.; Yang, D.; Conte, G.; d’Angelo, I.; Ungaro, F.; Ferrera, L.; et al. Pulmonary Safety Profile of Esc Peptides and Esc-Peptide-Loaded Poly(lactide-co-glycolide) Nanoparticles: A Promising Therapeutic Approach for Local Treatment of Lung Infectious Diseases. Pharmaceutics 2022, 14, 2297. [Google Scholar] [CrossRef]
- Zhu, H.; Bandara, R.; Conibear, T.C.; Thuruthyil, S.J.; Rice, S.A.; Kjelleberg, S.; Givskov, M.; Willcox, M.D. Pseudomonas aeruginosa with lasI quorum-sensing deficiency during corneal infection. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1897–1903. [Google Scholar] [CrossRef] [Green Version]
- Dutta, D.; Willcox, M.D. A laboratory assessment of factors that affect bacterial adhesion to contact lenses. Biology 2013, 2, 1268–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, D.M.; Li, L.; Fisher, S.; Pearce, V.P.; Shay, J.W.; Wright, W.E.; Cavanagh, H.D.; Jester, J.V. Characterization of growth and differentiation in a telomerase-immortalized human corneal epithelial cell line. Investig. Ophthalmol. Vis. Sci. 2005, 46, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Cappiello, F.; Di Grazia, A.; Segev-Zarko, L.A.; Scali, S.; Ferrera, L.; Galietta, L.; Pini, A.; Shai, Y.; Di, Y.P.; Mangoni, M.L. Esculentin-1a-Derived Peptides Promote Clearance of Pseudomonas aeruginosa Internalized in Bronchial Cells of Cystic Fibrosis Patients and Lung Cell Migration: Biochemical Properties and a Plausible Mode of Action. Antimicrob. Agents Chemother. 2016, 60, 7252–7262. [Google Scholar] [CrossRef] [Green Version]
- Di Grazia, A.; Cappiello, F.; Imanishi, A.; Mastrofrancesco, A.; Picardo, M.; Paus, R.; Mangoni, M.L. The Frog Skin-Derived Antimicrobial Peptide Esculentin-1a(1-21)NH2 Promotes the Migration of Human HaCaT Keratinocytes in an EGF Receptor-Dependent Manner: A Novel Promoter of Human Skin Wound Healing? PLoS ONE 2015, 10, e0128663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reins, R.Y.; Baidouri, H.; McDermott, A.M. Vitamin D Activation and Function in Human Corneal Epithelial Cells During TLR-Induced Inflammation. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7715–7727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, F.W.; Gallagher, C.; O’Reilly, N.; Clynes, M.; O’Sullivan, F.; Kavanagh, K. Exposure of a corneal epithelial cell line (hTCEpi) to Demodex-associated Bacillus proteins results in an inflammatory response. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7019–7028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, A.; Lietman, T.; Rose-Nussbaumer, J. Update on the Management of Infectious Keratitis. Ophthalmology 2017, 124, 1678–1689. [Google Scholar] [CrossRef]
- Khoo, P.; Cabrera-Aguas, M.P.; Nguyen, V.; Lahra, M.M.; Watson, S.L. Microbial keratitis in Sydney, Australia: Risk factors, patient outcomes, and seasonal variation. Graefe’s Arch. Clin. Exp. Ophthalmol. 2020, 258, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Suzuki, T.; Kobayashi, T.; Oka, N.; Ishikawa, E.; Shinomiya, H.; Ohashi, Y. Genotypic analysis of Pseudomonas aeruginosa isolated from ocular infection. J. Infect. Chemother. 2014, 20, 407–411. [Google Scholar] [CrossRef]
- Szczotka-Flynn, L.B.; Shovlin, J.P.; Schnider, C.M.; Caffery, B.E.; Alfonso, E.C.; Carnt, N.A.; Chalmers, R.L.; Collier, S.; Jacobs, D.S.; Joslin, C.E.; et al. American Academy of Optometry Microbial Keratitis Think Tank. Optom. Vis. Sci. 2021, 98, 182–198. [Google Scholar] [CrossRef]
- Cabrera-Aguas, M.; Khoo, P.; Watson, S.L. Infectious keratitis: A review. Clin. Exp. Ophthalmol. 2022, 50, 543–562. [Google Scholar] [CrossRef] [PubMed]
- Jadi, P.K.; Sharma, P.; Bhogapurapu, B.; Roy, S. Alternative Therapeutic Interventions: Antimicrobial Peptides and Small Molecules to Treat Microbial Keratitis. Front. Chem. 2021, 9, 694998. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial peptides and wound healing: Biological and therapeutic considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Carretero, M.; Escamez, M.J.; Garcia, M.; Duarte, B.; Holguin, A.; Retamosa, L.; Jorcano, J.L.; Rio, M.D.; Larcher, F. In vitro and in vivo wound healing-promoting activities of human cathelicidin LL-37. J. Investig. Dermatol. 2008, 128, 223–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Guo, J.L.; Han, J.; Si, R.J.; Liu, P.P.; Zhang, Z.R.; Wang, A.M.; Zhang, J. Chitosan hydrogel encapsulated with LL-37 peptide promotes deep tissue injury healing in a mouse model. Mil. Med. Res. 2020, 7, 20. [Google Scholar] [CrossRef]
- Belvedere, R.; Pessolano, E.; Porta, A.; Tosco, A.; Parente, L.; Petrella, F.; Perretti, M.; Petrella, A. Mesoglycan induces the secretion of microvesicles by keratinocytes able to activate human fibroblasts and endothelial cells: A novel mechanism in skin wound healing. Eur. J. Pharmacol. 2020, 869, 172894. [Google Scholar] [CrossRef]
- Huang, H.N.; Rajanbabu, V.; Pan, C.Y.; Chan, Y.L.; Wu, C.J.; Chen, J.Y. Use of the antimicrobial peptide Epinecidin-1 to protect against MRSA infection in mice with skin injuries. Biomaterials 2013, 34, 10319–10327. [Google Scholar] [CrossRef]
- Cappiello, F.; Ranieri, D.; Carnicelli, V.; Casciaro, B.; Chen, H.T.; Ferrera, L.; Di, Y.P.; Mangoni, M.L. Bronchial epithelium repair by Esculentin-1a-derived antimicrobial peptides: Involvement of metalloproteinase-9 and interleukin-8, and evaluation of peptides’ immunogenicity. Sci. Rep. 2019, 9, 18988. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Chen, C.; Lin, Z.; Zhang, L.; Guan, S.; Zhuang, X.; Dong, G.; Shen, J. The Antimicrobial Peptide Esculentin-1a(1-21)NH2 Stimulates Wound Healing by Promoting Angiogenesis through the PI3K/AKT Pathway. Biol. Pharm. Bull. 2023, 46, 382–393. [Google Scholar] [CrossRef]
- Casciaro, B.; d’Angelo, I.; Zhang, X.; Loffredo, M.R.; Conte, G.; Cappiello, F.; Quaglia, F.; Di, Y.P.; Ungaro, F.; Mangoni, M.L. Poly(lactide-co-glycolide) Nanoparticles for Prolonged Therapeutic Efficacy of Esculentin-1a-Derived Antimicrobial Peptides against Pseudomonas aeruginosa Lung Infection: In Vitro and in Vivo Studies. Biomacromolecules 2019, 20, 1876–1888. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, L.; Su, L.; van der Mei, H.C.; Jutte, P.C.; Ren, Y.; Busscher, H.J. Nanotechnology-based antimicrobials and delivery systems for biofilm-infection control. Chem. Soc. Rev. 2019, 48, 428–446. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Piao, Y.Z.; Chen, H.; Shi, K.; Dai, J.; Wang, S.; Zhou, T.; Le, A.T.; Wang, Y.; Wu, F.; et al. Dynamic covalent nano-networks comprising antibiotics and polyphenols orchestrate bacterial drug resistance reversal and inflammation alleviation. Bioact. Mater. 2023, 27, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Makabenta, J.M.V.; Nabawy, A.; Li, C.H.; Schmidt-Malan, S.; Patel, R.; Rotello, V.M. Nanomaterial-based therapeutics for antibiotic-resistant bacterial infections. Nat. Rev. Microbiol. 2021, 19, 23–36. [Google Scholar] [CrossRef]
- Ljubimov, A.V.; Saghizadeh, M. Progress in corneal wound healing. Prog. Retin. Eye Res. 2015, 49, 17–45. [Google Scholar] [CrossRef] [Green Version]
- Fortingo, N.; Melnyk, S.; Sutton, S.H.; Watsky, M.A.; Bollag, W.B. Innate Immune System Activation, Inflammation and Corneal Wound Healing. Int. J. Mol. Sci. 2022, 23, 14933. [Google Scholar] [CrossRef]
- Ebihara, N.; Matsuda, A.; Nakamura, S.; Matsuda, H.; Murakami, A. Role of the IL-6 classic- and trans-signaling pathways in corneal sterile inflammation and wound healing. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8549–8557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisjoly, H.M.; Laplante, C.; Bernatchez, S.F.; Salesse, C.; Giasson, M.; Joly, M.C. Effects of EGF, IL-1 and their combination on in vitro corneal epithelial wound closure and cell chemotaxis. Exp. Eye Res. 1993, 57, 293–300. [Google Scholar] [CrossRef]
- Huang, L.C.; Petkova, T.D.; Reins, R.Y.; Proske, R.J.; McDermott, A.M. Multifunctional roles of human cathelicidin (LL-37) at the ocular surface. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2369–2380. [Google Scholar] [CrossRef]
- O’Kane, S.; Ferguson, M.W. Transforming growth factor beta s and wound healing. Int. J. Biochem. Cell Biol. 1997, 29, 63–78. [Google Scholar] [CrossRef]
- Saika, S. TGF-beta signal transduction in corneal wound healing as a therapeutic target. Cornea 2004, 23, S25–S30. [Google Scholar] [CrossRef]
- Tandon, A.; Tovey, J.C.; Sharma, A.; Gupta, R.; Mohan, R.R. Role of transforming growth factor Beta in corneal function, biology and pathology. Curr. Mol. Med. 2010, 10, 565–578. [Google Scholar] [CrossRef]
- Wilson, S.E. Corneal wound healing. Exp. Eye Res. 2020, 197, 108089. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Esposito, A. Focus on molecules: Interleukin-1: A master regulator of the corneal response to injury. Exp. Eye Res. 2009, 89, 124–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.H.; Hsieh, T.L.; Ji, A.T.; Hsu, W.T.; Liu, C.Y.; Lee, O.K.; Ho, J.H. Stromal Tissue Rigidity Promotes Mesenchymal Stem Cell-Mediated Corneal Wound Healing Through the Transforming Growth Factor beta Signaling Pathway. Stem Cells 2016, 34, 2525–2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, D.; Son, K.N.; Kalmodia, S.; Lee, B.S.; Ali, M.; Balasubramaniam, A.; Shukla, D.; Aakalu, V.K. Wound Healing Properties of Histatin-5 and Identification of a Functional Domain Required for Histatin-5-Induced Cell Migration. Mol. Ther. Methods Clin. Dev. 2020, 17, 709–716. [Google Scholar] [CrossRef]
- Nos-Barbera, S.; Portoles, M.; Morilla, A.; Ubach, J.; Andreu, D.; Paterson, C.A. Effect of hybrid peptides of cecropin A and melittin in an experimental model of bacterial keratitis. Cornea 1997, 16, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Kasus-Jacobi, A.; Noor-Mohammadi, S.; Griffith, G.L.; Hinsley, H.; Mathias, L.; Pereira, H.A. A multifunctional peptide based on the neutrophil immune defense molecule, CAP37, has antibacterial and wound-healing properties. J. Leukoc. Biol. 2015, 97, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Ting, D.S.J.; Goh, E.T.L.; Mayandi, V.; Busoy, J.M.F.; Aung, T.T.; Periayah, M.H.; Nubile, M.; Mastropasqua, L.; Said, D.G.; Htoon, H.M.; et al. Hybrid derivative of cathelicidin and human beta defensin-2 against Gram-positive bacteria: A novel approach for the treatment of bacterial keratitis. Sci. Rep. 2021, 11, 18304. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappiello, F.; Verma, S.; Lin, X.; Moreno, I.Y.; Casciaro, B.; Dutta, D.; McDermott, A.M.; Willcox, M.; Coulson-Thomas, V.J.; Mangoni, M.L. Novel Peptides with Dual Properties for Treating Pseudomonas aeruginosa Keratitis: Antibacterial and Corneal Wound Healing. Biomolecules 2023, 13, 1028. https://doi.org/10.3390/biom13071028
Cappiello F, Verma S, Lin X, Moreno IY, Casciaro B, Dutta D, McDermott AM, Willcox M, Coulson-Thomas VJ, Mangoni ML. Novel Peptides with Dual Properties for Treating Pseudomonas aeruginosa Keratitis: Antibacterial and Corneal Wound Healing. Biomolecules. 2023; 13(7):1028. https://doi.org/10.3390/biom13071028
Chicago/Turabian StyleCappiello, Floriana, Sudhir Verma, Xiao Lin, Isabel Y. Moreno, Bruno Casciaro, Debarun Dutta, Alison M. McDermott, Mark Willcox, Vivien J. Coulson-Thomas, and Maria Luisa Mangoni. 2023. "Novel Peptides with Dual Properties for Treating Pseudomonas aeruginosa Keratitis: Antibacterial and Corneal Wound Healing" Biomolecules 13, no. 7: 1028. https://doi.org/10.3390/biom13071028