
Deciphering Plant-Insect-Microorganism Signals for Sustainable Crop Production
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
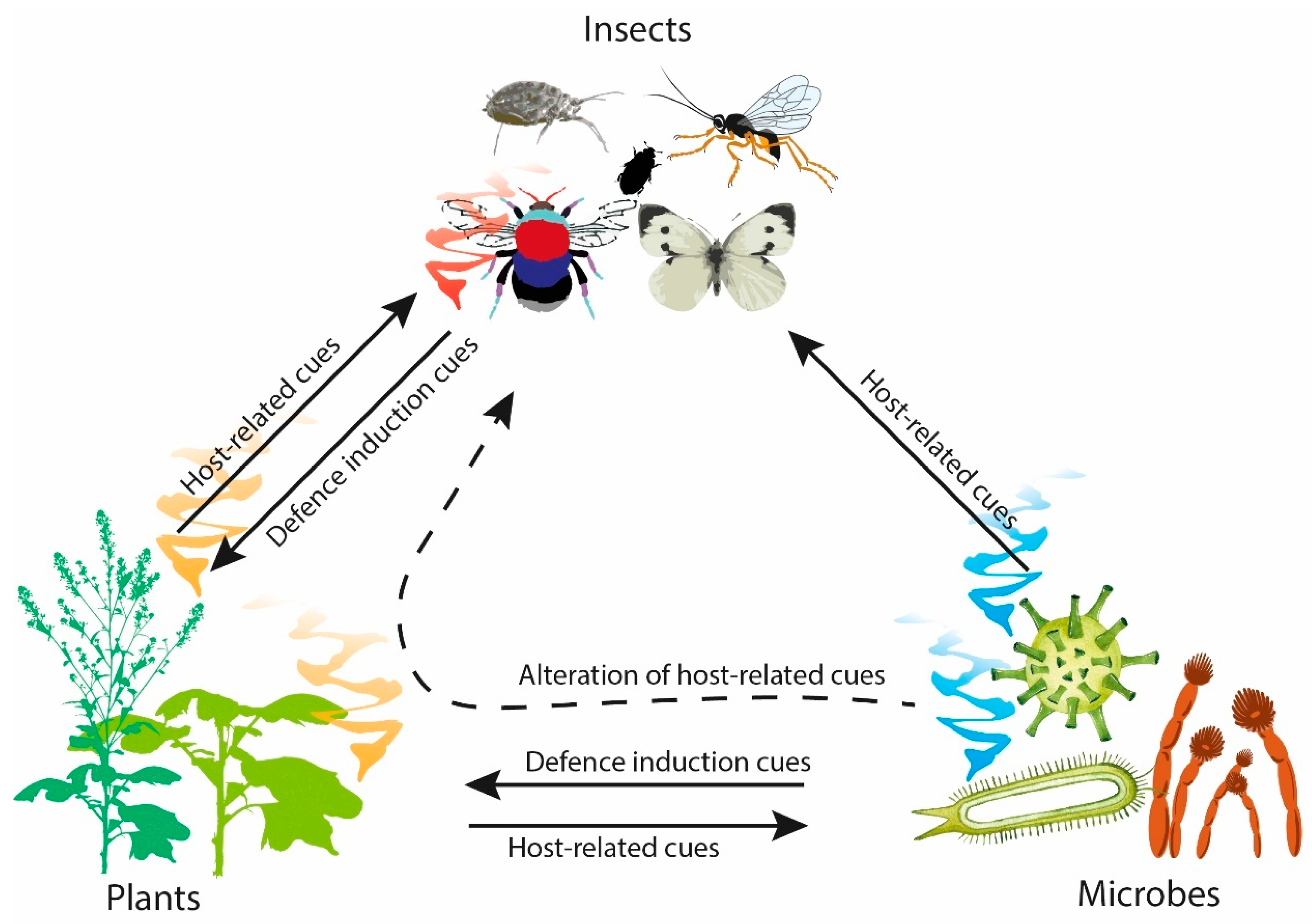
2. Plant-Insect Signals
2.1. Plant VOCs Induction by Insect Pests
2.2. Exogenous Application of Natural or Synthetic Compounds
2.3. Crop Genetic Diversity and VOC Emissions
2.4. Insect Pheromone Perception by Plants
3. Insect-Microbe Signals
3.1. Pre-Harvest Pest Management
3.2. Post-Harvest Pest Management
4. Plant-Microbe Signals
4.1. Plant Defence Induction by VOCs of Soil-Borne Microbes against Plant Pathogens
4.2. Microbe-Induced Plant Volatiles
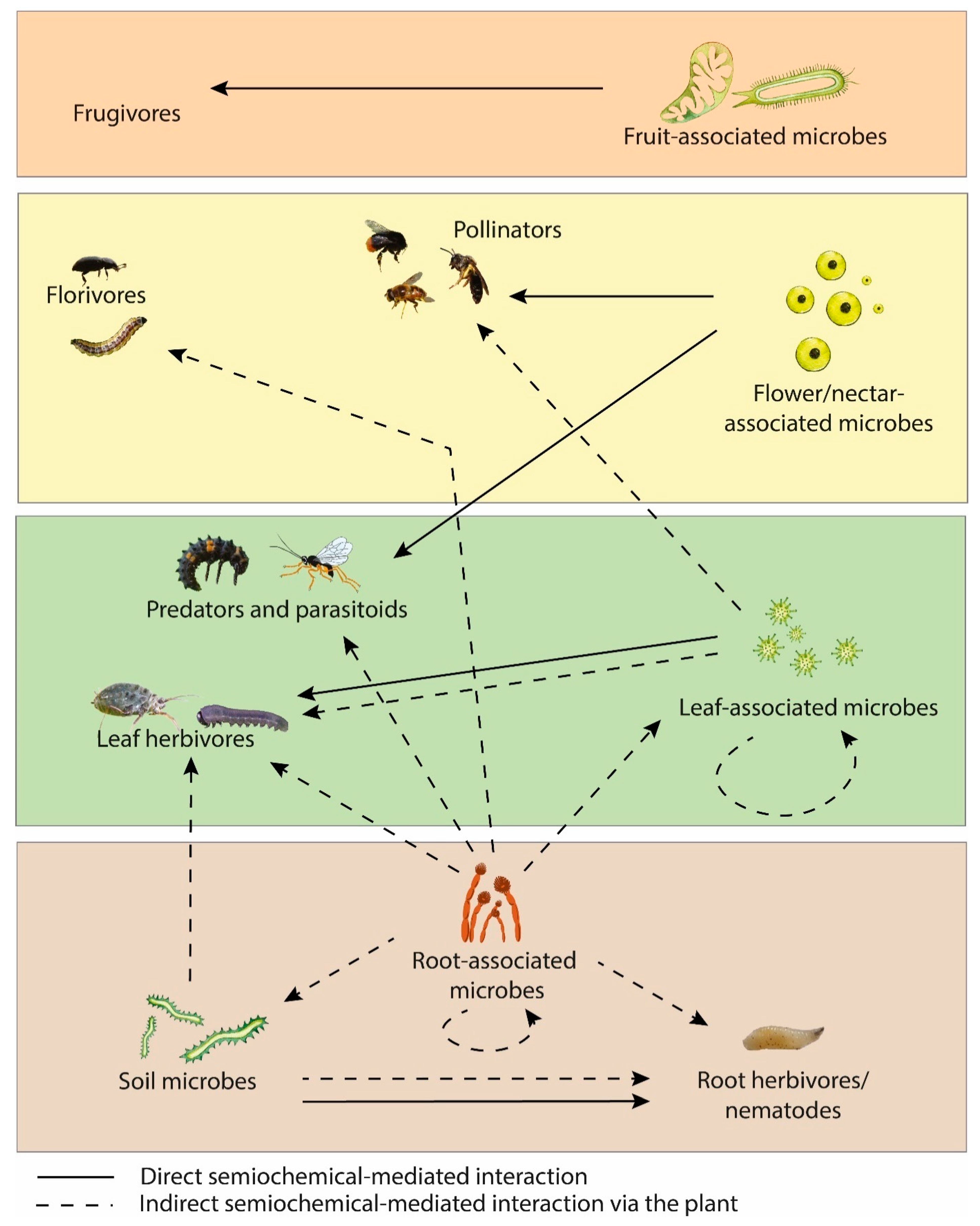
5. Plant-Microbe-Insect Signals
5.1. Influence of Plant-Microbe Interactions on Herbivores
5.2. Influence of Plant-Microbe Interactions on Natural Enemies’ Behaviour
5.3. Influence of Plant-Microbe Interactions on Pollinator Behaviour
5.4. Influence of Insect-Associated Microbes on Plant VOC Emissions
6. Application of Semiochemicals to Improve Sustainability in Agriculture
6.1. Examples of Pest Management Products Based on Semiochemicals
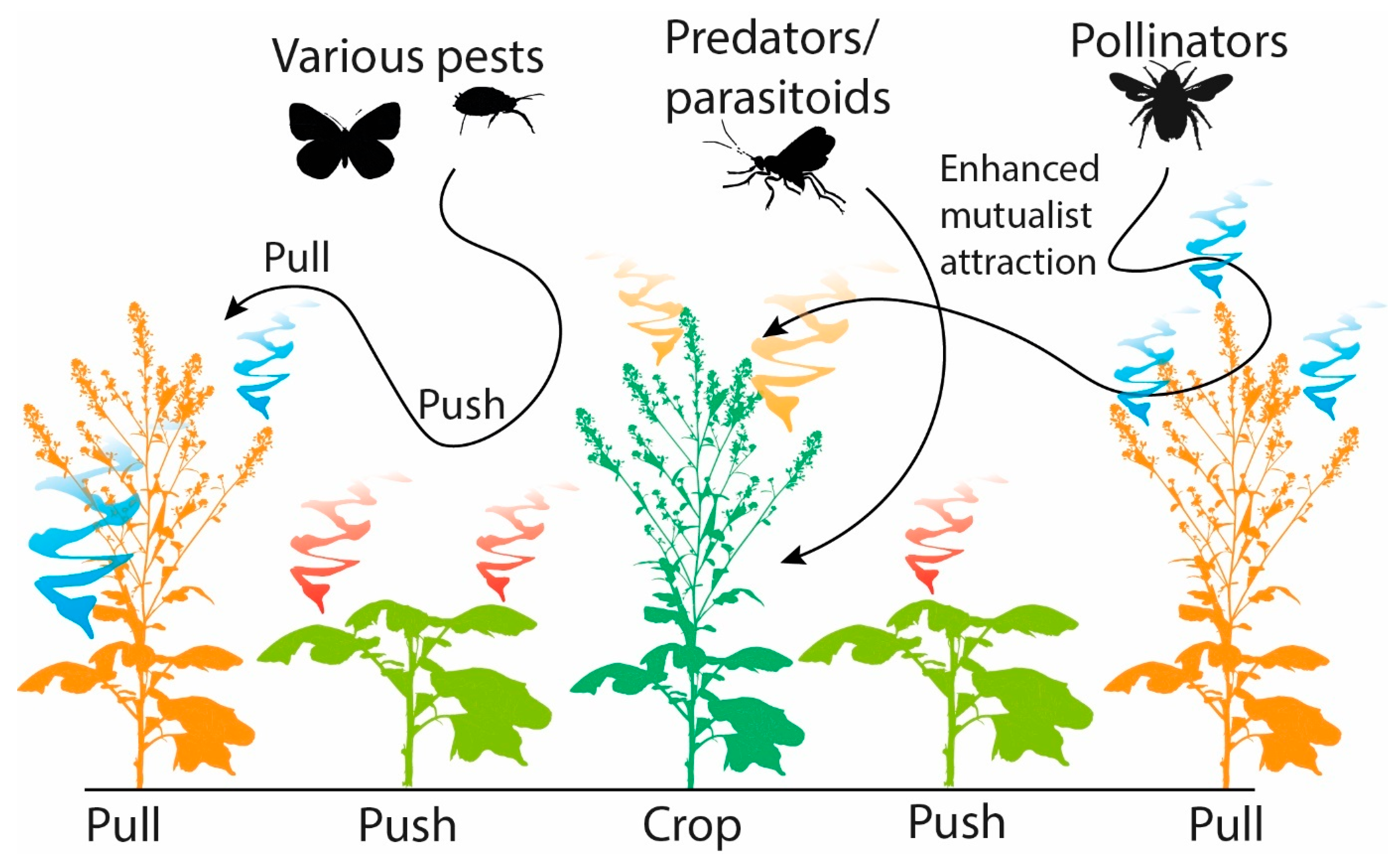
6.2. Push-Pull
6.3. Transgenic Approaches for Insect Pheromone Biosynthesis in Plants
7. Future Perspectives and Research Priorities
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Osei-Owusu, J.; Vuts, J.; Caulfield, J.C.; Woodcock, C.M.; Withall, D.M.; Hooper, A.M.; Osafo-Acquaah, S.; Birkett, M.A. Identification of Semiochemicals from Cowpea, Vigna unguiculata, for Low-Input Management of the Legume Pod Borer, Maruca vitrata. J. Chem. Ecol. 2020, 46, 288–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
- Raza, M.M.; Bebber, D.P. Climate Change and Plant Pathogens. Curr. Opin. Microbiol. 2022, 70, 102233. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Guerra, C.A.; Cano-Díaz, C.; Egidi, E.; Wang, J.-T.; Eisenhauer, N.; Singh, B.K.; Maestre, F.T. The Proportion of Soil-Borne Pathogens Increases with Warming at the Global Scale. Nat. Clim. Chang. 2020, 10, 550–554. [Google Scholar] [CrossRef]
- Gould, F.; Brown, Z.S.; Kuzma, J. Wicked Evolution: Can We Address the Sociobiological Dilemma of Pesticide Resistance? Science 2018, 360, 728–732. [Google Scholar] [CrossRef] [Green Version]
- Lamberth, C.; Jeanmart, S.; Luksch, T.; Plant, A. Current Challenges and Trends in the Discovery of Agrochemicals. Science 2013, 341, 742–746. [Google Scholar] [CrossRef] [PubMed]
- Smith, S. Combinatorial Chemistry in the Development of New Crop Protection Products. Pestic. Outlook 2003, 14, 21–26. [Google Scholar] [CrossRef]
- Law, J.H.; Regnier, F.E. Pheromones. Annu. Rev. Biochem. 1971, 40, 533–548. [Google Scholar] [CrossRef]
- Pickett, J.A.; Aradottír, G.I.; Birkett, M.A.; Bruce, T.J.A.; Hooper, A.M.; Midega, C.A.O.; Jones, H.D.; Matthes, M.C.; Napier, J.A.; Pittchar, J.O.; et al. Delivering Sustainable Crop Protection Systems via the Seed: Exploiting Natural Constitutive and Inducible Defence Pathways. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20120281. [Google Scholar] [CrossRef] [Green Version]
- Schlaeger, S.; Pickett, J.A.; Birkett, M.A. Prospects for Management of Whitefly Using Plant Semiochemicals, Compared with Related Pests. Pest Manag. Sci. 2018, 74, 2405–2411. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A.; Pickett, J.A. Perception of Plant Volatile Blends by Herbivorous Insects—Finding the Right Mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Hassemer, M.J.; Borges, M.; Withall, D.M.; Pickett, J.A.; Laumann, R.A.; Birkett, M.A.; Blassioli-Moraes, M.C. Development of Pull and Push-Pull Systems for Management of Lesser Mealworm, Alphitobius diaperinus, in Poultry Houses Using Alarm and Aggregation Pheromones. Pest Manag. Sci. 2019, 75, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Sobhy, I.S.; Caulfield, J.C.; Pickett, J.A.; Birkett, M.A. Sensing the Danger Signals: Cis-Jasmone Reduces Aphid Performance on Potato and Modulates the Magnitude of Released Volatiles. Front. Ecol. Evol. 2020, 7, 499. [Google Scholar] [CrossRef] [Green Version]
- Lee Díaz, A.S.; Rizaludin, M.S.; Zweers, H.; Raaijmakers, J.M.; Garbeva, P. Exploring the Volatiles Released from Roots of Wild and Domesticated Tomato Plants under Insect Attack. Molecules 2022, 27, 1612. [Google Scholar] [CrossRef] [PubMed]
- Schäffler, I.; Steiner, K.E.; Haid, M.; van Berkel, S.S.; Gerlach, G.; Johnson, S.D.; Wessjohann, L.; Dötterl, S. Diacetin, a Reliable Cue and Private Communication Channel in a Specialized Pollination System. Sci. Rep. 2015, 5, 12779. [Google Scholar] [CrossRef] [Green Version]
- Morawo, T.; Fadamiro, H. Identification of Key Plant-Associated Volatiles Emitted by Heliothis virescens Larvae That Attract the Parasitoid, Microplitis croceipes: Implications for Parasitoid Perception of Odor Blends. J. Chem. Ecol. 2016, 42, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.C.; Reinecke, A.; Gershenzon, J.; Unsicker, S.B. Feeding Experience Affects the Behavioral Response of Polyphagous Gypsy Moth Caterpillars to Herbivore-Induced Poplar Volatiles. J. Chem. Ecol. 2016, 42, 382–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kigathi, R.N.; Weisser, W.W.; Reichelt, M.; Gershenzon, J.; Unsicker, S.B. Plant Volatile Emission Depends on the Species Composition of the Neighboring Plant Community. BMC Plant Biol. 2019, 19, 58. [Google Scholar] [CrossRef] [PubMed]
- Michereff, M.F.F.; Magalhães, D.M.; Hassemer, M.J.; Laumann, R.A.; Zhou, J.-J.; Ribeiro, P.E.D.A.; Viana, P.A.; Guimarães, P.E.D.O.; Schimmelpfeng, P.H.C.; Borges, M.; et al. Variability in Herbivore-Induced Defence Signalling across Different Maize Genotypes Impacts Significantly on Natural Enemy Foraging Behaviour. J. Pest Sci. 2019, 92, 723–736. [Google Scholar] [CrossRef]
- Aartsma, Y.; Leroy, B.; van der Werf, W.; Dicke, M.; Poelman, E.H.; Bianchi, F.J.J.A. Intraspecific Variation in Herbivore-Induced Plant Volatiles Influences the Spatial Range of Plant-Parasitoid Interactions. Oikos 2019, 128, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Effah, E.; Holopainen, J.K.; McCormick, A.C. Potential Roles of Volatile Organic Compounds in Plant Competition. Perspect. Plant Ecol. Evol. Syst. 2019, 38, 58–63. [Google Scholar] [CrossRef]
- Hilker, M.; Fatouros, N.E. Plant Responses to Insect Egg Deposition. Annu. Rev. Entomol. 2015, 60, 493–515. [Google Scholar] [CrossRef] [PubMed]
- Turlings, T.C.J.; Erb, M. Tritrophic Interactions Mediated by Herbivore-Induced Plant Volatiles: Mechanisms, Ecological Relevance, and Application Potential. Annu. Rev. Entomol. 2018, 63, 433–452. [Google Scholar] [CrossRef] [PubMed]
- Mutyambai, D.M.; Bruce, T.J.A.; Midega, C.A.O.; Woodcock, C.M.; Caulfield, J.C.; Van Den Berg, J.; Pickett, J.A.; Khan, Z.R. Responses of Parasitoids to Volatiles Induced by Chilo partellus Oviposition on Teosinte, a Wild Ancestor of Maize. J. Chem. Ecol. 2015, 41, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.K.; Woods, H.A. Innate and Learned Olfactory Responses in a Wild Population of the Egg Parasitoid Trichogramma (Hymenoptera: Trichogrammatidae). J. Insect Sci. 2016, 16, 110. [Google Scholar] [CrossRef] [Green Version]
- Murali-Baskaran, R.K.; Mooventhan, P.; Das, D.; Dixit, A.; Sharma, K.C.; Senthil-Nathan, S.; Kaushal, P.; Ghosh, P.K. The Future of Plant Volatile Organic Compounds (PVOCs) Research: Advances and Applications for Sustainable Agriculture. Environ. Exp. Bot. 2022, 200, 104912. [Google Scholar] [CrossRef]
- Rusman, Q.; Lucas-Barbosa, D.; Poelman, E.H.; Dicke, M. Ecology of Plastic Flowers. Trends Plant Sci. 2019, 24, 725–740. [Google Scholar] [CrossRef] [PubMed]
- Rusman, Q.; Poelman, E.H.; Nowrin, F.; Polder, G.; Lucas-Barbosa, D. Floral Plasticity: Herbivore-species-specific-induced Changes in Flower Traits with Contrasting Effects on Pollinator Visitation. Plant Cell Environ. 2019, 42, 1882–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayelo, P.M.; Pirk, C.W.W.; Yusuf, A.A.; Chailleux, A.; Mohamed, S.A.; Deletre, E. Exploring the Kairomone-Based Foraging Behaviour of Natural Enemies to Enhance Biological Control: A Review. Front. Ecol. Evol. 2021, 9, 641974. [Google Scholar] [CrossRef]
- Silva, D.B.; Urbaneja, A.; Pérez-Hedo, M. Response of Mirid Predators to Synthetic Herbivore-induced Plant Volatiles. Entomol. Exp. Appl. 2021, 169, 125–132. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, X.; Zhan, Y.; Wang, K.; Francis, F.; Liu, Y. New Slow Release Mixture of (E)-β-farnesene with Methyl Salicylate to Enhance Aphid Biocontrol Efficacy in Wheat Ecosystem. Pest Manag. Sci. 2021, 77, 3341–3348. [Google Scholar] [CrossRef] [PubMed]
- Edwards, L.J.; Siddall, J.B.; Dunham, L.L.; Uden, P.; Kislow, C.J. Trans-β-Farnesene, Alarm Pheromone of the Green Peach Aphid, Myzus persicae (Sulzer). Nature 1973, 241, 126–127. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Gershenzon, J. The Chemical Diversity of Floral Scent. In Biology of Floral Scent; Pichersky, N.D.A.E., Ed.; Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 57–78. [Google Scholar]
- Salamanca, J.; Souza, B.; Kyryczenko-Roth, V.; Rodriguez-Saona, C. Methyl Salicylate Increases Attraction and Function of Beneficial Arthropods in Cranberries. Insects 2019, 10, 423. [Google Scholar] [CrossRef] [Green Version]
- Ding, P.; Ding, Y. Stories of Salicylic Acid: A Plant Defense Hormone. Trends Plant Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Uefune, M.; Abe, J.; Shiojiri, K.; Urano, S.; Nagasaka, K.; Takabayashi, J. Targeting Diamondback Moths in Greenhouses by Attracting Specific Native Parasitoids with Herbivory-Induced Plant Volatiles. R. Soc. Open Sci. 2020, 7, 201592. [Google Scholar] [CrossRef] [PubMed]
- Uefune, M.; Yoneya, K.; Yamamoto, M.; Takabayashi, J. The Use of Synthetic Herbivory-Induced Plant Volatiles That Attract Specialist Parasitoid Wasps, Cotesia vestalis, for Controlling the Incidence of Diamondback Moth Larvae in Open Agricultural Fields. Front. Ecol. Evol. 2021, 9, 702314. [Google Scholar] [CrossRef]
- Przybylska, A.; Ćwintal, M.; Pszczółkowski, P.; Sawicka, B. Effect of Attractants and Micronutrient Biofortification on the Yield and Quality of Red Clover (Trifolium pratense L.) Seeds. Agronomy 2021, 11, 152. [Google Scholar] [CrossRef]
- Cook, S.M.; Rasmussen, H.B.; Birkett, M.A.; Murray, D.A.; Pye, B.J.; Watts, N.P.; Williams, I.H. Behavioural and Chemical Ecology Underlying the Success of Turnip Rape (Brassica rapa) Trap Crops in Protecting Oilseed Rape (Brassica napus) from the Pollen Beetle (Meligethes aeneus). Arthropod-Plant Interact. 2007, 1, 57. [Google Scholar] [CrossRef]
- Theis, N.; Barber, N.A.; Gillespie, S.D.; Hazzard, R.V.; Adler, L.S. Attracting Mutualists and Antagonists: Plant Trait Variation Explains the Distribution of Specialist Floral Herbivores and Pollinators on Crops and Wild Gourds. Am. J. Bot. 2014, 101, 1314–1322. [Google Scholar] [CrossRef] [Green Version]
- Theis, N.; Adler, L.S. Advertising to the Enemy: Enhanced Floral Fragrance Increases Beetle Attraction and Reduces Plant Reproduction. Ecology 2012, 93, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Farina, W.M.; Arenas, A.; Díaz, P.C.; Susic Martin, C.; Corriale, M.J. In-Hive Learning of Specific Mimic Odours as a Tool to Enhance Honey Bee Foraging and Pollination Activities in Pear and Apple Crops. Sci. Rep. 2022, 12, 20510. [Google Scholar] [CrossRef] [PubMed]
- Ali, J.; Covaci, A.D.; Roberts, J.M.; Sobhy, I.S.; Kirk, W.D.J.; Bruce, T.J.A. Effects of Cis-Jasmone Treatment of Brassicas on Interactions With Myzus persicae Aphids and Their Parasitoid Diaeretiella Rapae. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Birkett, M.A.; Campbell, C.A.M.; Chamberlain, K.; Guerrieri, E.; Hick, A.J.; Martin, J.L.; Matthes, M.; Napier, J.A.; Pettersson, J.; Pickett, J.A.; et al. New Roles for Cis-Jasmone as an Insect Semiochemical and in Plant Defense. Proc. Natl. Acad. Sci. USA 2000, 97, 9329–9334. [Google Scholar] [CrossRef] [Green Version]
- Sobhy, I.S.; Woodcock, C.M.; Powers, S.J.; Caulfield, J.C.; Pickett, J.A.; Birkett, M.A. Cis-Jasmone Elicits Aphid-Induced Stress Signalling in Potatoes. J. Chem. Ecol. 2017, 43, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Moraes, M.C.B.; Laumann, R.A.; Pareja, M.; Sereno, F.T.P.S.; Michereff, M.F.F.; Birkett, M.A.; Pickett, J.A.; Borges, M. Attraction of the Stink Bug Egg Parasitoid Telenomus podisi to Defence Signals from Soybean Activated by Treatment with Cis-Jasmone. Entomol. Exp. Appl. 2009, 131, 178–188. [Google Scholar] [CrossRef]
- Hegde, M.; Oliveira, J.N.; da Costa, J.G.; Loza-Reyes, E.; Bleicher, E.; Santana, A.E.G.; Caulfield, J.C.; Mayon, P.; Dewhirst, S.Y.; Bruce, T.J.A.; et al. Aphid Antixenosis in Cotton Is Activated by the Natural Plant Defence Elicitor Cis-Jasmone. Phytochemistry 2012, 78, 81–88. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Matthes, M.C.; Chamberlain, K.; Woodcock, C.M.; Mohib, A.; Webster, B.; Smart, L.E.; Birkett, M.A.; Pickett, J.A.; Napier, J.A. Cis-Jasmone Induces Arabidopsis Genes That Affect the Chemical Ecology of Multitrophic Interactions with Aphids and Their Parasitoids. Proc. Natl. Acad. Sci. USA 2008, 105, 4553–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.H.; Gols, R.; Benrey, B. Crop Domestication and Its Impact on Naturally Selected Trophic Interactions. Annu. Rev. Entomol. 2015, 60, 35–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Jander, G. Molecular Ecology of Plant Volatiles in Interactions with Insect Herbivores. J. Exp. Bot. 2022, 73, 449–462. [Google Scholar] [CrossRef]
- Salvagnin, U.; Malnoy, M.; Thöming, G.; Tasin, M.; Carlin, S.; Martens, S.; Vrhovsek, U.; Angeli, S.; Anfora, G. Adjusting the Scent Ratio: Using Genetically Modified Vitis vinifera Plants to Manipulate European Grapevine Moth Behaviour. Plant Biotechnol. J. 2018, 16, 264–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ode, P.J. Caterpillars, Plant Chemistry, and Parasitoids in Natural vs. Agroecosystems. In Caterpillars in the Middle. Fascinating Life Sciences; Springer International Publishing: Cham, Switzerland, 2022; pp. 395–423. [Google Scholar] [CrossRef]
- Coll-Aráoz, M.V.; Hill, J.G.; Luft-Albarracin, E.; Virla, E.G.; Fernandez, P.C. Modern Maize Hybrids Have Lost Volatile Bottom-Up and Top-Down Control of Dalbulus maidis, a Specialist Herbivore. J. Chem. Ecol. 2020, 46, 906–915. [Google Scholar] [CrossRef] [PubMed]
- De Lange, E.S.; Laplanche, D.; Guo, H.; Xu, W.; Vlimant, M.; Erb, M.; Ton, J.; Turlings, T.C.J. Spodoptera frugiperda Caterpillars Suppress Herbivore-Induced Volatile Emissions in Maize. J. Chem. Ecol. 2020, 46, 344–360. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Su, S.; Liu, Q.; Jiao, Y.; Peng, Y.; Li, Y.; Turlings, T.C. Caterpillar-Induced Rice Volatiles Provide Enemy-Free Space for the Offspring of the Brown Planthopper. Elife 2020, 9, e55421. [Google Scholar] [CrossRef]
- Liu, Q.; Hu, X.; Su, S.; Ning, Y.; Peng, Y.; Ye, G.; Lou, Y.; Turlings, T.C.J.; Li, Y. Cooperative Herbivory between Two Important Pests of Rice. Nat. Commun. 2021, 12, 6772. [Google Scholar] [CrossRef]
- Sun, Y.L.; Dong, J.F.; Huang, L.Q.; Wang, C.Z. The Cotton Bollworm Endoparasitoid Campoletis chlorideae Is Attracted by Cis-Jasmone or Cis-3-Hexenyl Acetate but Not by Their Mixtures. Arthropod-Plant Interact. 2020, 14, 169–179. [Google Scholar] [CrossRef]
- Rincker, C.M.; Marble, V.L.; Brown, D.E.; Johansen, C.A. Seed Production Practices. In:Alfalfa and Alfalfa Improvement (Agronomy monogram 29); Hanson, A.A., Barnes, D.K., Hill, R.R., Jr., Eds.; ASA, CSSA, SSSA Publishers: Madison, WI, USA, 1988; pp. 985–1021. [Google Scholar]
- Henning, J.A.; Peng, Y.-S.; Montague, M.A.; Teuber, L.R. Honey Bee (Hymenoptera: Apidae) Behavioral Response to Primary Alfalfa (Rosales: Fabaceae) Floral Volatiles. J. Econ. Entomol. 1992, 85, 233–239. [Google Scholar] [CrossRef]
- Lukas, K.; Harig, T.; Schulz, S.; Hadersdorfer, J.; Dötterl, S. Flowers of European Pear Release Common and Uncommon Volatiles That Can Be Detected by Honey Bee Pollinators. Chemoecology 2019, 29, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Ma, W.; Zhang, Q.; Hu, X.; Ding, G.; Jiang, Y.; Huang, J. Honey Bee Foraging Decisions Influenced by Pear Volatiles. Agriculture 2022, 12, 1074. [Google Scholar] [CrossRef]
- Stasiak, A.; Latocha, P. Comparative Analysis of Volatile Compounds in Flowers of Different Actinidia Species. Plants 2020, 9, 1675. [Google Scholar] [CrossRef] [PubMed]
- Grajales-Conesa, J.; Meléndez Ramírez, V.; Cruz-López, L.; Sánchez Guillén, D. Effect of Citrus Floral Extracts on the Foraging Behavior of the Stingless Bee Scaptotrigona pectoralis (Dalla Torre). Rev. Bras. Entomol. 2012, 56, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Mas, F.; Harper, A.; Horner, R.; Welsh, T.; Jaksons, P.; Suckling, D.M. The Importance of Key Floral Bioactive Compounds to Honey Bees for the Detection and Attraction of Hybrid Vegetable Crops and Increased Seed Yield. J. Sci. Food Agric. 2018, 98, 4445–4453. [Google Scholar] [CrossRef]
- Kallure, G.S.; Kumari, A.; Shinde, B.A.; Giri, A.P. Characterized Constituents of Insect Herbivore Oral Secretions and Their Influence on the Regulation of Plant Defenses. Phytochemistry 2022, 193, 113008. [Google Scholar] [CrossRef] [PubMed]
- Helms, A.M.; De Moraes, C.M.; Tooker, J.F.; Mescher, M.C. Exposure of Solidago altissima Plants to Volatile Emissions of an Insect Antagonist (Eurosta solidaginis) Deters Subsequent Herbivory. Proc. Natl. Acad. Sci. USA 2013, 110, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helms, A.M.; De Moraes, C.M.; Mescher, M.C.; Tooker, J.F. The Volatile Emission of Eurosta solidaginis Primes Herbivore-Induced Volatile Production in Solidago altissima and Does Not Directly Deter Insect Feeding. BMC Plant Biol. 2014, 14, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helms, A.M.; De Moraes, C.M.; Tröger, A.; Alborn, H.T.; Francke, W.; Tooker, J.F.; Mescher, M.C. Identification of an Insect-Produced Olfactory Cue That Primes Plant Defenses. Nat. Commun. 2017, 8, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhães, D.M.; Da Silva, I.T.F.A.; Borges, M.; Laumann, R.A.; Blassioli-Moraes, M.C. Anthonomus grandis Aggregation Pheromone Induces Cotton Indirect Defence and Attracts the Parasitic Wasp Bracon vulgaris. J. Exp. Bot. 2019, 70, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial Volatile Emissions as Insect Semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef]
- Beck, J.J.; Vannette, R.L. Harnessing Insect–Microbe Chemical Communications To Control Insect Pests of Agricultural Systems. J. Agric. Food Chem. 2017, 65, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Hamby, K.A.; Becher, P.G. Current Knowledge of Interactions between Drosophila suzukii and Microbes, and Their Potential Utility for Pest Management. J. Pest Sci. 2016, 89, 621–630. [Google Scholar] [CrossRef]
- Bueno, E.; Martin, K.R.; Raguso, R.A.; Mcmullen, J.G.; Hesler, S.P.; Loeb, G.M.; Douglas, A.E. Response of Wild Spotted Wing Drosophila (Drosophila suzukii) to Microbial Volatiles. J. Chem. Ecol. 2020, 46, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Ljunggren, J.; Borrero-Echeverry, F.; Chakraborty, A.; Lindblom, T.U.T.; Hedenström, E.; Karlsson, M.; Witzgall, P.; Bengtsson, M. Yeast Volatomes Differentially Affect Larval Feeding in an Insect Herbivore. Appl. Environ. Microbiol. 2019, 85, e01761-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becher, P.G.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpędowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piškur, J. Chemical Signaling and Insect Attraction Is a Conserved Trait in Yeasts. Ecol. Evol. 2018, 8, 2962–2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baig, F.; Farnier, K.; Piper, A.M.; Speight, R.; Cunningham, J.P. Yeasts Influence Host Selection and Larval Fitness in Two Frugivorous Carpophilus Beetle Species. J. Chem. Ecol. 2020, 46, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Fountain, M.T.; Günther, C.S.; Eady, P.E.; Goddard, M.R. Separate and Combined Hanseniaspora uvarum and Metschnikowia pulcherrima Metabolic Volatiles Are Attractive to Drosophila suzukii in the Laboratory and Field. Sci. Rep. 2021, 11, 1201. [Google Scholar] [CrossRef]
- Davis, T.S.; Landolt, P.J. A Survey of Insect Assemblages Responding to Volatiles from a Ubiquitous Fungus in an Agricultural Landscape. J. Chem. Ecol. 2013, 39, 860–868. [Google Scholar] [CrossRef]
- Bego, A.; Burul, F.; Popović, M.; Jukić Špika, M.; Veršić Bratinčević, M.; Pošćić, F.; Vitanović, E. Bactrocera oleae (Rossi) (Diptera: Tephritidae) Response to Different Blends of Olive Fruit Fly-Associated Yeast Volatile Compounds as Attractants. Agronomy 2021, 12, 72. [Google Scholar] [CrossRef]
- Henneken, J.; Farnier, K.; Cunningham, J.P. A Synthetic Blend of Fruit and Live Yeast Odours Shows Promise for Trapping Mated Female Queensland Fruit Fly, Bactrocera tryoni, in the Field. J. Chem. Ecol. 2022, 48, 817–826. [Google Scholar] [CrossRef]
- Tasin, M.; Larsson Herrera, S.; Knight, A.L.; Barros-Parada, W.; Fuentes Contreras, E.; Pertot, I. Volatiles of Grape Inoculated with Microorganisms: Modulation of Grapevine Moth Oviposition and Field Attraction. Microb. Ecol. 2018, 76, 751–761. [Google Scholar] [CrossRef] [Green Version]
- Larsson Herrera, S.; Rikk, P.; Köblös, G.; Szelényi, M.O.; Molnár, B.P.; Dekker, T.; Tasin, M. Designing a Species-Selective Lure Based on Microbial Volatiles to Target Lobesia Botrana. Sci. Rep. 2020, 10, 6512. [Google Scholar] [CrossRef] [Green Version]
- Kendra, P.E.; Tabanca, N.; Cruz, L.F.; Menocal, O.; Schnell, E.Q.; Carrillo, D. Volatile Emissions and Relative Attraction of the Fungal Symbionts of Tea Shot Hole Borer (Coleoptera: Curculionidae). Biomolecules 2022, 12, 97. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Cruz, A.; Robledo, N.; Rosete-Enríquez, M.; Romero-López, A.A. Attraction of Adults of Cyclocephala lunulata and Cyclocephala barrerai (Coleoptera: Scarabaeoidea: Melolonthidae) towards Bacteria Volatiles Isolated from Their Genital Chambers. Molecules 2020, 25, 4430. [Google Scholar] [CrossRef] [PubMed]
- Goelen, T.; Sobhy, I.S.; Vanderaa, C.; Boer, J.G.; Delvigne, F.; Francis, F.; Wäckers, F.; Rediers, H.; Verstrepen, K.J.; Wenseleers, T.; et al. Volatiles of Bacteria Associated with Parasitoid Habitats Elicit Distinct Olfactory Responses in an Aphid Parasitoid and Its Hyperparasitoid. Funct. Ecol. 2020, 34, 507–520. [Google Scholar] [CrossRef]
- Goelen, T.; Sobhy, I.S.; Vanderaa, C.; Wäckers, F.; Rediers, H.; Wenseleers, T.; Jacquemyn, H.; Lievens, B. Bacterial Phylogeny Predicts Volatile Organic Compound Composition and Olfactory Response of an Aphid Parasitoid. Oikos 2020, 129, 1415–1428. [Google Scholar] [CrossRef]
- Goelen, T.; Vuts, J.; Sobhy, I.S.; Wäckers, F.; Caulfield, J.C.; Birkett, M.A.; Rediers, H.; Jacquemyn, H.; Lievens, B. Identification and Application of Bacterial Volatiles to Attract a Generalist Aphid Parasitoid: From Laboratory to Greenhouse Assays. Pest Manag. Sci. 2021, 77, 930–938. [Google Scholar] [CrossRef]
- Đurović, G.; Van Neerbos, F.A.C.; Bossaert, S.; Herrera-Malaver, B.; Steensels, J.; Arnó, J.; Wäckers, F.; Sobhy, I.S.; Verstrepen, K.J.; Jacquemyn, H.; et al. The Pupal Parasitoid Trichopria drosophilae Is Attracted to the Same Yeast Volatiles as Its Adult Host. J. Chem. Ecol. 2021, 47, 788–798. [Google Scholar] [CrossRef]
- van Neerbos, F.A.C.; Dewitte, P.; Wäckers, F.; Wenseleers, T.; Jacquemyn, H.; Lievens, B. Bacterial Volatiles Elicit Differential Olfactory Responses in Insect Species from the Same and Different Trophic Levels. Insect Sci. 2023, 1–17. [Google Scholar] [CrossRef]
- Cusumano, A.; Bella, P.; Peri, E.; Rostás, M.; Guarino, S.; Lievens, B.; Colazza, S. Nectar-Inhabiting Bacteria Affect Olfactory Responses of an Insect Parasitoid by Altering Nectar Odors. Microb. Ecol. 2022, 1–13. [Google Scholar] [CrossRef]
- Kandasamy, D.; Gershenzon, J.; Andersson, M.N.; Hammerbacher, A. Volatile Organic Compounds Influence the Interaction of the Eurasian Spruce Bark Beetle (Ips typographus) with Its Fungal Symbionts. ISME J. 2019, 13, 1788–1800. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, D.; Zaman, R.; Nakamura, Y.; Zhao, T.; Hartmann, H.; Andersson, M.N.; Hammerbacher, A.; Gershenzon, J. Bark Beetles Locate Fungal Symbionts by Detecting Volatile Fungal Metabolites of Host Tree Resin Monoterpenes. bioRxiv 2021. [Google Scholar] [CrossRef]
- Morrison, W.R.; Scully, E.D.; Campbell, J.F. Towards Developing Areawide Semiochemical-mediated, Behaviorally-based Integrated Pest Management Programs for Stored Product Insects. Pest Manag. Sci. 2021, 77, 2667–2682. [Google Scholar] [CrossRef] [PubMed]
- Ponce, M.A.; Kim, T.N.; Morrison, W. R, III. A Systematic Review of the Behavioral Responses by Stored-Product Arthropods to Individual or Blends of Microbially Produced Volatile Cues. Insects 2021, 12, 391. [Google Scholar] [CrossRef] [PubMed]
- Van Winkle, T.; Ponce, M.; Quellhorst, H.; Bruce, A.; Albin, C.E.; Kim, T.N.; Zhu, K.Y.; Morrison, W.R. Microbial Volatile Organic Compounds from Tempered and Incubated Grain Mediate Attraction by a Primary but Not Secondary Stored Product Insect Pest in Wheat. J. Chem. Ecol. 2022, 48, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Scheff, D.S.; Campbell, J.F.; Arthur, F.H. Seasonal, Landscape, and Attractant Effects on Lesser Grain Borer, Rhyzopertha dominica (F.), Captures in Northeast Kansas. Agronomy 2021, 12, 99. [Google Scholar] [CrossRef]
- Jia, F.; Toews, M.D.; Campbell, J.F.; Ramaswamy, S.B. Survival and Reproduction of Lesser Grain Borer, Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae) on Flora Associated with Native Habitats in Kansas. J. Stored Prod. Res. 2008, 44, 366–372. [Google Scholar] [CrossRef]
- Campos, M.; Phillips, T.W. Attract-and-Kill and Other Pheromone-Based Methods to Suppress Populations of the Indianmeal Moth (Lepidoptera: Pyralidae). J. Econ. Entomol. 2014, 107, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, R.V.; Campbell, J.F.; Zhu, K.Y.; Starkus, L.A.; McKay, T.; Morrison, W.R. Long-Lasting Insecticide-Incorporated Netting and Interception Traps at Pilot-Scale Warehouses and Commercial Facilities Prevents Infestation by Stored Product Beetles. Front. Sustain. Food Syst. 2021, 4, 561820. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical Diversity of Microbial Volatiles and Their Potential for Plant Growth and Productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-Kingdom Interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.; Withall, D.; Birkett, M. Harnessing Microbial Volatiles to Replace Pesticides and Fertilizers. Microb. Biotechnol. 2020, 13, 1366–1376. [Google Scholar] [CrossRef]
- Garbeva, P.; Weisskopf, L. Airborne Medicine: Bacterial Volatiles and Their Influence on Plant Health. New Phytol. 2020, 226, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Weisskopf, L.; Schulz, S.; Garbeva, P. Microbial Volatile Organic Compounds in Intra-Kingdom and Inter-Kingdom Interactions. Nat. Rev. Microbiol. 2021, 19, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Paré, P.W.; Kloepper, J.W. Bacterial Volatiles Promote Growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [Green Version]
- Lammers, A.; Lalk, M.; Garbeva, P. Air Ambulance: Antimicrobial Power of Bacterial Volatiles. Antibiotics 2022, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Kloepper, J.W.; Paré, P.W. Bacterial Volatiles Induce Systemic Resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [Green Version]
- Song, G.C.; Riu, M.; Ryu, C.-M. Beyond the Two Compartments Petri-Dish: Optimising Growth Promotion and Induced Resistance in Cucumber Exposed to Gaseous Bacterial Volatiles in a Miniature Greenhouse System. Plant Methods 2019, 15, 9. [Google Scholar] [CrossRef]
- Kong, H.G.; Shin, T.S.; Kim, T.H.; Ryu, C.-M. Stereoisomers of the Bacterial Volatile Compound 2,3-Butanediol Differently Elicit Systemic Defense Responses of Pepper against Multiple Viruses in the Field. Front. Plant Sci. 2018, 9, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.Y.; Seo, S.Y.; Oh, B.-R.; Seo, J.-W.; Kim, Y.J. 2,3-Butanediol Induces Systemic Acquired Resistance in the Plant Immune Response. J. Plant Biol. 2018, 61, 424–434. [Google Scholar] [CrossRef]
- Han, S.H.; Lee, S.J.; Moon, J.H.; Park, K.H.; Yang, K.Y.; Cho, B.H.; Kim, K.Y.; Kim, Y.W.; Lee, M.C.; Anderson, A.J.; et al. GacS-Dependent Production of 2R, 3R-Butanediol by Pseudomonas chlororaphis O6 Is a Major Determinant for Eliciting Systemic Resistance Against Erwinia carotovora but Not Against Pseudomonas syringae Pv. Tabaci in Tobacco. Mol. Plant-Microbe Interact. 2006, 19, 924–930. [Google Scholar] [CrossRef] [Green Version]
- Song, G.C.; Choi, H.K.; Ryu, C.-M. Gaseous 3-Pentanol Primes Plant Immunity against a Bacterial Speck Pathogen, Pseudomonas syringae Pv. Tomato via Salicylic Acid and Jasmonic Acid-Dependent Signaling Pathways in Arabidopsis. Front. Plant Sci. 2015, 6, 821. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.K.; Song, G.C.; Yi, H.-S.; Ryu, C.-M. Field Evaluation of the Bacterial Volatile Derivative 3-Pentanol in Priming for Induced Resistance in Pepper. J. Chem. Ecol. 2014, 40, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Farag, M.A.; Park, H.B.; Kloepper, J.W.; Lee, S.H.; Ryu, C.-M. Induced Resistance by a Long-Chain Bacterial Volatile: Elicitation of Plant Systemic Defense by a C13 Volatile Produced by Paenibacillus polymyxa. PLoS ONE 2012, 7, e48744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riu, M.; Kim, M.S.; Choi, S.-K.; Oh, S.-K.; Ryu, C.-M. Elicitation of Innate Immunity by a Bacterial Volatile 2-Nonanone at Levels below Detection Limit in Tomato Rhizosphere. Mol. Cells 2022, 45, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Kottb, M.; Gigolashvili, T.; Großkinsky, D.K.; Piechulla, B. Trichoderma Volatiles Effecting Arabidopsis: From Inhibition to Protection against Phytopathogenic Fungi. Front. Microbiol. 2015, 6, 995. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Cornejo, H.A.; López-Bucio, J.S.; Méndez-Bravo, A.; Macías-Rodríguez, L.; Ramos-Vega, M.; Guevara-García, Á.A.; López-Bucio, J. Mitogen-Activated Protein Kinase 6 and Ethylene and Auxin Signaling Pathways Are Involved in Arabidopsis Root-System Architecture Alterations by Trichoderma atroviride. Mol. Plant-Microbe Interact. 2015, 28, 701–710. [Google Scholar] [CrossRef] [Green Version]
- da Silva, L.R.; Valadares-Inglis, M.C.; Peixoto, G.H.S.; de Luccas, B.E.G.; Muniz, P.H.P.C.; Magalhães, D.M.; Moraes, M.C.B.; de Mello, S.C.M. Volatile Organic Compounds Emitted by Trichoderma azevedoi Promote the Growth of Lettuce Plants and Delay the Symptoms of White Mold. Biol. Control 2021, 152, 104447. [Google Scholar] [CrossRef]
- Naznin, H.A.; Kiyohara, D.; Kimura, M.; Miyazawa, M.; Shimizu, M.; Hyakumachi, M. Systemic Resistance Induced by Volatile Organic Compounds Emitted by Plant Growth-Promoting Fungi in Arabidopsis thaliana. PLoS ONE 2014, 9, e86882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagiwa, Y.; Inagaki, Y.; Ichinose, Y.; Toyoda, K.; Hyakumachi, M.; Shiraishi, T. Talaromyces wortmannii FS2 Emits β-Caryophyllene, Which Promotes Plant Growth and Induces Resistance. J. Gen. Plant Pathol. 2011, 77, 336–341. [Google Scholar] [CrossRef]
- Song, G.C.; Im, H.; Jung, J.; Lee, S.; Jung, M.; Rhee, S.; Ryu, C. Plant Growth-promoting Archaea Trigger Induced Systemic Resistance in Arabidopsis thaliana against Pectobacterium carotovorum and Pseudomonas syringae. Environ. Microbiol. 2019, 21, 940–948. [Google Scholar] [CrossRef]
- Song, G.; Ryu, C.-M. Two Volatile Organic Compounds Trigger Plant Self-Defense against a Bacterial Pathogen and a Sucking Insect in Cucumber under Open Field Conditions. Int. J. Mol. Sci. 2013, 14, 9803–9819. [Google Scholar] [CrossRef] [Green Version]
- Sharifi, R.; Lee, S.-M.; Ryu, C.-M. Microbe-Induced Plant Volatiles. New Phytol. 2018, 220, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Song, G.C.; Jeon, J.; Choi, H.K.; Sim, H.; Kim, S.; Ryu, C. Bacterial Type III Effector–Induced Plant C8 Volatiles Elicit Antibacterial Immunity in Heterospecific Neighbouring Plants via Airborne Signalling. Plant Cell Environ. 2022, 45, 236–247. [Google Scholar] [CrossRef]
- Kong, H.G.; Song, G.C.; Sim, H.-J.; Ryu, C.-M. Achieving Similar Root Microbiota Composition in Neighbouring Plants through Airborne Signalling. ISME J. 2021, 15, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Laupheimer, S.; Kurzweil, L.; Proels, R.; Unsicker, S.B.; Stark, T.D.; Dawid, C.; Hückelhoven, R. Volatile-mediated Signalling in Barley Induces Metabolic Reprogramming and Resistance against the Biotrophic Fungus Blumeria hordei. Plant Biol. 2023, 25, 72–84. [Google Scholar] [CrossRef]
- Xu, Y.; Tong, Z.; Zhang, X.; Zhang, X.; Luo, Z.; Shao, W.; Li, L.; Ma, Q.; Zheng, X.; Fang, W. Plant Volatile Organic Compound (E)-2-hexenal Facilitates Botrytis cinerea Infection of Fruits by Inducing Sulfate Assimilation. New Phytol. 2021, 231, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Linder, T. Assimilation of Alternative Sulfur Sources in Fungi. World J. Microbiol. Biotechnol. 2018, 34, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, X.; Granjel, R.R.; Fuente, M.; Fernández-Conradi, P.; Pasch, V.; Soengas, P.; Turlings, T.C.J.; Vázquez-González, C.; Abdala-Roberts, L.; Rasmann, S. Apparent Inhibition of Induced Plant Volatiles by a Fungal Pathogen Prevents Airborne Communication between Potato Plants. Plant Cell Environ. 2021, 44, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.J.; Alborn, H.T.; Block, A.K.; Christensen, S.A.; Hunter, C.T.; Rering, C.C.; Seidl-Adams, I.; Stuhl, C.J.; Torto, B.; Tumlinson, J.H. Interactions Among Plants, Insects, and Microbes: Elucidation of Inter-Organismal Chemical Communications in Agricultural Ecology. J. Agric. Food Chem. 2018, 66, 6663–6674. [Google Scholar] [CrossRef]
- Simon, J.-C.; Biere, A.; Sugio, A. The Promises and Challenges of Research on Plant-Insect-Microbe Interactions. Insect Sci. 2017, 24, 904–909. [Google Scholar] [CrossRef]
- Wilberts, L.; Vuts, J.; Caulfield, J.C.; Thomas, G.; Birkett, M.A.; Herrera-Malaver, B.; Verstrepen, K.J.; Sobhy, I.S.; Jacquemyn, H.; Lievens, B. Impact of Endophytic Colonization by Entomopathogenic Fungi on the Behavior and Life History of the Tobacco Peach Aphid Myzus persicae Var. Nicotianae. PLoS ONE 2022, 17, e0273791. [Google Scholar] [CrossRef]
- González-Mas, N.; Sánchez-Ortiz, A.; Valverde-García, P.; Quesada-Moraga, E. Effects of Endophytic Entomopathogenic Ascomycetes on the Life-History Traits of Aphis gossypii Glover and Its Interactions with Melon Plants. Insects 2019, 10, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, E.; Woestmann, L.; Biere, A.; Saastamoinen, M. A Plant Pathogen Modulates the Effects of Secondary Metabolites on the Performance and Immune Function of an Insect Herbivore. Oikos 2018, 127, 1539–1549. [Google Scholar] [CrossRef] [Green Version]
- Peñaflor, M.F.G.V.; Bento, J.M.S. Red-Rot Infection in Sugarcane Attenuates the Attractiveness of Sugarcane Borer-Induced Plant Volatiles to Parasitoid. Arthropod-Plant Interact. 2019, 13, 117–125. [Google Scholar] [CrossRef]
- Zehnder, G.; Kloepper, J.; Tuzun, S.; Yao, C.; Wei, G.; Chambliss, O.; Shelby, R. Insect Feeding on Cucumber Mediated by Rhizobacteria-Induced Plant Resistance. Entomol. Exp. Appl. 1997, 83, 81–85. [Google Scholar] [CrossRef]
- Van Oosten, V.R.; Bodenhausen, N.; Reymond, P.; Van Pelt, J.A.; Van Loon, L.C.; Dicke, M.; Pieterse, C.M.J. Differential Effectiveness of Microbially Induced Resistance Against Herbivorous Insects in Arabidopsis. Mol. Plant-Microbe Interact. 2008, 21, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela-Soto, J.H.; Estrada-Hernández, M.G.; Ibarra-Laclette, E.; Délano-Frier, J.P. Inoculation of Tomato Plants (Solanum lycopersicum) with Growth-Promoting Bacillus Subtilis Retards Whitefly Bemisia tabaci Development. Planta 2010, 231, 397–410. [Google Scholar] [CrossRef]
- Zebelo, S.; Song, Y.; Kloepper, J.W.; Fadamiro, H. Rhizobacteria Activates (+)-δ-Cadinene Synthase Genes and Induces Systemic Resistance in Cotton against Beet Armyworm (Spodoptera exigua). Plant Cell Environ. 2016, 39, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Gadhave, K.R.; Gange, A.C. Plant-Associated Bacillus Spp. Alter Life-History Traits of the Specialist Insect Brevicoryne brassicae L. Agric. For. Entomol. 2016, 18, 35–42. [Google Scholar] [CrossRef]
- Disi, J.O.; Mohammad, H.K.; Lawrence, K.; Kloepper, J.; Fadamiro, H. A Soil Bacterium Can Shape Belowground Interactions between Maize, Herbivores and Entomopathogenic Nematodes. Plant Soil 2019, 437, 83–92. [Google Scholar] [CrossRef]
- Ling, S.; Zhao, Y.; Sun, S.; Zheng, D.; Sun, X.; Zeng, R.; Chen, D.; Song, Y. Enhanced Anti-Herbivore Defense of Tomato Plants against Spodoptera litura by Their Rhizosphere Bacteria. BMC Plant Biol. 2022, 22, 254. [Google Scholar] [CrossRef]
- Pineda, A.; Soler, R.; Weldegergis, B.T.; Shimwela, M.M.; Van Loon, J.J.A.; Dicke, M. Non-Pathogenic Rhizobacteria Interfere with the Attraction of Parasitoids to Aphid-Induced Plant Volatiles via Jasmonic Acid Signalling. Plant Cell Environ. 2013, 36, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Moisan, K.; Dicke, M.; Raaijmakers, J.M.; Rachmawati, E.; Cordovez, V. Volatiles from the Fungus Fusarium oxysporum Affect Interactions of Brassica rapa Plants with Root Herbivores. Ecol. Entomol. 2021, 46, 240–248. [Google Scholar] [CrossRef]
- Moisan, K.; Cordovez, V.; van de Zande, E.M.; Raaijmakers, J.M.; Dicke, M.; Lucas-Barbosa, D. Volatiles of Pathogenic and Non-Pathogenic Soil-Borne Fungi Affect Plant Development and Resistance to Insects. Oecologia 2019, 190, 589–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrieri, E.; Lingua, G.; Digilio, M.C.; Massa, N.; Berta, G. Do Interactions between Plant Roots and the Rhizosphere Affect Parasitoid Behaviour? Ecol. Entomol. 2004, 29, 753–756. [Google Scholar] [CrossRef]
- Battaglia, D.; Bossi, S.; Cascone, P.; Digilio, M.C.; Prieto, J.D.; Fanti, P.; Guerrieri, E.; Iodice, L.; Lingua, G.; Lorito, M.; et al. Tomato Below Ground–Above Ground Interactions: Trichoderma longibrachiatum Affects the Performance of Macrosiphum euphorbiae and Its Natural Antagonists. Mol. Plant-Microbe Interact. 2013, 26, 1249–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngumbi, E. Mechanisms of Olfaction in Parasitic Wasps: Analytical and Behavioral Studies of Response of A Specialist (Microplitis croceipes) and a Generalist (Cotesia marginiventris) Parasitoid to Host-Related Odor; Auburn University: Auburn, AL, USA, 2011. [Google Scholar]
- Nangle, K. Effects of Plant Growth-Promoting Rhizobacteria (PGPR) Treatment of Cotton on the Oviposition Behavior of Spodoptera exigua (Hübner) (Lepidoptera: noctuidae); Auburn University: Auburn, AL, USA, 2012. [Google Scholar]
- Pulido, H.; Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Combined Effects of Mutualistic Rhizobacteria Counteract Virus-Induced Suppression of Indirect Plant Defences in Soya Bean. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190211. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.E.; Meek, H.C. The Influence of Arbuscular Mycorrhizal Fungi on Plant Reproduction. J. Chem. Ecol. 2020, 46, 707–721. [Google Scholar] [CrossRef]
- Moore, J.A.M.; Abraham, P.E.; Michener, J.K.; Muchero, W.; Cregger, M.A. Ecosystem Consequences of Introducing Plant Growth Promoting Rhizobacteria to Managed Systems and Potential Legacy Effects. New Phytol. 2022, 234, 1914–1918. [Google Scholar] [CrossRef]
- Becklin, K.M.; Gamez, G.; Uelk, B.; Raguso, R.A.; Galen, C. Soil Fungal Effects on Floral Signals, Rewards, and Aboveground Interactions in an Alpine Pollination Web. Am. J. Bot. 2011, 98, 1299–1308. [Google Scholar] [CrossRef] [Green Version]
- Groen, S.C.; Jiang, S.; Murphy, A.M.; Cunniffe, N.J.; Westwood, J.H.; Davey, M.P.; Bruce, T.J.A.; Caulfield, J.C.; Furzer, O.J.; Reed, A.; et al. Virus Infection of Plants Alters Pollinator Preference: A Payback for Susceptible Hosts? PLoS Pathog. 2016, 12, e1005790. [Google Scholar] [CrossRef] [Green Version]
- Vannette, R.L. The Floral Microbiome: Plant, Pollinator, and Microbial Perspectives. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 363–386. [Google Scholar] [CrossRef]
- Sobhy, I.S.; Baets, D.; Goelen, T.; Herrera-Malaver, B.; Bosmans, L.; Van den Ende, W.; Verstrepen, K.J.; Wäckers, F.; Jacquemyn, H.; Lievens, B. Sweet Scents: Nectar Specialist Yeasts Enhance Nectar Attraction of a Generalist Aphid Parasitoid Without Affecting Survival. Front. Plant Sci. 2018, 9, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffer, R.N.; Mei, Y.Z.; Andicoechea, J.; Manson, J.S.; Irwin, R.E. Consequences of a Nectar Yeast for Pollinator Preference and Performance. Funct. Ecol. 2017, 31, 613–621. [Google Scholar] [CrossRef]
- Schaeffer, R.N.; Rering, C.C.; Maalouf, I.; Beck, J.J.; Vannette, R.L. Microbial Metabolites Elicit Distinct Olfactory and Gustatory Preferences in Bumblebees. Biol. Lett. 2019, 15, 20190132. [Google Scholar] [CrossRef]
- Rering, C.C.; Beck, J.J.; Hall, G.W.; McCartney, M.M.; Vannette, R.L. Nectar-inhabiting Microorganisms Influence Nectar Volatile Composition and Attractiveness to a Generalist Pollinator. New Phytol. 2018, 220, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helletsgruber, C.; Dötterl, S.; Ruprecht, U.; Junker, R.R. Epiphytic Bacteria Alter Floral Scent Emissions. J. Chem. Ecol. 2017, 43, 1073–1077. [Google Scholar] [CrossRef] [Green Version]
- Rering, C.C.; Vannette, R.L.; Schaeffer, R.N.; Beck, J.J. Microbial Co-Occurrence in Floral Nectar Affects Metabolites and Attractiveness to a Generalist Pollinator. J. Chem. Ecol. 2020, 46, 659–667. [Google Scholar] [CrossRef]
- Giron, D.; Dedeine, F.; Dubreuil, G.; Huguet, E.; Mouton, L.; Outreman, Y.; Vavre, F.; Simon, J.-C. Influence of Microbial Symbionts on Plant–Insect Interactions. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2017; Volume 81, pp. 225–257. ISBN 978-0-12-803318-0. [Google Scholar]
- Wamonje, F.O.; Tungadi, T.D.; Murphy, A.M.; Pate, A.E.; Woodcock, C.; Caulfield, J.C.; Mutuku, J.M.; Cunniffe, N.J.; Bruce, T.J.A.; Gilligan, C.A.; et al. Three Aphid-Transmitted Viruses Encourage Vector Migration From Infected Common Bean (Phaseolus vulgaris) Plants Through a Combination of Volatile and Surface Cues. Front. Plant Sci. 2020, 11, 613772. [Google Scholar] [CrossRef]
- Arinaitwe, W.; Guyon, A.; Tungadi, T.D.; Cunniffe, N.J.; Rhee, S.-J.; Khalaf, A.; Mhlanga, N.M.; Pate, A.E.; Murphy, A.M.; Carr, J.P. The Effects of Cucumber Mosaic Virus and Its 2a and 2b Proteins on Interactions of Tomato Plants with the Aphid Vectors Myzus persicae and Macrosiphum euphorbiae. Viruses 2022, 14, 1703. [Google Scholar] [CrossRef]
- Franco, F.P.; Túler, A.C.; Gallan, D.Z.; Gonçalves, F.G.; Favaris, A.P.; Peñaflor, M.F.G.V.; Leal, W.S.; Moura, D.S.; Bento, J.M.S.; Silva-Filho, M.C. Fungal Phytopathogen Modulates Plant and Insect Responses to Promote Its Dissemination. ISME J. 2021, 15, 3522–3533. [Google Scholar] [CrossRef]
- Rodriguez-Saona, C.; Kaplan, I.; Braasch, J.; Chinnasamy, D.; Williams, L. Field Responses of Predaceous Arthropods to Methyl Salicylate: A Meta-Analysis and Case Study in Cranberries. Biol. Control 2011, 59, 294–303. [Google Scholar] [CrossRef]
- Benelli, G.; Lucchi, A. From Insect Pheromones to Mating Disruption: Theory and Practice. Insects 2021, 12, 698. [Google Scholar] [CrossRef] [PubMed]
- Arnault, I.; Fleurance, C.; Vey, F.; Du Fretay, G.; Auger, J. Use of Alliaceae Residues to Control Soil-Borne Pathogens. Ind. Crop. Prod. 2013, 49, 265–272. [Google Scholar] [CrossRef]
- Gómez-Tenorio, M.A.; Zanón, M.J.; de Cara, M.; Lupión, B.; Tello, J.C. Efficacy of Dimethyl Disulfide (DMDS) against Meloidogyne Sp. and Three Formae Speciales of Fusarium oxysporum under Controlled Conditions. Crop. Prot. 2015, 78, 263–269. [Google Scholar] [CrossRef]
- Papazlatani, C.; Rousidou, C.; Katsoula, A.; Kolyvas, M.; Genitsaris, S.; Papadopoulou, K.K.; Karpouzas, D.G. Assessment of the Impact of the Fumigant Dimethyl Disulfide on the Dynamics of Major Fungal Plant Pathogens in Greenhouse Soils. Eur. J. Plant Pathol. 2016, 146, 391–400. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Tabanca, N.; Owens, D.; Epsky, N.D. Utility of Essential Oils for Development of Host-Based Lures for Xyleborus glabratus (Coleoptera: Curculionidae: Scolytinae), Vector of Laurel Wilt. Open Chem. 2018, 16, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Schnell, E.Q.; Deyrup, M.A.; Epsky, N.D. Evaluation of Seven Essential Oils Identifies Cubeb Oil as Most Effective Attractant for Detection of Xyleborus glabratus. J. Pest Sci. 2014, 87, 681–689. [Google Scholar] [CrossRef]
- Cloonan, K.R.; Montgomery, W.S.; Narvaez, T.I.; Carrillo, D.; Kendra, P.E. Community of Bark and Ambrosia Beetles (Coleoptera: Curculionidae: Scolytinae and Platypodinae) in Agricultural and Forest Ecosystems with Laurel Wilt. Insects 2022, 13, 971. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Deyrup, M.A.; Wakarchuk, D. Improved Lure for Redbay Ambrosia Beetle Developed by Enrichment of α-Copaene Content. J. Pest Sci. 2016, 89, 427–438. [Google Scholar] [CrossRef]
- Kendra, P.E.; Owens, D.; Montgomery, W.S.; Narvaez, T.I.; Bauchan, G.R.; Schnell, E.Q.; Tabanca, N.; Carrillo, D. α-Copaene Is an Attractant, Synergistic with Quercivorol, for Improved Detection of Euwallacea nr. fornicatus (Coleoptera: Curculionidae: Scolytinae). PLoS ONE 2017, 12, e0179416. [Google Scholar] [CrossRef]
- Owens, D.; Kendra, P.E.; Tabanca, N.; Narvaez, T.I.; Montgomery, W.S.; Schnell, E.Q.; Carrillo, D. Quantitative Analysis of Contents and Volatile Emissions from α-Copaene and Quercivorol Lures, and Longevity for Attraction of Euwallacea nr. fornicatus in Florida. J. Pest Sci. 2019, 92, 237–252. [Google Scholar] [CrossRef]
- Malerbo-Souza, D.T.; Nogueira-Couto, R.H.; Couto, L.A. Honey Bee Attractants and Pollination in Sweet Orange, Citrus sinensis (L.) Osbeck, Var. Pera-Rio. J. Venom. Anim. Toxins Incl. Trop. Dis. 2004, 10, 144–153. [Google Scholar] [CrossRef]
- Ellis, A.; Delaplane, K.S. An Evaluation of Fruit-BoostTM as an Aid for Honey Bee Pollination under Conditions of Competing Bloom. J. Apic. Res. 2009, 48, 15–18. [Google Scholar] [CrossRef]
- Williamson, J.; Adams, C.G.; Isaacs, R.; Gut, L.J. Evaluation of Nasonov Pheromone Dispensers for Pollinator Attraction in Apple, Blueberry, and Cherry. J. Econ. Entomol. 2018, 111, 1658–1663. [Google Scholar] [CrossRef] [PubMed]
- Jailyang, L.; Sharma, N.C.; Chandel, J.S.; Rana, V.S.; Rana, K.; Chauhan, P. Influence of Bee Scent and Other Indigenous Bee Attractants on Bee Activity and Fruiting Behaviour of Kiwifruit (Actinidia deliciosa A. Chev.). Sci. Hortic. 2022, 295, 110869. [Google Scholar] [CrossRef]
- Free, J.B.; Ferguson, A.W.; Simpkins, J.R.; Al-Sa’ad, B.N. Effect of Honeybee Nasonov and Alarm Pheromone Components on Behaviour at the Nest Entrance. J. Apic. Res. 1983, 22, 214–223. [Google Scholar] [CrossRef]
- Schmidt, J.O. Attraction of Reproductive Honey Bee Swarms to Artificial Nests by Nasonov Pheromone. J. Chem. Ecol. 1994, 20, 1053–1056. [Google Scholar] [CrossRef] [PubMed]
- Pernal, S.F. The Social Life of Honey Bees. Vet. Clin. N. Am. Food Anim. Pract. 2021, 37, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.G.; Calam, D.H. Pheromones of the Honey Bee—The Secretion of the Nassanoff Gland of the Worker. J. Insect Physiol. 1969, 15, 237–244. [Google Scholar] [CrossRef]
- Pickett, J.A.; Woodcock, C.M.; Midega, C.A.; Khan, Z.R. Push–Pull Farming Systems. Curr. Opin. Biotechnol. 2014, 26, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, Z.; Midega, C.A.O.; Hooper, A.; Pickett, J. Push-Pull: Chemical Ecology-Based Integrated Pest Management Technology. J. Chem. Ecol. 2016, 42, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Sobhy, I.S.; Tamiru, A.; Chiriboga Morales, X.; Nyagol, D.; Cheruiyot, D.; Chidawanyika, F.; Subramanian, S.; Midega, C.A.O.; Bruce, T.J.A.; Khan, Z.R. Bioactive Volatiles From Push-Pull Companion Crops Repel Fall Armyworm and Attract Its Parasitoids. Front. Ecol. Evol. 2022, 10, 883020. [Google Scholar] [CrossRef]
- Mauchline, A.L.; Hervé, M.R.; Cook, S.M. Semiochemical-Based Alternatives to Synthetic Toxicant Insecticides for Pollen Beetle Management. Arthropod-Plant Interact. 2018, 12, 835–847. [Google Scholar] [CrossRef] [Green Version]
- Blassioli-Moraes, M.C.; Venzon, M.; Silveira, L.C.P.; Gontijo, L.M.; Togni, P.H.B.; Sujii, E.R.; Haro, M.M.; Borges, M.; Michereff, M.F.F.; de Aquino, M.F.S.; et al. Companion and Smart Plants: Scientific Background to Promote Conservation Biological Control. Neotrop Entomol. 2022, 51, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Togni, P.H.B.; Laumann, R.A.; Medeiros, M.A.; Sujii, E.R. Odour Masking of Tomato Volatiles by Coriander Volatiles in Host Plant Selection of Bemisia tabaci Biotype B. Entomol. Exp. Appl. 2010, 136, 164–173. [Google Scholar] [CrossRef]
- Pouët, C.; Deletre, E.; Rhino, B. Repellency of Wild Oregano Plant Volatiles, Plectranthus amboinicus, and Their Essential Oils to the Silverleaf Whitefly, Bemisia tabaci, on Tomato. Neotrop Entomol. 2022, 51, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lu, X.; Zhang, Z.; Huang, J.; Zhang, J.; Wang, L.; Hafeez, M.; Fernández-Grandon, G.M.; Lu, Y. Intercropping Rosemary (Rosmarinus officinalis) with Sweet Pepper (Capsicum annum) Reduces Major Pest Population Densities without Impacting Natural Enemy Populations. Insects 2021, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.; Lortie, C.J. Finding the Bees Knees: A Conceptual Framework and Systematic Review of the Mechanisms of Pollinator-Mediated Facilitation. Perspect. Plant Ecol. Evol. Syst. 2019, 36, 33–40. [Google Scholar] [CrossRef]
- Gilpin, A.-M.; Kobel, C.; Brettell, L.E.; O’Brien, C.; Cook, J.M.; Power, S.A. Co-Flowering Species Richness Increases Pollinator Visitation to Apple Flowers. Agriculture 2022, 12, 1246. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Aradottir, G.I.; Smart, L.E.; Martin, J.L.; Caulfield, J.C.; Doherty, A.; Sparks, C.A.; Woodcock, C.M.; Birkett, M.A.; Napier, J.A.; et al. The First Crop Plant Genetically Engineered to Release an Insect Pheromone for Defence. Sci. Rep. 2015, 5, 11183. [Google Scholar] [CrossRef] [Green Version]
- Beale, M.H.; Birkett, M.A.; Bruce, T.J.A.; Chamberlain, K.; Field, L.M.; Huttly, A.K.; Martin, J.L.; Parker, R.; Phillips, A.L.; Pickett, J.A.; et al. Aphid Alarm Pheromone Produced by Transgenic Plants Affects Aphid and Parasitoid Behavior. Proc. Natl. Acad. Sci. USA 2006, 103, 10509–10513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-L.; Ding, B.-J.; Dai, J.-Q.; Nazarenus, T.J.; Borges, R.; Mafra-Neto, A.; Cahoon, E.B.; Hofvander, P.; Stymne, S.; Löfstedt, C. Insect Pest Management with Sex Pheromone Precursors from Engineered Oilseed Plants. Nat. Sustain. 2022, 5, 981–990. [Google Scholar] [CrossRef]
- Xia, Y.-H.; Ding, B.-J.; Dong, S.-L.; Wang, H.-L.; Hofvander, P.; Löfstedt, C. Release of Moth Pheromone Compounds from Nicotiana benthamiana upon Transient Expression of Heterologous Biosynthetic Genes. BMC Biol. 2022, 20, 80. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.-H.; Ding, B.-J.; Wang, H.-L.; Hofvander, P.; Jarl-Sunesson, C.; Löfstedt, C. Production of Moth Sex Pheromone Precursors in Nicotiana Spp.: A Worthwhile New Approach to Pest Control. J. Pest Sci. 2020, 93, 1333–1346. [Google Scholar] [CrossRef]
- Holkenbrink, C.; Ding, B.-J.; Wang, H.-L.; Dam, M.I.; Petkevicius, K.; Kildegaard, K.R.; Wenning, L.; Sinkwitz, C.; Lorántfy, B.; Koutsoumpeli, E.; et al. Production of moth sex pheromones for pest control by yeast fermentation. Metab. Eng. 2020, 62, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Paul, R.; Ba Tis, T.; Saville, A.C.; Hansel, J.C.; Yu, T.; Ristaino, J.B.; Wei, Q. Non-Invasive Plant Disease Diagnostics Enabled by Smartphone-Based Fingerprinting of Leaf Volatiles. Nat. Plants 2019, 5, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Cao, L.; Acosta, N.; Zhu, H.; Ling, P.P. Development of Portable E-Nose System for Fast Diagnosis of Whitefly Infestation in Tomato Plant in Greenhouse. Chemosensors 2021, 9, 297. [Google Scholar] [CrossRef]
- Cui, S.; Inocente, E.A.A.; Acosta, N.; Keener, H.M.; Zhu, H.; Ling, P.P. Development of Fast E-Nose System for Early-Stage Diagnosis of Aphid-Stressed Tomato Plants. Sensors 2019, 19, 3480. [Google Scholar] [CrossRef] [Green Version]
- Kendra, P.E.; Roda, A.L.; Montgomery, W.S.; Schnell, E.Q.; Niogret, J.; Epsky, N.D.; Heath, R.R. Gas Chromatography for Detection of Citrus Infestation by Fruit Fly Larvae (Diptera: Tephritidae). Postharvest Biol. Technol. 2011, 59, 143–149. [Google Scholar] [CrossRef]
- Copolovici, L.; Popitanu, A.C.; Copolovici, D.-M. Volatile Organic Compound Emission and Residual Substances from Plants in Light of the Globally Increasing CO2 Level. Curr. Opin. Environ. Sci. Health 2021, 19, 100216. [Google Scholar] [CrossRef]
- Chidawanyika, F.; Mudavanhu, P.; Nyamukondiwa, C. Global Climate Change as a Driver of Bottom-Up and Top-Down Factors in Agricultural Landscapes and the Fate of Host-Parasitoid Interactions. Front. Ecol. Evol. 2019, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.-A.; Kansman, J.; Chuang, W.-P.; Robert, C.; Erb, M.; Felton, G.W. Water Availability and Plant–Herbivore Interactions. J. Exp. Bot. 2023, 74, 2811–2828. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M.; Ganji, S.; Gross, J.; Giesen, N.; Rid, M.; Lo, P.L.; Kokeny, A.; Unelius, C.R. Climate Change Risk to Pheromone Application in Pest Management. Sci. Nat. 2021, 108, 47. [Google Scholar] [CrossRef] [PubMed]
- Boullis, A.; Detrain, C.; Francis, F.; Verheggen, F.J. Will Climate Change Affect Insect Pheromonal Communication? Curr. Opin. Insect Sci. 2016, 17, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Stratton, C.A.; Hansen, P.; Thompson, Y.; Morrison, W.R.; Murrell, E.G. UafR: An Open-Source R Package That Automates Mass Spectrometry Data Processing. BMC J. Cheminform. 2023, in press. [Google Scholar]
- Power, A.G. Ecosystem Services and Agriculture: Tradeoffs and Synergies. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2959–2971. [Google Scholar] [CrossRef]
- Knight, I.A.; Cronin, J.T.; Gill, M.; Nyman, J.A.; Wilson, B.E.; Diaz, R. Investigating Plant Phenotype, Salinity, and Infestation by the Roseau Cane Scale as Factors in the Die-Back of Phragmites australis in the Mississippi River Delta, USA. Wetlands 2020, 40, 1327–1337. [Google Scholar] [CrossRef]
- Wheeler, G.S.; Schaffner, U. Improved Understanding of Weed Biological Control Safety and Impact with Chemical Ecology: A Review. Invasive Plant Sci. Manag. 2013, 6, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Gaffke, A.M.; Alborn, H.T.; Dudley, T.L.; Bean, D.W. Using Chemical Ecology to Enhance Weed Biological Control. Insects 2021, 12, 695. [Google Scholar] [CrossRef]
- Park, I.; Eigenbrode, S.D.; Cook, S.P.; Harmon, B.L.; Hinz, H.L.; Schaffner, U.; Schwarzländer, M. Examining Olfactory and Visual Cues Governing Host-Specificity of a Weed Biological Control Candidate Species to Refine Pre-Release Risk Assessment. BioControl 2018, 63, 377–389. [Google Scholar] [CrossRef]
- Park, I.; Schwarzländer, M.; Hinz, H.L.; Schaffner, U.; Eigenbrode, S.D. A Simple Approach to Evaluate Behavioral Responses of Insect Herbivores to Olfactory and Visual Cues Simultaneously: The Double Stacked y-Tube Device and Portable Volatile Collection System. Arthropod-Plant Interact. 2019, 13, 139–149. [Google Scholar] [CrossRef]
- Suckling, D.M.; Gibb, A.R.; Gourlay, H.; Conant, P.; Hirayama, C.; Leen, R.; Szöcs, G. Sex Attractant for the Gorse Biocontrol Agent Agonopterix ulicetella (Oecophoridae). N. Zeal. Plant Prot. 2000, 53, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Suckling, D.M.; Gibb, A.R.; Johnson, T.; Hall, D.R. Examination of Sex Attractants for Monitoring Weed Biological Control Agents in Hawaii. Biocontrol Sci. Technol. 2006, 16, 919–927. [Google Scholar] [CrossRef]
- Bartelt, R.J.; Cossé, A.A.; Zilkowski, B.W.; Weisleder, D.; Momany, F.A. Male-Specific Sesquiterpenes from Phyllotreta and Aphthona Flea Beetles. J. Chem. Ecol. 2001, 27, 2397–2423. [Google Scholar] [CrossRef]
- Gaffke, A.M.; Sing, S.E.; Dudley, T.L.; Bean, D.W.; Russak, J.A.; Mafra-Neto, A.; Peterson, R.K.D.; Weaver, D.K. Establishing Diorhabda carinulata: Impact of Release Disturbances on Pheromone Emission and Influence of Pheromone Lures on Establishment. J. Chem. Ecol. 2020, 46, 378–386. [Google Scholar] [CrossRef]
- Gaffke, A.M.; Sing, S.E.; Dudley, T.L.; Bean, D.W.; Russak, J.A.; Mafra-Neto, A.; Grieco, P.A.; Peterson, R.K.; Weaver, D.K. Semiochemicals to Enhance Herbivory by Diorhabda carinulata Aggregations in Saltcedar (Tamarix Spp.) Infestations. Pest Manag. Sci. 2018, 74, 1494–1503. [Google Scholar] [CrossRef] [Green Version]
- Gaffke, A.M.; Sing, S.E.; Dudley, T.L.; Bean, D.W.; Russak, J.A.; Mafra-Neto, A.; Peterson, R.K.D.; Weaver, D.K. Field Demonstration of a Semiochemical Treatment That Enhances Diorhabda carinulata Biological Control of Tamarix Spp. Sci. Rep. 2019, 9, 13051. [Google Scholar] [CrossRef] [Green Version]
- Oburger, E.; Jones, D.L. Sampling Root Exudates—Mission Impossible? Rhizosphere 2018, 6, 116–133. [Google Scholar] [CrossRef]
- Hsu, K.-C.; Hsieh, J.-R.; Chen, Y.-C.; Hsu, P.-F.; Hung, C.-C.; Huang, Y.-L. A Microdialysis-Based Analytical System for Dynamic Monitoring of Arsenic Transformation under Microbial Activity. Analyst 2017, 142, 4737–4743. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, G.; Rusman, Q.; Morrison, W.R., III; Magalhães, D.M.; Dowell, J.A.; Ngumbi, E.; Osei-Owusu, J.; Kansman, J.; Gaffke, A.; Pagadala Damodaram, K.J.; et al. Deciphering Plant-Insect-Microorganism Signals for Sustainable Crop Production. Biomolecules 2023, 13, 997. https://doi.org/10.3390/biom13060997
Thomas G, Rusman Q, Morrison WR III, Magalhães DM, Dowell JA, Ngumbi E, Osei-Owusu J, Kansman J, Gaffke A, Pagadala Damodaram KJ, et al. Deciphering Plant-Insect-Microorganism Signals for Sustainable Crop Production. Biomolecules. 2023; 13(6):997. https://doi.org/10.3390/biom13060997
Chicago/Turabian StyleThomas, Gareth, Quint Rusman, William R. Morrison, III, Diego M. Magalhães, Jordan A. Dowell, Esther Ngumbi, Jonathan Osei-Owusu, Jessica Kansman, Alexander Gaffke, Kamala Jayanthi Pagadala Damodaram, and et al. 2023. "Deciphering Plant-Insect-Microorganism Signals for Sustainable Crop Production" Biomolecules 13, no. 6: 997. https://doi.org/10.3390/biom13060997